
Abstract
Intense droughts in recent years are a global concern. The duration and timing of drought for forests have not been researched as much as crops. This study aims to utilize big earth data to analyse the consequences of drought on forest based on drought and vegetation indices derived from remote sensing. The study area is Sal Forest (Shorea robusta) in Bangladesh. Vegetation indices such as NDVI and EVI are correlated with drought indices for the years 2000–2020. The data is collected via Google Earth Engine and the R statistical software environment is used for data analysis. Seasonal adjustment was conducted to remove the biases due to seasonality. Results showed that the timing for the loss of forest vitality is dependent on drought severity. Lagged correlation was used to identify the lags between drought severity and vitality via drought and vegetation indices. From 2000 to 2020, the intensity of drought was moderate (0.95). There is a lag between drought and a decrease in forest vitality. This lag decreases as the severity and duration of drought increase. Severe droughts in Sal Forest create an almost immediate impact with worsening conditions but show a lag of 3–4 months and 2–3 months for low and moderate drought severity, respectively. The NDVI anomaly shows that drought has adverse effects during growth periods. The methodology of this study can be replicated across various forest types, and further improvements would require in situ data coupled with phenological observations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Drought is different from most other natural hazards because of not having a set time frame. It is often termed a “creeping phenomenon” (Wilhite et al. 2014). The incidence and severity of droughts have increased globally (Spinoni et al. 2014; Vogel et al. 2020). However, defining drought is complex due to variability in duration, intensity, timing, species and vegetation sensitivity and spatial coverage (Camarero et al. 2015; Anderegg et al. 2015; Peltier et al. 2016; Huang et al. 2018; Kannenberg et al. 2019; Bose et al. 2020, 2021; Serra-Maluquer et al. 2021). Tree mortality and forest dieback as direct impacts of drought are growing concerns (Spinoni et al. 2014). Despite extensive research, uncertainties remain about the impact of droughts on trees (Anderegg et al. 2015; Gazol et al. 2017, 2020; Bose et al. 2021; Ovenden et al. 2021). In-depth knowledge of a particular tree or species is necessary because of the variability of impacts and recovery among trees. The objective of this paper is to quantify the lag between drought severity and forest vitality in Shorea robusta (Sal Forest) from the onset of drought in Bangladesh.
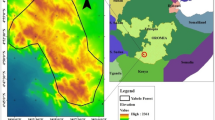
Bangladesh ranks among the top five disaster-prone countries in the Asia–Pacific region (Ahmad 2021). However, drought has gained little attention compared to other hazards with immediate devastating impacts i.e. floods and tropical cyclones. Drought is a recurrent event at an average 2.5-year interval in Bangladesh (Adnan 1993; Sultana et al. 2021). The frequency of drought is expected to rise due to climate change (National Drought Mitigation Center 2021), and Bangladesh is expected to have lower rainfall during the dry season and more intense rainfall during monsoon (Christensen et al. 2007; Shahid and Behrawan 2008). The north-western region of Bangladesh is the most drought prone due to rain variability (Shahid and Behrawan 2008; Habiba and Shaw 2013). Based on the Palmer Drought Severity Index (PDSI), the regional drought variability of Bangladesh between 2000 and 2020 is shown in Fig. 1.

Spatiotemporal characteristic of drought (PDSI) in Bangladesh in 2000–2020
Application of remote sensing for drought impact assessment on forests is gaining popularity due to an acceptable level of accuracy and replicability (Deshayes et al. 2006). However, accuracy of remote sensing is subject to image resolution, the selection of tree species and on researchers’ expertise. The use of MODIS (moderate resolution imaging spectroradiometer) is popular among researchers due to its revisit time (see for example, Haroon et al. 2016; Byer and Jin 2017; Kim et al. 2017; Xulu et al. 2018; Cammalleri et al. 2019). High-resolution images are rarely used for drought impact analysis because of the limited temporal resolution and accessibility (Garrity et al. 2013; Elhag and Zhang 2018). Additionally, there is complexity over the selection of appropriate tree or forest-specific indices. The Handbook of Drought Indicators and Indices, 2017 outlines 10 remotely sensed indices suitable for analysing drought impact.Footnote 1 All the indices, except the Soil Adjusted Vegetation Index (SAVI), are easy to use, readily and freely available and require minimum data (Svoboda and Fuchs 2017).
This study aims to utilize big earth data to analyse the consequences of drought on forest based on drought and vegetation indices derived from remote sensing. The true strength of such a study is the vast amount of data that can be used to interpret results between the drought indices and vegetation indices to assess the condition of forests after drought. This study uses correlations with a delay in time (lag) to find how the intensity and duration of drought translate to this lag with respect to vegetation indices. To the best of our knowledge, there has been no such study in Bangladesh that uses remote sensing to assess drought lag time impact on Sal Forest. This is particularly useful for monitoring forest status continuously, thus helping policymakers strategize forest conservation based on past drought experiences.
2 Materials and methods
2.1 Study area
Shorea robusta is a semi-evergreen species despite it being widely distributed in tropical moist and dry forests. It can survive in different habitats depending on climatic conditions (Singh and Kushwaha 2005). The largest belt of Shorea robusta lies in the central part of the Ganges–Brahmaputra-Meghna Delta. The Forest Department of Bangladesh manages four major types of forestlands covering 10.54% (4.65% hill forests, 4.09% natural mangroves, 0.97% mangrove plantations and 0.83% Sal Forests) of the country’s total area (Arfin Khan et al. 2007). The distribution of Shorea robusta in Bangladesh is patchy but mostly found in the drier central and northern parts of the country.
The research focused on Sal Forest located in the drier central part of Bangladesh, including Gazipur, Tangail, Mymensingh and Jamalpur districts (Fig. 2 (a)). Anthropogenic factors are attributed to the continuous depletion of Sal Forest. As per the Food and Agricultural Organization (FAO), an estimated 10% of the total Shorea robusta remained in 1990 (Rahman et al. 2010). The central region of Bangladesh, which is the study area of this study, has the most threatened forest cover due to population and development pressures (Alam et al. 2008). The study area is part of a reserve park which is free from rampant deforestation and logging. This ensures that the impact of drought and its consequences are the major causes of loss of forest vitality.

a Study area location and b elevation
The area is mostly covered by Pleistocene terraces and recent alluvial floodplains (Dhar and Mridha 2006). The forest soil in this area is usually sandy loam to silty loam in texture. Figure 2b illustrates the flat topographic conditions with no major variation in elevation in Sal Forest. Sal tree are the dominant plant species in the study area. It is one of the most significant wood-producing plants in Bangladesh due to its enduring capacity.
