
Abstract
Metamorphosis in amphibians is a fascinating phenomenon that offers physiologists a number of opportunities to study aspects of tissue growth and lysis in animals. In larval amphibians, tissues are formed early on during larval growth, then are vastly reorganized and broken down during metamorphosis, and the effects of this phenomenon on the relative distribution of white blood cells in circulation (i.e., leukocyte profiles) was first examined in the 1920s. Nearly a century later, there has been little work to continue this research avenue, although our knowledge of the roles of certain leukocyte types has improved. Moreover, recent interest in counts of leukocytes in amphibians for environmental monitoring purposes emphasizes the need to understand how white blood cells naturally vary throughout larval life. In this study, I examined blood smears from wild-caught larval and adult bullfrogs (Rana catesbeiana) of varying developmental stages and quantified the proportions and abundance (estimated from blood smears) of all leukocyte types at each stage. Lymphocytes were the most common cell found and appeared most abundant during the early growth phases of larvae. Neutrophils were the second most common cell and were also associated most with earlier phases. Eosinophils appeared directly associated with metamorphosis, generally increasing in abundance to a peak at metamorphic climax. Monocytes were also found most frequently in individuals experiencing metamorphic climax. Results in this study concerning eosinophils are consistent with those of urodeles and point to their role in the breakdown of tissues, which may resemble their role in modulating inflammation responses.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Amphibian larval growth and metamorphosis have long been subjects of research by animal physiologists (Hayes 1997), especially with respect to their effects on blood cells such as erythrocytes (Jordan and Speidel 1923; Hollyfield 1966; Maniatis and Ingram 1971; Wakahara and Yamaguchi 2001) and leukocytes (Jordan and Speidel 1922, 1924). Part of the long-standing interest in these topics no doubt stems from the fact that these two phases have opposing effects on tissues of these animals; during the earlier growth phases, tissues such as legs are being formed (in anurans) and body size increases, but later metamorphosis causes a massive tissue restructuring and breakdown (such as tail resorption), leading to a reduction in overall body size. This unique phenomenon in amphibians gives animal physiologists an opportunity to understand how blood cells are involved in both processes.
Although the effect of metamorphosis on the abundance of different leukocyte types in circulation was first studied nearly a century ago (Jordan and Speidel 1922, 1924), today, this work remains as one of the few studies of the subject. Moreover, even though these studies were largely observational, they yielded important insights into the role of these cells during tissue growth and breakdown in animals. For example, Jordan and Speidel’s observations indicated that lymphocytes were more important during tissue growth and that neutrophils and eosinophils both played important roles later in the breakdown, or lysis, of tissues. Neutrophils, which are known as phagocytic cells involved in non-specific pathogen defense and inflammation (Jain 1993; Thrall 2004), were suspected to have a role in tissue breakdown by producing “secretions” that furthered the process of tissue lysis (Jordan and Speidel 1924). Eosinophils are mostly known for their association with metazoan parasite defense (Kiesecker 2002), but during metamorphosis, these cells were also thought to give off “secretions” that aided in tissue lysis (Jordan and Speidel 1924). Although they did not know what these secretions were, Jordan and Speidel’s early speculations were surprisingly intuitive, since we now know that granulocytic leukocytes such as eosinophils produce a variety of chemical substances (via their granules) such as cytokines, chemokines, lipid mediators, and neuromodulators (Rothenberg and Hogan 2006), and these substances act to initiate and modulate the inflammatory response in tissues. We also now know that eosinophils can respond to tissue injury, infections, allergens, and tumors (Adamko et al. 2005; Rothenberg and Hogan 2006) besides protecting the body from parasites. Interestingly, in the only other study of metamorphosis-related changes in leukocytes of amphibians, eosinophils were also shown to be particularly abundant during induced metamorphosis of axolotls (Ambystoma mexicanum; Ussing and Rosenkilde 1995). However, since ambystomatid salamanders tend to have unusually high numbers of this cell type (Davis and Maerz 2008a, b), results from this genus may not be typical of other amphibians. As for the remaining two white blood cell types, Jordan and Speidel (1924) noted that basophils were most abundant during later stages of metamorphosis, though their function was unclear, and that monocytes (also a phagocytic cell) were also most abundant during the later stages of metamorphosis, presumably in response to the increased cellular debris left over from the tissue lysis.
