
Abstract
The newly discovered strain CM-3, a Gram-negative, rod-shaped bacterium from gold mine tailings of the Central Mine in Nopiming Provincial Park, Canada, is capable of dissimilatory anaerobic reduction of tellurite, tellurate, and selenite. CM-3 possesses very high level resistance to these oxides, both aerobically and anaerobically. During aerobic growth, tellurite and tellurate resistance was up to 1500 and 1000 µg/ml, respectively. In the presence of selenite, growth occurred at the highest concentration tested, 7000 µg/ml. Under anaerobic conditions, resistance was decreased to 800 µg/ml for the Te oxides; however, much like under aerobic conditions, growth with selenite still took place at 7000 µg/ml. In the absence of oxygen, CM-3 couples oxide reduction to an increase in biomass. Following an initial drop in viable cells, due to switching from aerobic to anaerobic conditions, there was an increase in CFU/ml greater than one order of magnitude in the presence of tellurite (6.6 × 103–8.6 × 104 CFU/ml), tellurate (4.6 × 103–1.4 × 105 CFU/ml), and selenite (2.7 × 105–5.6 × 106 CFU/ml). A control culture without metalloid oxides showed a steady decrease in CFU/ml with no recovery. ATP production was also increased in the presence of each oxide, further indicating anaerobic respiration. Partial 16S rRNA gene sequencing revealed a 99.0 % similarity of CM-3 to Pseudomonas reactans.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Extreme environments have provided insight into how microorganisms can adapt to survive and thrive under the harshest of conditions. Microorganisms have been found in many environments where they are least expected such as arid deserts and hydrothermal vents (Csotonyi et al. 2006, 2010). Bacteria have evolved unique abilities that allow them to inhabit ecological niches that would be considered to be extreme for most forms of life. Some of the localities have elevated concentrations of highly toxic oxides (Knott et al. 1995; Cooper 1971) of tellurium (Te) and selenium (Se), leading to the evolution of microbes capable of tolerating the increased presence of toxins (Yurkov and Csotonyi 2003).
Te is a metalloid element related to oxygen and sulfur in group 16 of the periodic table. It possesses stable oxidation states of VI (tellurate), IV (tellurite), 0 (elemental tellurium), and II (telluride). Tellurate is most common in the hydrosphere, while in the lithosphere it is found as tellurides of gold and silver (Cooper 1971). Overall, Te has a very low global abundance (10−2–10−8 ppm) and its distribution is not homogenous (Yurkov and Csotonyi 2003). For example, in gold mines it can be significantly concentrated (14.8 ppm) (Wray 1998). Oxides of Te are highly toxic to microorganisms at levels as low as 1 µg/ml (Yurkov et al. 1996), but through aerobic reduction, certain species can resist levels as high as 2500–4000 µg/ml (Yurkov et al. 1999; Pearion and Jablonski 1999). The majority of tellurite and tellurate resistance is found under aerobic conditions, although there are some exceptions. Rhodobacter sphaeroides can reduce up to 600 µg/ml tellurite during anaerobic photosynthesis to dispose of excess reducing equivalents (Moore and Kaplan 1994). In the case of Desulfovibrio desulfuricans, resting cells can couple the reduction of tellurite to the oxidation of formate, but no energy is conserved (Lloyd et al. 2001).
Se parallels Te in many instances. It is also a group 16 element related to sulfur and oxygen with the same oxidation states of VI (selenate), IV (selenite), 0 (elemental selenium), and II (selenide). However, the toxicity of Se oxides is lower than that of Te, resulting in microbial resistance in excess of 7000 µg/ml (Rathgerber et al. 2006). Global abundance of Se, while very low, is still much higher than that of Te, around 0.05 ppm (Krauskopf 1982) and like Te, it can be locally concentrated. Sulfide ores of iron, nickel, copper, zinc and lead result in increased Se levels in mine tailings, and natural Se abundance in bedrock can lead to increased levels in soils (Haug et al. 2007). Selenite and selenate are the most prevalent forms found, sometimes at levels so high that their toxic effect can be observed in animals inhabiting those areas in the form of selenosis (Tinggi 2005). Much like Te, aerobic microbial resistance and reduction is more commonly reported than anaerobic resistance and reduction.