Bangladesh is a tropical country. The country has four distinct seasons of rainfall: (i) dry winter (December-February); (ii) pre-monsoon hot summer (March–May); (iii) monsoon (June–September); and (iv) post-monsoon (October–November). About 60–75% of the annual rainfall occurs during the monsoon (Islam and Uyeda 2005; Shahid 2010). The annual average winter temperature ranges from 18 °C in the north to 21 °C in the south, and the summer temperature ranges from 27 °C in the coastal (east) to 30 °C in the west (Shahid 2011; Sarker 2021). The annual rainfall varies from 1499 mm in the northwest to over 4000 mm in the northeast (Shahid 2010). The climatic conditions are very consistent over Sal Forest. It has an annual average rainfall of 2000–2300 mm and a 26.3 °C of temperature (Rahman et al. 2010).
2.2 Methods
2.2.1 Data acquisition
Data for the various indices were acquired using Google Earth Engine. Ensuring data consistency is essential when comparing various types of land uses. To maintain data consistency, the “Hansen Global Forest Change v1.8 (2000–2020)” data was used to determine forest areas. It is especially useful in determining annual forest extent and canopy cover change. The data is in raster format and provides forested pixels based on tree canopy cover for vegetation taller than 5 m. It was necessary to obtain time series data at pixels that were and remained as forest land cover. This was essential to negate the effects of deforestation and logging.
For data acquisition, the JavaScript API of Google Earth Engine (Gorelick et al. 2017) was used. The R environment v3.6.3 (R Core Team, 2021) was used for data processing. QGIS (QGIS Development Team, 2021) was used for data visualization. The data and various packages used in R data processing and analysis are outlined in Table 1.
2.2.2 Vegetation and drought indices
Vegetation and drought indices were extracted from MODIS and TerraClimate, respectively. A total of 3 indices, namely, Normalized Difference Vegetation Index (NDVI), Enhanced Vegetation Index (EVI) and Palmer Drought Severity Index (PDSI), along with climatic water balance data (precipitation and evapotranspiration), were derived.
The MODIS Terra Vegetation Indices 16-Day Global 250 m data product (MOD13Q1.006) was used. Vegetation layers (NDVI and EVI) were obtained using the JavaScript API of Google Earth Engine. On the same platform, quality assurance (QA) (Vegetation Index quality and QA bitmask) was also performed to remove inconsistencies caused by cloud and shadow in satellite images. The inconsistencies were removed using the VI (Vegetation Index) quality indicators present in the dataset. There are specific data masking options for specific issues such as quality, usefulness, aerosol quantity, adjacent cloud detected, atmospheric correction, mixed clouds, land/water mask, possible snow/ice and possible shadow. Images dated from 2000 to 2020 were used. The MOD13Q1.006 dataset has a spatial resolution of 250 m and a temporal resolution of 16 days. The 16-day intervals were aggregated monthly; therefore, if there are 2 images for a particular month, then the median values are taken for all the pixels in the study area. This was done for all the months from the year 2001 to 2020. Since the data acquisition started from 2000–02-18, median values of 11 months were considered for the year 2000. Time series data was then extracted from the forested areas. This was also used for anomaly calculation in NDVI values for the study area.
Drought indices depend on various inputs such as precipitation, evapotranspiration and temperature along with an appropriate energy balance method. Precipitation, evapotranspiration and temperature data were acquired from the TerraClimate dataset via Google Earth Engine from 2000 to 2020. A total of 251 images were used for the vegetation and drought indices. They were partitioned monthly for the entire period from 2000 to 2020. Monthly medians were used for both the indices. The dataset has a spatial resolution of 4638.3 m and a temporal resolution of 1 month. The Palmer Drought Severity Index (PDSI) is a standardized index that is derived from temperature and precipitation data (Alley 1984). PDSI is generally used for long-term droughts and is susceptible to drought scenarios that occur on smaller timescales. One of the major drawbacks of PDSI is thus avoided as the study area does not exhibit delayed runoff from snow or ice. Calculation of drought at smaller timescales is necessary for a thorough examination of drought resistance within a forest ecosystem. Therefore, the Standardized Precipitation Evapotranspiration Index (SPEI) was also taken into consideration (Vicente-Serrano et al. 2010; Ogaya et al. 2015; Potopová et al. 2015). SPEI is derived from precipitation and potential evapotranspiration (PET). It has a significant advantage over other frequently used drought indices that address the effect of PET on drought severity. This is because its multi-scalar properties allow for the identification of various drought types. Precipitation data was normalized using a Box-Cox transformation, and data processing was done through the “SPEI” R package. SPEI has timescales ranging from 1 to 48 months. To identify lags, a 1-month and 3-month timescale was used, as further aggregation could produce misleading results.
2.2.3 Data processing
The time series data for each of the indices was mined for forest pixels that lay within the study area. Data pre-processing at this stage included the removal of null values and smoothing the time series to prepare them for data analysis. Random forest pixels were also taken to identify potential outliers in the dataset. Drought indices were normalized along with vegetation indices and fed as time series data in R.
Each dataset’s time series data was decomposed into its primitive components (trend, seasonal structure and randomness associated within the dataset) using two R package (i.e. “astsa” v1.13 and “tseries” v0.10–48) (Stoffer 2014; Trapletti et al. 2015). The datasets subsequently underwent seasonal adjustment (Panuju et al. 2021). This was done by subtracting the raw time series from the seasonal time series component of the data.
The stationarity of the time series was examined and inspected to determine if the individual datasets were additive or multiplicative in nature. Although this was necessary for vegetation indices, drought indices were not investigated because they did not have any seasonal pattern (Fig. 1). Stationarity for the time series of the datasets was verified using the Augmented Dickey-Fuller and Kwiatkowski-Phillips-Schmidt-Shin tests (Nielsen 2006). Seasonal adjustment added value to the dataset and showed how forests behave normally, aside from following a cycle of growth to maturity (Kebłowski and Welfe, 2004). This was further cemented via insignificant results with autocorrelation and partial autocorrelation. Combining the median values for the NDVI and PDSI datasets does not reveal any specific trends that explain the relationship between them (Fig. 3).