While these qualitative observations are important, it should also be pointed out that they were made on frogs that were experimentally induced to metamorphose and that nearly all of the animals died before they completed metamorphosis (Jordan and Speidel 1922, 1924), which indicates that they were unnaturally stressed from the procedure. A more complete picture would be gained by examining wild larvae in various stages of natural metamorphosis. Interestingly though, a thorough literature review by the author indicates that the early work by Jordan and Speidel has rarely been followed, except for a study of blood cells of lab-reared axolotls (A. mexicanum, Ussing and Rosenkilde 1995), and for investigations of certain cell types (lymphocytes) in organ tissues of lab-reared Xenopus laevis (Rollins-Smith et al. 1997). This is despite a recent interest by ecologists and wildlife researchers in amphibian white blood cells (Forbes et al. 2006; Forson and Storfer 2006; Gervasi and Foufopoulos 2008) because of their value in health evaluation of these environmentally sensitive animals (Cabagna et al. 2005; Raffel et al. 2006; Barni et al. 2007). Indeed, modern investigations of amphibians are increasingly incorporating counts of some or all circulating white blood cells into their research goals, and some of these investigations involve examination of larval white blood cells (e.g., Kiesecker 2002; Barni et al. 2007). Because of this recent interest in amphibian larval leukocytes, there is now a pressing need to understand how numbers of circulating leukocytes naturally vary during larval growth and metamorphosis. The current project was initiated with the intent to address this question, by providing an updated and more quantitative evaluation of leukocyte changes during (anuran) larval growth and metamorphosis than the observations made by Jordan and Speidel nearly a century ago. The species studied was a model larval anuran (the bullfrog, Rana catesbeiana), and because the intent was to study the effects of natural metamorphosis, the study involved examination of wild-caught individuals that naturally ranged in development stages.
Materials and methods
Capturing and processing larvae
Bullfrog larvae were dip-netted from three local ponds around the University of Georgia campus in Clarke Co., GA in the month of June 2008. All larvae were transported to the lab within 20 min of capture where they were processed immediately. Processing involved anesthetizing them via immersion in a 1% solution of MS-222, then weighing with an electronic balance, and determining their developmental stage according to Gosner (1960). A blood sample was obtained by cutting the tip of the tail so that a small amount of blood welled from the cut, touching this blood directly to a clean microscope slide, and using a second slide to make a standard blood smear. For late-stage larvae with little tail remaining, animals were decapitated after anesthetizing and blood siphoned from the heart region with a microcapillary tube. Slides were air-dried, then later stained with Giemsa.
A set of adult bullfrogs were also hand-captured from the same ponds. These animals were processed in the field; they were each weighed with a portable electronic balance, then a blood sample was non-destructively obtained by puncturing the midline abdominal vein with a 27-gauge needle and siphoning 20 µl of blood with a microcapillary tube (Wright 2001). This sample was used to prepare a standard blood smear, which was air-dried and stained as before. All procedures were approved by the University of Georgia Institutional Animal Care and Use Committee.
Reading blood smears
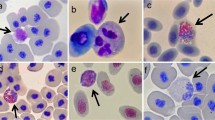
All blood smears were examined by the author under 1,000× (oil) with a standard light microscope, while blood cells were identified as neutrophils, lymphocytes, eosinophils, basophils, and monocytes following Hadji-Azimi et al. (1987) and Thrall (2004). Slides were viewed in a standard zigzag pattern, covering all parts of the blood smear, and all leukocytes were counted in each field of view until 100 cells were counted or 150 fields of view were reached. The proportion of each cell type and the estimated number per 2,000 erythrocytes were calculated following Davis et al. (2004) and Davis and Maerz (2008a). The number of red blood cells per field of view was estimated from examination of 50 fields (from five different animals), which showed an average of 47 cells (±7 SD) per field. With this value, plus the number of cells of each leukocyte per field of view, the estimated number of leukocytes per 2,000 red blood cells was calculated for each cell type. These values were used in the analyses of leukocyte abundance throughout larval development (below).
Data analysis
Development stages were paired to increase sample sizes, except for the final stage of development (46, froglets), which resulted in 12 groups (including adult frogs) for analysis. Analysis of variance was then used to compare estimated white blood cell numbers across these 12 stages, with each cell type examined separately. Cell data were log-transformed prior to analyses to approximate normal distributions. Analyses were conducted using Statistica 6.1 software (Statistica 2003).