Dissimilatory electron transport (anaerobic respiration) to tellurite, tellurate, and selenite is a rare ability. To date, there are only seven examples of bacteria that have been confirmed to utilize these oxides as terminal electron acceptors during anaerobic growth. The first report of dissimilatory anaerobic use of tellurate was first published for the taxonomically unclassified strain ER-Te-48 (Csotonyi et al. 2006). Both Bacillus selenitireducens and B. beveridgei are the only known bacteria able to respire anaerobically using all three oxides (Baseman et al. 2007, 2009). Sulfurospirillum barnesii can undergo respiration while growing on tellurite and tellurate (Baseman et al. 2007). Lastly, there are three isolates (Desulfospirillium indicum, strain HGMK1, and HGMK3), which can use selenite for respiration (Rauschenbach et al. 2011; Takai et al. 2002). Some other bacteria are suspected to have this capability (Klonowska et al. 2005; Etezad et al. 2009), but none have been confirmed. The use of metalloid oxides as terminal electron acceptors in respiratory pathways is consistent with our current knowledge of energetics. The redox couple of TeO3 2−/Te (0.827 V), TeO4 2−/Te (1.719 V), and SeO3 2−/Se (0.875 V) are more favorable for anaerobic respiration than the SO2−/HS− redox couple (−0.217 V) utilized by sulfate reducers (Bouroushian 2010). Further supporting this line of logic, reduction of selenite to Se [ΔG f° = −132.4 kJ (mol electrons)−1] (Blum et al. 1998) and tellurite to Te [ΔG f° = −71.3 kJ (mol electrons)−1] coupled to the oxidation of lactate is highly exergonic (Baseman et al. 2009), providing more than enough energy for growth. Likely factors contributing to the infrequent use for respiration is the low global abundance and high toxicity. Nevertheless, respiration on other toxic oxides is known (Lovley 1993), indicating toxicity does not always prevent inclusion in metabolism. Until recently, Te has been considered a biologically unimportant element. However, the discovery of respiration based on tellurite and tellurate revealed newly discovered biological relevance (Csotonyi et al. 2006). Much like Se, which was considered biologically insignificant for many years, it is now understood that Se is essential for some forms of life (Haug et al. 2007), and Te may follow a path similar to Se as future research focuses on the metalloid.
The goal of this work was to identify bacteria possessing very high levels of aerobic and anaerobic resistance to metal(loid) oxides, as well as the ability to perform anaerobic respiration, isolated from tailings of the Central Mine in Nopiming Provincial Park, Manitoba, Canada. This gold mine opened in 1927 and closed in 1937 (Sherriff et al. 2007). Since then, the site has been virtually untouched, providing ideal conditions for resident bacteria to establish a stable community. We describe a novel bacterium, strain CM-3, capable of very high level aerobic and anaerobic resistance to tellurite, tellurate, and selenite and of anaerobic respiration.
Materials and methods
Isolation and metalloid oxide resistance
Samples from the tailings of the Central Mine in Nopiming Provincial Park, Manitoba, Canada, were obtained in August of 2011. A 1 cm3 sample was resuspended in sterile water (10 ml) and used to directly inoculate Balch tubes containing Minimal Salts (MS) medium (Yurkov et al. 1996) supplemented with 1 g/l lactate and 25 µg/ml K2TeO3 for enrichment of anaerobic tellurite-resistant bacteria. Tubes were incubated at 28 °C in the dark and observed for visual blackening due to reduction of tellurite to elemental Te (Yurkov et al. 1996) as well as monitoring of tellurite concentration in the medium by the NaBH4 method (Molina et al. 2010). A sample was taken from tubes showing reduction and used to inoculate rich organic (RO) (Yurkov et al. 1996) plates containing 25 µg/ml K2TeO3, which were incubated at 28 °C in the dark in an anaerobic chamber. Resulting colonies were selected and replated on RO plates and incubated with the same conditions as previously for both aerobic and anaerobic conditions. Phase contrast microscopy was performed using a Ziess Axioskope 2 microscope on the selected strains and a representative was chosen for all further testing.