Median data values of NDVI and PDSI (normalized) from 2000 to 2020
2.2.4 Drought intensity and duration
The number of months between the onset and offset of a drought event is the monthly duration of the drought (Gustafson and Sturtevant 2013; Cammalleri and Vogt 2019). During the entire duration (m), the absolute sum of SPEI is the severity (Se). Intensity (DIe) is therefore calculated by dividing severity by duration (Tan et al. 2015). Runs theory is popular for this process (Yevjevich 1969; Pei et al. 2020; Sáenz et al. 2021). Since the absolute value is taken into consideration, greater intensity equals severe drought consequences. The equations for drought severity and intensity are:
where e = drought event; j = month; Indexj = SPEI value in month j.
Temporal characteristics were further explored by calculating the trend of a drought event in a particular year. The trend lines have been calculated using a second-degree polynomial curve equation. The area under these curves was calculated using the following equation.
where a and b are the limits of the curve in x-axis.
A threshold value of − 3.00 was selected for the drought year detection. This value is subjected to variation depending on the region and climatic factors that govern that region.
2.2.5 Lagged correlation
To calculate the timing of drought events in the forest, time series data were correlated with time lags to obtain correlation coefficients. The lagged correlation coefficients are pitted against drought severity and duration to assess the impact of drought on forests and are carried out on a yearly basis. The lagged correlation is taken in months to assess the immediate impact of drought on forests. This study does not deal with projected impact felt by forests and thus, long-term lags to drought have been excluded from the scope.
Direct relations between vegetation and drought indices did not yield any significant relationship. The onset of the drought itself was not met with the immediate deterioration of forest and there was a delay between them. Instead, a lag correlation was identified between the vegetation indices and the drought indices (Eq. 4).
where rk(x,y) refers to the series of correlation coefficients between two indices under time lag k, xi and yi are the series of drought and vegetation indices (NDVI and EVI against PDSI and SPEI) and n is the length of the series. The overall methodological approach is graphically presented in Fig. 4.

Methodological approach of the study
3 Results
3.1 Drought intensity and duration
Forest roots and their interaction with root zone soil moisture provide a better understanding of the delayed effects of drought (Brunner et al. 2015; Leuschner and Meier 2018). A deeper root system is an important characteristic of Shorea robusta (Singh and Kushwaha 2005). This enables Sal trees to sustain themselves during drought onset. Correlation coefficients from the lagged time series analysis show different behaviours for different years. This is attributed to the intensity and duration of drought coupled with tree characteristics such as growth period and phenology. There is an inverse relationship between the lag and intensity of drought (Tenhumberg et al. 2018). As drought intensifies, the lag time observed for the impacts of drought is reduced. The drought year detection taking a threshold of − 3.00 using the second-degree polynomial curve show that the years 2006, 2012–2015, 2018 and 2019 experienced greater drought stress compared to the other years. The area covered by drought has been shown explicitly in Figs. 5 and 6. This is specifically observed for the years 2006 and 2018–2019. Analysis of SPEI (1 month) shows how the droughts occurred from 2000 to 2020 (Fig. 5) with an intensity of 0.95 (moderate).

SPEI (1 month) over the study area from 2000 to 2020

SPEI (1 month) values aggregated over the study area with a polynomial trend line
We can estimate the consequence of drought on the study area by analysing the drought intensity and duration as a second-degree polynomial trend line (Fig. 6). Aside from the drought of 2016, the trend line of droughts is similar with varying intensities. This shows how the drought has progressed over the entire year. A substantial impact is felt over the growing season for the forest. A deeper curvature of the trend line shows its intensity for the study area. The area under the curves (raw data and not trend line) for the drought years was calculated to provide further insights. The drought of 2013 was by far the most severe. It should also be noted that drought has intensified over the latter half of the decadal study. The first decade (2000–2010) did not experience much drought apart from the year 2006. However, multiple drought years were observed for the second decade (2011–2020) of the study. There are extensions of drought that cover more than a single year, which can be seen from Fig. 1 for the years 2014–2015. It is also important to note that in the latter part of the time series data (2011–2020), drought seems to have intensified across the study area. This may have been an aftermath of global warming and climate extremes that are being experienced all over the world (Hartmann et al. 2015).
3.2 NDVI and EVI anomaly
The mean of the median values for each month from 2000 to 2020 was computed and the same was done for the individual drought years. The mean of medians has been used since it is more susceptible to outliers than mean values. A shift from the normal values was thus calculated and plotted in Fig. 7 and Fig. 8. The anomaly was thus based on data from over two decades. This was necessary given the shift in drought over the second decade. Shorea robusta is deciduous in nature and consists of dormant and growth seasons. After the end of the rainy season, this dormant period is cast out by the growth period, which is during the autumn season. It occurs from mid-August to mid-December. Therefore, changes in values for vegetation indices during this period are essential for understanding the effect of drought and its consequences. Other factors that may hinder growth during this period are pests, popularly known as Sal borer (Hoplocerambix spinicornis) (Rahman et al. 2010). Nonetheless, it is due to the impact of drought that weakens trees and aggravates the effect of pests.

Anomaly in NDVI values from the averaged median values from 2000 to 2020 for drought years with positive values indicating worsening conditions

Anomaly in EVI values from the averaged median values from 2000 to 2020 for drought years with positive values indicating worsening conditions
Anomaly values have spread out over the years and are mainly concentrated during the middle of the year, which is the growing season for Shorea robusta. This is important as it may lead to stunted growth for the forest and reduce mortality. It provides a much-needed understanding between forest vitality and drought, but ecological factors, along with forest insects and diseases, are also necessary to be examined (Kolb et al. 2016). Forest pest loss has been observed to be drought-driven, as forest resistance to drought decreases (Hendrik and Maxime 2017). Modelling of drought which takes into account the type of pest and the environment owing to climatic factors is complicated (Kolb et al. 2016). Irregularities aside, drought events that lead to massive losses in forest vitality are increasing globally (Steinkamp and Hickler 2015). Therefore, focused efforts have been made by this study to identify the timeframe for drought induced effects during drought years of the last two decades. A key observation is the increased anomaly during the growing seasons due to the increase in drought duration in recent years.
3.3 Lagged correlation
Significant results were observed after the onset of drought with respect to forest vitality. Correlation coefficients between the datasets are negative for the years 2013 and 2019. Positive correlation increases as time progresses and then returns to non-significance as the effect of drought ceases to exist. The increase implies that the effect of drought is being sensed by the ecology of the forest after the onset of drought. Furthermore, it indicates the resistance of forests to drought, although this depends on drought severity and duration. This is true for all years when there was a drought. Correlations with time lags tend to show a realistic representation since seasonal impacts were removed from the time series datasets (Verbesselt et al. 2010). This is because forest vitality follows a seasonal pattern unlike drought (Hänsel et al. 2019). Lag values are given in monthly format. All significant values are coloured green and red according to the positive and negative correlations, respectively. The ranges (0–5) are used to establish the months from the initial drought forward in time. Therefore, 0 is the onset of drought, 1 is 1 month later and so on. For example, if drought occurred in March, then 0 refers to March, 1 refers to April and so on (Table 2).