A 5 % inoculum of an overnight culture of aerobically grown cells was used to determine resistance to tellurite, tellurate, selenite, and selenate. Aerobic resistance was tested in RO liquid medium containing 100 µg/ml of one of these four oxides. Cultures were incubated at 28 °C in the dark on an incubator shaker at 200 rpm. Resistance and reduction were monitored visually in terms of blackening (tellurite and tellurate) or reddening (selenite, selenate) of the culture indicating the appearance of elemental Te and Se, respectively (Rathgerber et al. 2006). The level of resistance was examined using 250, 500, 750, 1000, 1500, 2000 µg/ml for tellurite and tellurate and 1000, 2000, 3000, 4000, 5000, 6000, and 7000 µg/ml for selenite. Balch tubes containing liquid anaerobic metalloid respiration (AMR) medium [in g/l: KH2PO4, NH4Cl, (0.5); CaCl2, (0.1); yeast extract, lactate (1); MgSO4, (trace); vitamins and trace metals solution (2 ml/l) (Drews 1983)] and 100 µg/ml of one of K2TeO3, K2TeO4, Na2SeO3 or Na2SeO4 were used to investigate anaerobic resistance. Tubes were incubated at 28 °C in the dark. Level of resistance anaerobically was monitored using (in µg/ml) 200, 400, 600, 800, and 1000 for the Te oxides and 500, 1000, 1500, 2000, 3000, 4000, 5000, 6000 and 7000 for selenite. All experiments were performed in triplicate.
Kinetics experiments
Aerobically grown cells of CM-3 were injected into 120-ml crimp-sealed bottles containing 75 ml of AMR medium amended with tellurate, tellurite, or selenite at 50 µg/ml under a headspace of N2. Samples were taken every 24 h for CFU/ml and ATP counts. CFU/ml was monitored as an assay for growth by serial dilution to 10−5 followed by plating on RO plates incubated at 28 °C. Colonies were counted after 7 days. ATP was measured using an ATP Bioluminescence Kit from Sigma–Aldrich.
Phylogenetic analysis
Isolation of genomic DNA was performed as described (Chen and Kuo 1993). Partial sequencing of the 16S rRNA gene was performed by PCR utilizing bacterial primers U1 and U1R (James 2010). Amplifications were performed in 50 µl reaction volumes containing 25 µl DreamTaq PCR Master Mix, 0.25 μM of each primer, and between 10 and 50 ng of DNA in a thermal cycler with the following conditions: initial denaturing at 95 °C for 5 min, then denaturing at 95 °C for 30 s, annealing at 46 °C for 30 s, extension at 72 °C for 1.5 min for 35 cycles with a final extension at 72 °C for 10 min ending with a 7 °C hold. Preparation and sequencing of the PCR product, nucleotide sequence editing was carried out as published (Fontaine et al. 2012). An NCBI Blast search was performed to determine the nearest relative and a Maximum Likelihood phylogenetic tree created using MEGA 6.0 (Tamura et al. 2013). Bootstrap analyses were performed with 1000 replicates.
Nucleotide sequence accession number
The 16S rRNA gene sequence determined in this study for CM-3 was deposited in GenBank under the accession number KR150623.