The individual random pixel time series exhibited similar characteristics to the lagged correlations and the averaged aggregated data are shown here. Monthly median average NDVI values over the entire period show no inherent trait due to changes in fluctuating drought patterns. Therefore, wrapping the data in individual drought year provides a better understanding of the lagged correlations (Zhao et al. 2020). Forest interaction with drought stress is complicated, but this form of relationship provides a better understanding of the data. Constant seasonal patterns are not exhibited by drought and thus, lagged values for certain times are irregular (Hänsel et al. 2019). The relationship between NDVI and EVI values and drought indices suggests that there is specific or immediate correlation for droughts of varying intensity and duration. The main obstacle that persists is when the effects of drought start to impact the forest ecosystem. Lagged correlation thus provides a better understanding of the situation. This is evident from the absence of spatiotemporal pattern in drought. The intensity and duration of drought, coupled with the spatiotemporal characteristics, present a highly volatile calculation scheme for lagged correlation. Similar results were obtained by Reinermann et al. (2019), using EVI and EVI lagged correlation for crops. Other factors that hamper the cross-correlation are the ecological status and growth period of the forest ecosystem. Anomalies in precipitation, soil moisture and root zone soil moisture are also viable factors that hinder the significance of time lagged correlation (Espinosa et al. 2019).
EVI lagged correlation coefficients show minor variation but similar results for forest vitality against drought. The values follow similar trends but occasionally show high variation in results, as in the case of year 2006 with lag 2 (Table 3). These values show how the forested area resists the onset of drought and eventually give in, although it is true that forests are more resistant than crops and do not show an immediate decrease in vitality given their access to root zone soil moisture (Devaranavadgi et al. 2010; Andivia et al. 2019).
4 Discussion
This study uses multi-decadal drought and vegetation indices to identify the relationship between them. Traditional methods of identifying such trends with in situ observations may be more accurate, but the sheer amount of time required to do so is problematic. Data has become more available due to advancements in remote sensing technologies combined with big data for earth observations (Gorelick et al. 2017). MODIS has been used for its revisit time (Xulu et al. 2018; Cammalleri and Vogt 2019; Cammalleri et al. 2019). The challenges that occur for such high temporal scale remote sensing data are the confidence in the prevalence of trees during the study. This was overcome by using the (Hansen et al. 2013) forest dataset that also provides data on forest pixels based on canopy cover (greater than 40% are taken here). The use of Landsat with a higher spatial resolution (30 m) can be an alternative, but it requires voluminous work for data consistency. This is because of the data collection from different sensors, particularly from Landsat 5, 7 and 8 (Banskota et al. 2014).
MODIS time series analysis for a very large temporal scale is therefore generally preferred over the Landsat images. Comparison of drought resilience cannot be done on a small temporal scale. Sentinel is not considered as data acquisition started in 2015 and thus unable to provide decadal evidence between drought stresses, lag and forest vitality. Time series analysis on big earth data offers a unique opportunity for spatiotemporal pattern recognition and relationship formation (Na et al. 2021). Drought impact on phenology based on time series (Máchová et al. 2018), trend and seasonal analysis (Verbesselt et al. 2010) and rainfall anomaly analysis via remote sensing have been extensively used for drought impact assessment (Winkler et al. 2017). Correlation with lags in time for forests has not been met with much attention (Gong et al. 2017). Seasonality-based study on trend analysis is increasing (Verbesselt et al. 2010; de Jong et al. 2011; Detsch et al. 2016), and this further necessitates the use of long-term time series analysis for measuring the impact of drought (de Jong et al. 2011).
Trees or forests tend to have higher resistance against drought because the roots can stretch to deep soil, thus reducing water scarcity. However, intense and prolonged drought can curtail this advantage (Buth et al. 2015). Additionally, drought weakens the capacity of trees to recuperate to a pre-drought level, making trees vulnerable to upcoming natural disasters (DeSoto et al. 2020; Bose et al. 2021). However, certain species, i.e. woody angiosperms, have shown compensatory recovery against droughts (Anderegg et al. 2015; Bose et al. 2021). A significant finding is that severe stresses affect the recovery time of Shorea robusta after subsequent droughts even if the drought is comparatively less severe. The year 2013 has had a deeper impact on forest response than the other years. The following years did not exhibit greater drought stress but had the recovery time shortened, as can be seen in Fig. 7 and Fig. 8. Similarly, since the impact of drought was short lived in 2018, it did not adversely affect the recovery time for the year 2019. Drought stress thus has a significant impact on forests even after it has dissipated and can even cause permanent damage. Identifying such areas where there is permanent damage is not a viable option due to the low resolution of satellite imageries used in this study.
Another important finding of this study is that severe drought has an immediate impact on forest vitality, as evidenced by the high correlation coefficients of NDVI and EVI against PDSI and SPEI (1-month and 3-month timescales). This is in accordance with similar studies that focused on specific severe droughts for forest mortality (Clark et al. 2016; Byer and Jin 2017; Zou et al. 2020). The real value brought by this study is the lag response time for droughts of varying intensities based on time series analysis of multi-decadal data on drought and forest. The lag times (months) for low drought severity are 3–4 months, 2–3 months for moderate droughts and 1 month for severe droughts with an immediate impact on forest vitality. Studies of this temporal scale have been conducted for crops (Tenhumberg et al. 2018; Na et al. 2021), but are limited for forests (Xulu et al. 2018; Reinermann et al. 2019).
The impact of drought and post-drought growth recovery varies across tree species (Peltier et al. 2016), which also depends on drought characteristics, i.e. duration, intensity, frequency and seasonality (Peltier and Ogle 2019; Bose et al. 2020). In this paper, we have accounted for the impact of drought duration, intensity and frequency on Shorea robusta. The drought intensity has been found to be 0.95 for the entire period (2000–2020). Additionally, the timing and seasonality of drought significantly impact the forest’s recovery. Huang et al. (2018) found that trees were not able to recover from summer season (March–May) drought during a 3-year post-drought period (see also Camarero et al. 2015; Gavinet et al. 2019). Previous studies on the impact of climatic factors on Shorea robusta in Bangladesh also indicate that precipitation is the key determinant of height and canopy area (Powers et al. 2009; Shishir et al. 2020). Our findings are similar to previous studies that found negative impact on forest vitality because of summer drought. The years 2006 and 2013 particularly have seen an inverse relationship between NDVI and precipitation due to the impact of drought. The recovery time, therefore, is elongated even during no drought periods.
In line with the discussion, climate change is hypothesized to worsen the drought condition in Bangladesh, resulting in the mass extinction of forest areas (Wiens et al. 2009; Barrows et al. 2010). Seasonal drought happens on a regular basis in the country. The consequent outcome is the low levels of precipitation and high temperatures during the extended summer seasons (Murad and Islam 2011; Sultana et al. 2021). Additionally, a delayed arrival and a shorter monsoon are expected due to climate change (Habiba and Shaw 2013). This drastic change in climatic conditions can potentially alter the distribution of species and ecosystems. Effective conservation policies and adaptive management practices thus require predicting tree-specific impacts of drought due to climate change (Pacifici et al. 2015; Sohel et al. 2017; Shishir et al. 2020). The time series data of drought as represented by PDSI and SPEI clearly show the impact of drought with varying intensities. Trend lines presented along with correlation between the indices illustrate that even with less drought intensity, forests in the study area are unable to recover quickly.
Finally, increasing drought intensity reduces tree-ring width, eventually decreasing the carbon storage capacity of trees or forests (Islam et al. 2019). Climate change-induced weather variability and the consequent depletion of trees’ carbon sequestration capacity are well documented in literature (Spannl et al. 2016). Along with the natural and climatic variables, anthropogenic pressures such as urbanization, rapidly expanding agriculture and plantations through invasive species, illegal logging, pollution and poor management practices are threatening the survival of Sal forests (Rahman et al. 2010). The local indigenous community is heavily dependent on the ecosystem services of the forest. This paper thus calls for reversing the vicious cycle of Sal Forest depletion. Identifying the existing inventory, as well as critical inventory, minimizing the anthropogenic pressures, and land reclamation by planting Shorea robusta are some of the noted forest revitalization strategies for forest managers and decision makers (Shishir et al. 2020).
5 Conclusion
This study has attempted to explore the relationship between drought and vegetation indices using remotely sensed data. The key results include the inverse relationship between the onset of drought and its impact on forests, which is dependent on the drought intensity. The stronger the drought, the less time it takes to impact forests. The lack of in situ analysis is an important aspect that further showcases the viability of remotely sensed data for monitoring drought effects in the study area. An inverse relationship between the lag and the intensity of drought has been observed, which decreases the lag time as drought duration and severity are increased. The intensity of drought over the entire temporal range was found to be 0.95. This is concerning even with the deeper root system of Shorea robusta. A lag time of more than 2 months will further affect the forest’s vitality. Therefore, subsequent drought events with lower intensity will also greatly impact the forest ecosystem. The methodological approach of this study offers an efficient technique for drought impact estimation on forests and thus can be a handy tool for policy makers.
One of the major limitations of this study is the lack of in situ analysis. The observations presented here are based on the selected drought indices, which do not consider ecological factors such as forest depletion caused by pests and beetles. Additionally, we fail to address the socio-economic factors of forest management.
Future research can focus on spatial analysis, such as cluster analysis, to distinguish areas affected by drought stress. The clusters can be based on overall drought stress and recovery time. The lag time calculated in this study can be used to further enhance the above analyses. However, this will require images with higher spatial resolution for better accuracy and precision to distinguish between the different clusters. Furthermore, integration of other indices, such as the Normalized Difference Water Index (NDWI), can be used to demonstrate plants’ water status. The root zone and surface soil moisture are inputs that can be added to scale the methodology presented in this study for different types of forests. Information regarding the interaction between different types of forests and drought can enhance drought recuperation strategies worldwide.
Data availability
The datasets used in the study are available from the corresponding author upon reasonable request.
Code availability
The codes used in the study are available from the corresponding author upon reasonable request.
Notes
Normalized Difference Vegetation Index (NDVI), Temperature Condition Index (TCI), Vegetation Condition Index (VCI), Vegetation Drought Response Index (VegDRI), Vegetation Health Index (VHI), Water Requirement Satisfaction Index (WRSI) and Geo-spatial WRSI, Normalized Difference Water Index (NDWI) and Land Surface Water Index (LSWI) and Soil Adjusted Vegetation Index (SAVI).
References
Abatzoglou JT, Dobrowski SZ, Parks SA, Hegewisch KC (2018) TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958-2015. Sci Data 5. https://doi.org/10.1038/sdata.2017.191
Adnan S (1993) Living without floods : lessons from the drought of 1992. Research & Advisory Services, Dhaka
Ahmad R (2021) Bangladesh among top 5 disaster-prone countries in Asia-Pacific | Dhaka Tribune. https://www.dhakatribune.com/bangladesh/environment/2021/03/08/bangladesh-among-top-5-disaster-prone-countries-in-asia-pacific. Accessed 20 Aug 2021
Alam M, Furukawa Y, … SS-IF, 2008 undefined Sustainability of Sal (Shorea robusta) forest in Bangladesh: past, present and future actions. ingentaconnect.com
Alley WM (1984) The Palmer Drought Severity Index: limitations and assumptions. J Clim Appl Meteorol 23:1100–1109. https://doi.org/10.1175/1520-0450(1984)023%3c1100:TPDSIL%3e2.0.CO;2
Anderegg WRL, Schwalm C, Biondi F, et al (2015) Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science (80- ) 349:528–532. https://doi.org/10.1126/science.aab1833