Results and discussion
Strain isolation and metalloid oxide resistance analysis
Extreme environments have provided us with numerous isolates capable of resistance to and reduction of metalloid oxides (Yurkov et al. 1999; Yurkov and Csotonyi 2003; Csotonyi et al. 2006; Rathgeber et al. 2006; Baseman et al. 2007, 2009). The same is true for the microbial community inhabiting the tailings of the Central Mine, which are spread out over a wide area (Fig. 1a). Sites chosen for sampling were representative of the environmental conditions found throughout the location. Site 1 (Fig. 1b), referred to as the Blue Pond, was a transient pond, with water levels dependent on precipitation. This location experienced fluctuating humidity, temperature, and pH. Oxide concentrations also varied through dilution (when water is present) and concentration (when dry). At the time of sampling, the Blue Pond was virtually dry due to lack of rain, with a pH of 4.1 at the moist bottom. Site 2 (Fig. 1c) was at the former mine drainage system and today where precipitation runs off from the mine site proper. The site was arid above and below the surface, with a pH of 3.9. Site 3 (Fig. 1d) was similar to Site 2 except while dry on the surface, there was still moisture below, resulting in a pH of 7.0. Lastly, Site 4 (Fig. 1e) was at a location called the Green Pond, which unlike the Blue Pond, was permanent and at the time of sampling contained water and had a pH of 6.9.

a Schematic map of mine tailings from the Central Mine in Nopiming Provincial Park, Manitoba, Canada, showing location of sampling; Photographs of b site 1, c site 2, d site 3, e site 4
Upon observation of Balch tubes with tellurite containing medium and inoculated with an environmental sample from each of the four sites, we noted blackening of the culture only at Site 3. To confirm formation of the elemental Te as a result of anaerobic tellurite reduction, levels of TeO3 2− in the growth medium were monitored over time (Fig. 2). Tellurite was decreasing in conjunction with visible darkening, confirming anaerobic reduction. To identify the microorganism(s) responsible, samples taken from an actively reducing enriched Balch tube were transferred onto plates. After 7 days, several small black colonies appeared and they were selected for further analysis. All isolates were facultative anaerobes growing under both aerobic and anaerobic conditions, forming small, circular colonies 1–2 mm in diameter and microscopically cells were similar (Fig. 3). One strain, CM-3, was further investigated for its resistance to Te and Se oxides in both aerobic and anaerobic conditions. This rod-shaped bacterium is 1.8–2.6 µm in length, Gram negative, and motile. During aerobic growth, CM-3 possesses very high level resistance to tellurite (1500 µg/ml), tellurate (1000 µg/ml), and selenite (>7000 µg/ml). No resistance to or reduction of selenate was observed. The range of oxide resistance was comparable under anaerobic conditions (Fig. 4), with level of resistance decreased, but still remaining very high (800 µg/ml for tellurite and tellurate and 7000 µg/ml for selenite). Hence, strain CM-3 can reduce and resist extreme levels of tellurite, tellurate, and selenite under both aerobic and anaerobic conditions.

Change in concentration of tellurite (µg/mL) in the growth medium over time (days) by CM-3 samples taken from Site 3

Phase contrast micrograph of strain CM-3 isolated from site 3 showing rod-shaped bacteria

Results of the test for anaerobic resistance and reduction of 100 µg/ml tellurite, tellurate and selenite by CM-3. The color change to black is due to reduction of tellurite/tellurate to elemental Te and reddening is due to reduction of selenite to elemental Se. − no oxide present, + oxide present
Kinetics of growth and ATP production
Reduction of an oxide under anaerobic conditions does not automatically confirm respiration. For evidence, we monitored growth (CFU/ml) in AMR medium containing one of tellurite, tellurate, or selenite as the only possible terminal electron acceptor. Growth and reduction in such conditions would indicate anaerobic respiration. An increase in CFU/ml of greater than an order of magnitude was detected for all three oxides. Initially there was a significant drop in CFU/ml for the Te oxides, which can be attributed to CM-3 adapting to anaerobic growth as well as to the initial toxic shock from newly introduced tellurite or tellurate. However, shortly after, CFU/ml increased from 6.6 × 103 to 8.6 × 104 in the presence of tellurite over 5 days (Fig. 5a) and 4.6 × 103–1.4 × 105 with tellurate over 7 days (Fig. 5b). In the case of the anaerobic selenite experiment, the initial drop in CFU/ml was not as significant as it might be less toxic to cells than the Te oxides and recovery was faster. The CFU/ml increased from 2.7 × 105 to 5.6 × 106 over 3 days (Fig. 5c). During the same time, the number of cells in control tubes without an oxide immediately began to decline without recovery.