Andivia E, Zuccarini P, Grau B et al (2019) Rooting big and deep rapidly: the ecological roots of pine species distribution in southern Europe. Trees - Struct Funct 33:293–303. https://doi.org/10.1007/s00468-018-1777-x
Arfin Khan MAS, Uddin MB, Uddin MS et al (2007) Distribution and status of forests in the tropics: Bangladesh perspective. Proc Natl Acad Sci U S A 44:145–153
Banskota A, Kayastha N, Falkowski MJ et al (2014) Forest monitoring using Landsat time series data: a review. Can J Remote Sens 40:362–384
Barrows CW, Rotenberry JT, Allen MF (2010) Assessing sensitivity to climate change and drought variability of a sand dune endemic lizard. Biol Conserv 143:731–736. https://doi.org/10.1016/j.biocon.2009.12.013
Begueria S, Vicente-Serrano SM (2017) SPEI: calculation of the Standardised Precipitation-Evapotranspiration Index. R package version 1.7. R-Package
Bivand R, Tim K, Rowlingson B (2021) Package “rgdal” Title Bindings for the “Geospatial” Data Abstraction Library
Bose AK, Gessler A, Bolte A et al (2020) Growth and resilience responses of Scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob Chang Biol 26:4521–4537. https://doi.org/10.1111/gcb.15153
Bose AK, Scherrer D, Camarero JJ, et al (2021) Climate sensitivity and drought seasonality determine post-drought growth recovery of Quercus petraea and Quercus robur in Europe. Sci Total Environ 784. https://doi.org/10.1016/j.scitotenv.2021.147222
Brunner I, Herzog C, Dawes MA et al (2015) How tree roots respond to drought. Front Plant Sci 6:1–16. https://doi.org/10.3389/fpls.2015.00547
Buth M, Kahlenborn W, Savelsberg J, et al (2015) Vulnerabilität Deutschlands gegenüber dem Klimawandel
Byer S, Jin Y (2017) Detecting drought-induced tree mortality in Sierra Nevada forests with time series of satellite data. Remote Sens 9:14–17. https://doi.org/10.3390/rs9090929
Camarero JJ, Franquesa M, Sangüesa-Barreda G (2015) Timing of drought triggers distinct growth responses in holm oak: implications to predict warming-induced forest defoliation and growth decline. Forests 6:1576–1597. https://doi.org/10.3390/f6051576
Cammalleri C, Vogt JV (2019) Non-stationarity in MODIS fAPAR time-series and its impact on operational drought detection. Int J Remote Sens 40:1428–1444. https://doi.org/10.1080/01431161.2018.1524603
Cammalleri C, Verger A, Lacaze R, Vogt JV (2019) Harmonization of GEOV2 fAPAR time series through MODIS data for global drought monitoring. Int J Appl Earth Obs Geoinf 80:1–12. https://doi.org/10.1016/j.jag.2019.03.017
Christensen JH, Hewitson B, Busuioc A, et al (2007) Chapter 11: regional climate projections. Clim Chang 2007 Phys Sci Basis Contrib Work Gr I to Fourth Assess Rep Intergov Panel Clim Chang 847–940
Clark JS, Iverson L, Woodall CW et al (2016) The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States. Glob Chang Biol 22:2329–2352
de Jong R, de Bruin S, de Wit A et al (2011) Analysis of monotonic greening and browning trends from global NDVI time-series. Remote Sens Environ 115:692–702. https://doi.org/10.1016/j.rse.2010.10.011
Deshayes M, Guyon D, Jeanjean H et al (2006) The contribution of remote sensing to the assessment of drought effects in forest ecosystems. Ann for Sci 63:579–595
DeSoto L, Cailleret M, Sterck F, et al (2020) Low growth resilience to drought is related to future mortality risk in trees. Nat Commun 11. https://doi.org/10.1038/s41467-020-14300-5
Detsch F, Otte I, Appelhans T et al (2016) Seasonal and long-term vegetation dynamics from 1-km GIMMS-based NDVI time series at Mt. Kilimanjaro. Tanzania Remote Sens Environ 178:70–83. https://doi.org/10.1016/j.rse.2016.03.007
Devaranavadgi SB, Patil SB, Jambagi MB et al (2010) Influence of various potting mixtures on germination and root length of tree species in vertisols of northern Karnataka. Karnataka J Agric Sci 23:663–664
Dhar PP, Mridha MAU (2006) Biodiversity of arbuscular mycorrhizal fungi in different trees of madhupur forest, Bangladesh. J for Res 17:201–205. https://doi.org/10.1007/s11676-006-0047-8
Didan K (2015) MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid V006. NASA EOSDIS Land Processes DAAC USGS 5:2002–2015
Elhag KM, Zhang W (2018) Monitoring and assessment of drought focused on its impact on sorghum yield over Sudan by using meteorological drought indices for the period 2001–2011. Remote Sens 10:1–21. https://doi.org/10.3390/rs10081231
Espinosa LA, Portela MM, Rodrigues R (2019) Spatio-temporal variability of droughts over past 80 years in Madeira Island. J Hydrol Reg Stud 25:100623. https://doi.org/10.1016/j.ejrh.2019.100623
Garrity SR, Allen CD, Brumby SP et al (2013) Quantifying tree mortality in a mixed species woodland using multitemporal high spatial resolution satellite imagery. Remote Sens Environ 129:54–65. https://doi.org/10.1016/j.rse.2012.10.029
Gavinet J, Ourcival JM, Limousin JM (2019) Rainfall exclusion and thinning can alter the relationships between forest functioning and drought. New Phytol 223:1267–1279. https://doi.org/10.1111/nph.15860
Gazol A, Camarero JJ, Anderegg WRL, Vicente-Serrano SM (2017) Impacts of droughts on the growth resilience of Northern Hemisphere forests. Glob Ecol Biogeogr 26:166–176. https://doi.org/10.1111/geb.12526
Gazol A, Camarero JJ, Sánchez-Salguero R et al (2020) Drought legacies are short, prevail in dry conifer forests and depend on growth variability. J Ecol 108:2473–2484. https://doi.org/10.1111/1365-2745.13435
Gong Z, Zhao S, Gu J (2017) Correlation analysis between vegetation coverage and climate drought conditions in North China during 2001–2013. J Geogr Sci 27:143–160. https://doi.org/10.1007/s11442-017-1369-5
Gorelick N, Hancher M, Dixon M et al (2017) Google Earth Engine: planetary-scale geospatial analysis for everyone. Remote Sens Environ 202:18–27. https://doi.org/10.1016/j.rse.2017.06.031
Gustafson EJ, Sturtevant BR (2013) Modeling forest mortality caused by drought stress: implications for climate change. Ecosystems 16:60–74. https://doi.org/10.1007/s10021-012-9596-1
Habiba U, Shaw R (2013) Drought scenario in Bangladesh. Community, Environ Disaster Risk Manag 13:213–245. https://doi.org/10.1108/S2040-7262(2013)0000013016
Hänsel S, Ustrnul Z, Łupikasza E, Skalak P (2019) Assessing seasonal drought variations and trends over Central Europe. Adv Water Resour 127:53–75. https://doi.org/10.1016/j.advwatres.2019.03.005
Hansen MC, Potapov P V., Moore R, et al (2013) High-resolution global maps of 21st-century forest cover change. Science (80- ) 342:850–853. https://doi.org/10.1126/science.1244693
Haroon MA, Zhang J, Yao F (2016) Drought monitoring and performance evaluation of MODIS-based drought severity index (DSI) over Pakistan. Nat Hazards 84:1349–1366. https://doi.org/10.1007/s11069-016-2490-y
Hartmann H, Adams HD, Anderegg WRL et al (2015) Research frontiers in drought-induced tree mortality: crossing scales and disciplines. New Phytol 205:965–969. https://doi.org/10.1111/nph.13246
Hendrik D, Maxime C (2017) Assessing drought-driven mortality trees with physiological process-based models. Agric for Meteorol 232:279–290. https://doi.org/10.1016/j.agrformet.2016.08.019
Hijmans RJ, van Etten J (2014) raster: geographic data analysis and modeling. R package
Huang M, Wang X, Keenan TF, Piao S (2018) Drought timing influences the legacy of tree growth recovery. Glob Chang Biol 24:3546–3559. https://doi.org/10.1111/gcb.14294
Islam M, Rahman M, Bräuning A (2019) Impact of extreme drought on tree-ring width and vessel anatomical features of Chukrasia tabularis. Dendrochronologia 53:63–72. https://doi.org/10.1016/j.dendro.2018.11.007
Islam MN, Uyeda H (2005) Comparison of TRMM 3B42 products with surface rainfall over Bangladesh. In: International Geoscience and Remote Sensing Symposium (IGARSS). pp 4112–4115
Kannenberg SA, Maxwell JT, Pederson N et al (2019) Drought legacies are dependent on water table depth, wood anatomy and drought timing across the eastern US. Ecol Lett 22:119–127
Kebłowski P, Welfe A (2004) The ADF-KPSS test of the joint confirmation hypothesis of unit autoregressive root. Econ Lett 85:257–263. https://doi.org/10.1016/j.econlet.2004.04.013
Kim Y, Lee SB, Yun H, et al (2017) A drought analysis method based on MODIS satellite imagery and AWS data. In: International Geoscience and Remote Sensing Symposium (IGARSS). pp 4862–4865
Kolb TE, Fettig CJ, Ayres MP et al (2016) Observed and anticipated impacts of drought on forest insects and diseases in the United States. For Ecol Manage 380:321–334. https://doi.org/10.1016/j.foreco.2016.04.051
Leuschner C, Meier IC (2018) The ecology of Central European tree species: trait spectra, functional trade-offs, and ecological classification of adult trees. Perspect Plant Ecol Evol Syst 33:89–103. https://doi.org/10.1016/j.ppees.2018.05.003
Máchová P, Trcková O, Cvrcková H (2018) Use of nuclear microsatellite loci for evaluating genetic diversity of selected populations of Picea abies (L.) Karsten in the Czech Republic. Forests 9. https://doi.org/10.3390/f9020091
Murad H, Islam AKMS (2011) Drought assessment using remote sensing and GIS in north-west region of Bangladesh. 3rd Int Conf Water Flood Manag 861–877
Na L, Na R, Bao Y, Zhang J (2021) Time-lagged correlation between soil moisture and intra-annual dynamics of vegetation on the Mongolian plateau. Remote Sens 13. https://doi.org/10.3390/rs13081527
National Drought Mitigation Center (2021) What is drought | National Drought Mitigation Center. https://drought.unl.edu/Education/DroughtIn-depth/WhatisDrought.aspx. Accessed 20 Aug 2021
Nielsen B (2006) Correlograms for non-stationary autoregressions. J R Stat Soc Ser B Stat Methodol 68:707–720. https://doi.org/10.1111/j.1467-9868.2006.00563.x
Ogaya R, Barbeta A, Başnou C, Peñuelas J (2015) Satellite data as indicators of tree biomass growth and forest dieback in a Mediterranean holm oak forest. Ann for Sci 72:135–144. https://doi.org/10.1007/s13595-014-0408-y
Ovenden TS, Perks MP, Clarke TK, et al (2021) Life after recovery: increased resolution of forest resilience assessment sheds new light on post-drought compensatory growth and recovery dynamics. J Ecol. https://doi.org/10.1111/1365-2745.13576
Pacifici M, Foden WB, Visconti P et al (2015) Assessing species vulnerability to climate change. Nat Clim Chang 5:215–225
Panuju DR, Paull DJ, Griffin AL, Trisasongko BH (2021) Mapping rice growth stages employing MODIS NDVI and ALOS AVNIR-2. In: Kumar P, Sajjad H, Chaudhary BS et al (eds) Remote sensing and GIScience. Springer International Publishing, Cham, pp 185–203
Pebesma E (2021) Simple features for R
Pei Z, Fang S, Wang L, Yang W (2020) Comparative analysis of drought indicated by the SPI and SPEI at various timescales in inner Mongolia, China. Water (Switzerland) 12. https://doi.org/10.3390/w12071925
Peltier DMP, Ogle K (2019) Legacies of more frequent drought in ponderosa pine across the western United States. Glob Chang Biol 25:3803–3816. https://doi.org/10.1111/gcb.14720
Peltier DMP, Fell M, Ogle K (2016) Legacy effects of drought in the southwestern United States: a multi-species synthesis. Ecol Monogr 86:312–326. https://doi.org/10.1002/ecm.1219
Potopová V, Štěpánek P, Možný M et al (2015) Performance of the standardised precipitation evapotranspiration index at various lags for agricultural drought risk assessment in the Czech Republic. Agric for Meteorol 202:26–38. https://doi.org/10.1016/j.agrformet.2014.11.022
Powers JS, Montgomery RA, Adair EC et al (2009) Decomposition in tropical forests: a pan-tropical study of the effects of litter type, litter placement and mesofaunal exclusion across a precipitation gradient. J Ecol 97:801–811. https://doi.org/10.1111/j.1365-2745.2009.01515.x
Rahman MM, Rahman MM, Guogang Z, Islam KS (2010) A review of the present threats to tropical moist deciduous Sal (Shorea robusta) forest ecosystem of central Bangladesh. Trop Conserv Sci 3:90–102. https://doi.org/10.1177/194008291000300108
Reinermann S, Gessner U, Asam S et al (2019) The effect of droughts on vegetation condition in Germany: an analysis based on two decades of satellite earth observation time series and crop yield statistics. Remote Sens 11:1783. https://doi.org/10.3390/rs11151783