Changes in growth of CM-3 (CFU/mL) over time (days) with added oxides as the sole terminal electron acceptor. a K2TeO3, b K2TeO4, c Na2SeO3. Error bars represent one standard deviation
The previous data showed anaerobic growth, with oxides as terminal electron acceptors. Next, ATP levels in the presence and absence of metalloid oxides were monitored to further confirm anaerobic respiration. As expected, ATP production per CFU/ml was much higher during log phase (Fig. 6a–c). Growth coupled to increased ATP production, along with cell death in the absence of an oxide, strongly confirms that CM-3 is performing dissimilatory reduction of tellurite, tellurate, and selenite under anaerobic conditions.

Change in ATP production (ATP/CFU/ml) over time (days) by cells of CM-3 during anaerobic growth in the presence of metalloid oxides. a K2TeO3, b K2TeO4, c Na2SeO3. Error bars represent one standard deviation
Phylogenetic analysis
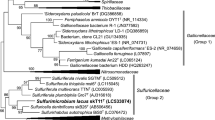
Partial sequencing of the 16S rRNA gene (726 bp) revealed CM-3 is a member of the genus Pseudomonas and most closely related to Pseudomonas reactans (99.0 %) (Fig. 7). Some members of this genus are capable of aerobic Te and Se oxide resistance and reduction (Lortie et al. 1992; Malik and Jaiswal 2000; Rajwade and Paknikar 2003; Hunter and Manter 2009); however, the reported resistance [e.g., tellurite MIC of 150 µg/ml (Zanaroli et al. 2002)] is extremely low when compared to CM-3. Pseudomonas stutzeri pn1 is able to anaerobically respire on selenate (Narasingarao and Haggblom 2007), while P. isachenkovii and P. vanadiumreductans can use metavanadate (Yurkova and Lyalikova 1990). However, there are no reports of either selenite or Te oxide respiration, and strain CM-3 makes the first example.

Maximum likelihood phylogenetic tree of strain CM-3 showing its position among species of the genus Pseudomonas. (Bar represents 0.02 %)
Summary
This work identified CM-3, a novel bacterium related to Pseudomonas reactans (99.0 %), which possesses very high levels of aerobic and anaerobic resistance to tellurite, tellurate, and selenite, as well as the ability to use oxides for dissimilatory anaerobic respiration. CM-3 is only the fourth reported case of a tellurite respiring microbe and fifth for tellurate and selenite. Also, other than B. selenitireducens and B. beveridgei, this is the only other isolate ever found capable of utilizing all three oxides and is the only Gram-negative representative. Our discovery provides evidence that Pseudomonas species possess the rare ability to anaerobically respire on tellurite, tellurate, and selenite as well as a wider range and greater level of metalloid resistance than previously thought.
With the search for more environmentally friendly methods for cleanup of locations contaminated by oxides, microbes, such as CM-3, show great potential. The use of bacteria for bioremediation might provide a more ecologically responsible method for removal of contaminants. As well, the ability to transform oxides to pure elemental forms may be useful in bacterial recycling and recovery of metal(loid)s, such as Te, for industrial purposes with the least amount of negative impact on the environment.