Sáenz C, Litago J, Wiese K et al (2021) Drought periods identification in Ecuador between 2001 and 2018 using SPEI and MODIS data. Eng Proc 9:24. https://doi.org/10.3390/engproc2021009024
Sarker MSH (2021) Regional spatial and temporal variability of rainfall, temperature over Bangladesh and Northern Bay of Bengal. Environ Challenges 5:100309. https://doi.org/10.1016/j.envc.2021.100309
Serra-Maluquer X, Granda E, Camarero JJ et al (2021) Impacts of recurrent dry and wet years alter long-term tree growth trajectories. J Ecol 109:1561–1574. https://doi.org/10.1111/1365-2745.13579
Shahid S (2010) Recent trends in the climate of Bangladesh. Clim Res 42:185–193. https://doi.org/10.3354/cr00889
Shahid S (2011) Impact of climate change on irrigation water demand of dry season Boro rice in northwest Bangladesh. Clim Change 105:433–453. https://doi.org/10.1007/s10584-010-9895-5
Shahid S, Behrawan H (2008) Drought risk assessment in the western part of Bangladesh. Nat Hazards 46:391–413. https://doi.org/10.1007/s11069-007-9191-5
Shishir S, Mollah TH, Tsuyuzaki S, Wada N (2020) Predicting the probable impact of climate change on the distribution of threatened Shorea robusta forest in Purbachal, Bangladesh. Glob Ecol Conserv 24. https://doi.org/10.1016/j.gecco.2020.e01250
Singh KP, Kushwaha CP (2005) Paradox of leaf phenology: Shorea robusta is a semi-evergreen species in tropical dry deciduous forests in India. Curr Sci 88:1820–1824
Sohel SI, Akhter S, Ullah H et al (2017) Predicting impacts of climate change on forest tree species of Bangladesh: evidence from threatened Dysoxylum binectariferum (Roxb.) Hook.f. ex Bedd. (Meliaceae). Iforest 10:154–160. https://doi.org/10.3832/ifor1608-009
Spannl S, Volland F, Pucha D et al (2016) Climate variability, tree increment patterns and ENSO-related carbon sequestration reduction of the tropical dry forest species Loxopterygium huasango of Southern Ecuador. Trees - Struct Funct 30:1245–1258. https://doi.org/10.1007/s00468-016-1362-0
Spinoni J, Naumann G, Carrao H et al (2014) World drought frequency, duration, and severity for 1951–2010. Int J Climatol 34:2792–2804. https://doi.org/10.1002/joc.3875
Steinkamp J, Hickler T (2015) Is drought-induced forest dieback globally increasing? J Ecol 103:31–43. https://doi.org/10.1111/1365-2745.12335
Stoffer D (2014) astsa: applied statistical time series analysis. R package version 1.3
Sultana MS, Gazi MY, Mia MB (2021) Multiple indices based agricultural drought assessment in the northwestern part of Bangladesh using geospatial techniques. Environ Challenges 4:100120. https://doi.org/10.1016/j.envc.2021.100120
Svoboda M, Fuchs B (2017) Handbook of drought indicators and indices*. In: Integrated drought management tools and guidelines series. pp 155–208
Tan C, Yang J, Li M (2015) Temporal-spatial variation of drought indicated by SPI and SPEI in Ningxia Hui Autonomous Region, China. Atmosphere (basel) 6:1399–1421. https://doi.org/10.3390/atmos6101399
Tenhumberg B, Crone EE, Ramula S, Tyre AJ (2018) Time-lagged effects of weather on plant demography: drought and Astragalus scaphoides. Ecology 99:915–925. https://doi.org/10.1002/ecy.2163
Trapletti A, Hornik K, LeBaron B (2015) tseries: time series analysis and computational finance. R package version 0.10–34
Verbesselt J, Hyndman R, Newnham G, Culvenor D (2010) Detecting trend and seasonal changes in satellite image time series. Remote Sens Environ 114:106–115. https://doi.org/10.1016/j.rse.2009.08.014
Vicente-Serrano SM, Beguería S, López-Moreno JI (2010) A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. J Clim 23:1696–1718. https://doi.org/10.1175/2009JCLI2909.1
Vogel MM, Hauser M, Seneviratne SI (2020) Projected changes in hot, dry and wet extreme events’ clusters in CMIP6 multi-model ensemble. Environ Res Lett 15. https://doi.org/10.1088/1748-9326/ab90a7
Wiens JA, Stralberg D, Jongsomjit D et al (2009) Niches, models, and climate change: assessing the assumptions and uncertainties. Proc Natl Acad Sci 106:19729–19736. https://doi.org/10.1073/PNAS.0901639106
Wilhite DA, Sivakumar MVK, Pulwarty R (2014) Managing drought risk in a changing climate: the role of national drought policy. Weather Clim Extrem 3:4–13. https://doi.org/10.1016/j.wace.2014.01.002
Winkler K, Gessner U, Hochschild V (2017) Identifying droughts affecting agriculture in Africa based on remote sensing time series between 2000-2016: rainfall anomalies and vegetation condition in the context of ENSO. Remote Sens 9. https://doi.org/10.3390/rs9080831
Xulu S, Peerbhay K, Gebreslasie M, Ismail R (2018) Drought influence on forest plantations in Zululand, South Africa, using MODIS time series and climate data. Forests 9:1–15. https://doi.org/10.3390/f9090528
Yevjevich V (1969) An objective approach to definitions and investigations of continental hydrologic droughts. J Hydrol 7:353. https://doi.org/10.1016/0022-1694(69)90110-3
Zeileis A, Grothendieck G (2005) Zoo: S3 infrastructure for regular and irregular time series. J Stat Softw 14:1–27. https://doi.org/10.18637/jss.v014.i06
Zhao A, Yu Q, Feng L et al (2020) Evaluating the cumulative and time-lag effects of drought on grassland vegetation: a case study in the Chinese Loess Plateau. J Environ Manage 261:110214. https://doi.org/10.1016/j.jenvman.2020.110214
Zou L, Cao S, Zhao A, Sanchez-Azofeifa A (2020) Assessing the temporal response of tropical dry forests to meteorological drought. Remote Sens 12:7–9. https://doi.org/10.3390/rs12142341
Author information
Authors and Affiliations
Contributions
Tanmoy Chakraborty: conceptualization, data curation, methodology, formal analysis, software, visualization, writing—original draft. Showmitra Kumar Sarkar: investigation, visualization, software, writing—original draft. Md. Manjur Morshed: conceptualization, supervision, writing—original draft, writing—review and editing.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Chakraborty, T., Sarkar, S.K. & Morshed, M.M. Big data and remote sensing for multi-decadal drought impact assessment on Shorea robusta. Theor Appl Climatol 148, 1587–1602 (2022). https://doi.org/10.1007/s00704-022-04019-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00704-022-04019-2