References
Baseman S, Bullen T, Dewald J, Zhang D, Curran S, Islam F, Beveridge T, Oremland R (2007) Formation of tellurium nanocrystals during anaerobic growth of bacteria that use Te oxyanions as respiratory electron acceptors. Appl Environ Microbiol 73(7):2135–2143
Baseman S, Stolz J, Kulp T (2009) Enrichment and isolation of Bacillus beveridgei sp. Nov., a facultative anaerobic haloalkaliphile from Mono Lake, California, that respires oxyanions of tellurium, selenium, and arsenic. Extremeophiles 13:695–705
Blum J, Bindi A, Buzzelli J, Stolz J, Oremland R (1998) Bacillus arsenicoselenatis, sp. nov., and Bacillus selenitireducens, sp. nov.: two haloalkaliphiles form Mono Lake, California that respire oxyanions of selenium and arsenic. Arch Microbiol 171:19–30
Bouroushian M (2010) Electrochemistry of metal chalcogenides. Springer-Verlag, Berlin
Chen W, Kuo T (1993) A simple and rapid method for the preparation of gram-negative bacterial genomic DNA. Nucleic Acids Res 21(9):2260
Cooper W (1971) Tellurium. Van Nostrand Reinhold Company, New York
Csotonyi J, Stachebrandt E, Yurkov V (2006) Anaerobic respiration on tellurate and other metalloids in bacteria from hydrothermal vent fields in the eastern Pacific Ocean. Appl Environ Microbiol 72(7):4950–4956
Csotonyi J, Swiderski J, Stackebrandt E, Yurkov V (2010) A new extreme environment for aerobic anoxygenic phototrophs: biological soil crusts. Adv Exp Med Biol 675:3–14
Drews G (1983) Mikrobiologisches praktikum. Springer-Verlag, Berlin
Etezad S, Khajeh K, Soudi M, Ghazvini P, Dabirmanesh B (2009) Evidence on the presence of two distinct enzymes responsible for the reduction of selenate and tellurite in Bacillus sp. STG-83. Enzyme Microb Technol 45:1–6
Fontaine K, Beck A, Stocker-Worgotter E, Piercey-Normore M (2012) Photobion relationships and phylogenetic history of Dermatocarpon luridum var. luridum and related Dermatocarpon species. Plants 1:39–60
Haug A, Graham R, Christophersen O, Lyons G (2007) How to use the world’s scarce selenium resources efficiently to increase the selenium concentration in food. Microb Ecol Health Disease 19:209–228
Hunter W, Manter D (2009) Reduction of selenite to elemental red selenium by Pseudomonas sp. strain CA5. Curr Microbiol 58:493–498
James G (2010) Universal bacterial identification by PCR and DNA sequencing of 16s rRNA gene. In: Schuller M et al (eds) PCR for clinical microbiology. Springer Science + Business Media B.V, Netherlands, pp 209–214
Klonowska A, Heulin T, Vermeglio A (2005) Selenite and tellurite reduction by Shewanella oneidensis. Appl Environ Microbiol 71(9):5607–5609
Knott R, Fallick A, Rickard D, Backer H (1995) Mineralogy and sulfur isotope characteristics of a massive sulfide boulder, Galapagos Rift, 85°55′W. In: Parson L, Dixon D, Walker C (eds) Hydrothermal vents and processes. Geological Society, London, pp 207–222
Krauskopf K (1982) Introduction to geochemistry, 2nd edn. McGraw-Hill Book Company, Singapore
Lloyd J, Mabbett A, Williams D, Macaskie L (2001) Metal reduction by sulphate-reducing bacteria: physiological diversity and metal specificity. Hydrometallurgy 59:327–337
Lortie L, Gould W, Rajan S, McCready R, Cheng K (1992) Reduction of selenate and selenite to elemental selenium by a Pseudomonas stutzeri isolate. Appl Environ Microbiol 58(12):4042–4044
Lovley D (1993) Dissimilatory metal reduction. Annu Rev Microbiol 47:263–290
Malik A, Jaiswal R (2000) Metal resistance in Pseudomonas strains isolated from soil treated with industrial wastewater. World J Microbiol Biotechnol 16:177–182
Molina R, Burra R, Perez-Donoso J, Elias A, Munoz C, Montes R, Chasteen T, Vasquez C (2010) Simple, fast, and sensitive method for quantification of tellurite in culture media. Appl Environ Microbiol 76(14):4901–4904
Moore M, Kaplan S (1994) Members of the family Rhodospirillaceae reduce heavy-metal oxyanions to maintain redox poise during photosynthetic growth. ASM News 60:17–23
Narasingarao P, Haggblom M (2007) Identification of anaerobic selenate-respiring bacteria from aquatic sediments. Appl Environ Microbiol 73(11):3519–3527
Pearion C, Jablonski P (1999) High level, intrinsic resistance of Natronococcus occultus to potassium tellurite. FEMS Microbiol Lett 174:19–23
Rajwade J, Paknikar K (2003) Bioreduction of tellurite to elemental tellurium by Pseudomonas mednocina MCM B-180 and its practical application. Hydrometallurgy 71:243–248
Rathgeber C, Yurkova N, Stackebrandt E, Schumann P, Humphrey E, Beatty T, Yurkov V (2006) Metalloid reducing bacteria isolated from deep ocean hydrothermal vents of the Juan de Fuca ridge, Pseudoalteromonas telluritireducens sp. nov. and Pseudoalteromonas spiralis sp. nov. Curr Microbiol 53:449–456
Rauschenbach I, Narasingarao P, Haggblom M (2011) Desulfurispirillum indicum sp. nov., a selenate- and selenite-respiring bacterium isolated from an estuarine canal. Int J Syst Evol Microbiol 61:645–658
Sherriff B, Sidenko N, Salzsauler K (2007) Differential settling and geochemical evolution of tailings’ surface water at the Central Manitoba Gold Mine. Appl Geochem 22:342–356
Takai K, Hirayama H, Sakihama Y, Inagaki F, Yamato Y, Horikoshi K (2002) Isolation and metabolic characteristics of previously uncultured members of the order Aquificales in a subsurface gold mine. Appl Environ Microbiol 68(6):3046–3054
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30(12):2725–2729
Tinggi U (2005) Selenium toxicity and its adverse health effects. Rev Food Nutr Toxic 4:29–55
Wray D (1998) The impact of unconfined mine tailings and anthropogenic pollution on a semi-arid environment—an initial study of the Rodalquilar mine district, southeast Spain. Environ Geochem Health 20:29–38
Yurkov V, Csotonyi J (2003) Aerobic anoxygenic phototrophs and heavy metalloid reducers from extreme environments. Recent Res Dev Bacteriol 1:247–300
Yurkov V, Jappe J, Vermeglio A (1996) Tellurite resistance and reduction by obligately aerobic photosynthetic bacteria. Appl Environ Microbiol 62:4195–4198
Yurkov V, Krieger S, Stackebrandt E, Beatty T (1999) Citromicrobium bathyomarinum, a novel aerobic bacterium isolated form deep-sea hydrothermal vent plume waters that contains photosynthetic pigment-protein complexes. J Bacteriol 181(15):4517–4525
Yurkova N, Lyalikova N (1990) New vanadate-reducing facultative chemolithotrophic bacteria. Mikrobiologiya 59(6):968–975
Zanaroli G, Fedi S, Carnevali M, Fava F, Zannoni D (2002) Use of potassium tellurite for testing the survival and viability of Pseudomonas pseudoalcaligens KF707 in soil microcosms contaminated with polychlorinated biphenyls. Res Microbiol 153:353–360
Acknowledgments
This work was supported by the NSERC Discovery and University of Manitoba GETS grants held by V. Yurkov and an NSERC Discovery grant held by M. Piercey-Normore. We would also like to thank Breanne Head and Elizabeth Hughes for their assistance with sampling.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Albers.
Rights and permissions
About this article

Cite this article
Maltman, C., Piercey-Normore, M.D. & Yurkov, V. Tellurite-, tellurate-, and selenite-based anaerobic respiration by strain CM-3 isolated from gold mine tailings. Extremophiles 19, 1013–1019 (2015). https://doi.org/10.1007/s00792-015-0776-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00792-015-0776-8