Results
General results
A total of 83 individuals were sampled in this study, consisting of 70 larvae in various stages of development, six newly metamorphosed froglets (Gosner stage 46) and seven adult frogs (Table 1). Wet weights of each paired stage showed a bell-shaped curve typical of anuran larval development (Fig. 1). Larval body weights showed a period of growth until around stages 38–39, and weights declined thereafter. Adult body weight (not shown in Fig. 1) averaged 95.4 g (±17.6 SE).

Average wet weights (±1SE) of bullfrog larvae across all development stages (following Gosner 1960) examined in the current study. Sample sizes for each development stage shown in Table 1. Average weight of adult bullfrogs not shown (95.4 g ± 17.6 SE). Dashed line indicates approximate stage when metamorphosis starts
Leukocyte profiles
The relative breakdown of leukocyte types (i.e., leukocyte profiles) across development stages is presented in Table 1. In general, lymphocytes were the most abundant cell type in all bullfrogs examined, with an average of 72.7% of all white blood cells. The percentage of neutrophils varied more so, ranging from 7.5% to 25.6%. A higher percentage of neutrophils was seen in early stages (26–39) of development (i.e., those characterized by growth) than later stages (20.8% versus 12.1%, respectively). The percentage of eosinophils tended to increase throughout development stages until a peak at metamorphic climax, and a similar trend was noted in monocytes, though the increase began later in this cell type. The percentage of basophils tended to increase throughout development. Overall, the average leukocyte profile of larval and adult bullfrogs found in this study was not largely different from published values (of adults) for this species, which are presented at the bottom of Table 1 (Cathers et al. 1997).
Leukocyte numbers
Statistical comparisons of the estimated numbers of cells across the 12 development categories considered in this study showed significant variation in all leukocyte types, and most trends in abundance largely followed those seen in the proportions in Table 1. Lymphocyte numbers varied significantly throughout development (F 11,71 = 2.66, p = 0.007), peaking during stages 30–33 at 210 cells per 2,000 erythrocytes, then declining to approximately 120 cells for the remainder of larval development (Fig. 2a). The number of neutrophils varied throughout development stages (F 11,71 = 3.62, p < 0.001), with highest estimated counts during early stages (26–31), then somewhat low counts during subsequent stages, except for a small jump during stages 38–39 (Fig. 2b). There was a significant variation in estimated eosinophil counts (F 11,71 = 2.98, p = 0.003), and these showed a trend of gradual increase during development until a sharp peak at metamorphic climax (Fig. 2c). Estimated basophil counts varied significantly (F 11,71 = 4.17, p < 0.001) and showed a general increase throughout development to the adult stage (Fig. 2d). Finally, estimated counts of monocytes varied significantly (F 11,71 = 2.83, p = 0.004) and showed generally low abundance throughout most stages but a sharp increase in abundance during the climax stages of metamorphosis (Fig. 2e).

Changes in relative numbers of lymphocytes (a), neutrophils (b), eosinophils (c), basophils (d), and monocytes (e) in bullfrog larvae during development and metamorphosis (including adults). Means ± 1 SE shown for each cell type. Sample sizes for each development stage shown in Table 1. Dashed line indicates approximate stage when metamorphosis starts
Discussion
The results from this study provide a clear picture of how numbers of white blood cells change during development and metamorphosis in anuran amphibians, and several trends appear consistent with the early observations by Jordan and Speidel (1922, 1924). For example, these data indicate that lymphocytes are most abundant during the growth phase of larval development and appear to have little association with metamorphic climax (Table 1, Fig. 2a). The same could be said of neutrophils, except for an unusual spike in (estimated) numbers of this cell at the peak time of larval growth (stages 38–39). Besides that increase, these cells were actually least abundant during the metamorphic period. The high numbers of neutrophils during the early larval periods (Table 1) however are intriguing. Given that this cell is not thought to be involved in tissue growth (Jordan and Speidel 1922, 1924) and since it is one of the first cells to respond to sites of infection (Jain 1993; Thrall 2004), it may be that anuran larvae require more non-specific immune investment during these stages than during later ones, although this idea is merely a speculation and further work would be needed to confirm it.
This study also demonstrated that eosinophils are clearly involved in anuran metamorphosis, as has been shown in urodeles (Ussing and Rosenkilde 1995). Numbers and proportions of this white blood cell type increased during the metamorphic stages (Table 1, Fig. 2c), peaked at metamorphic climax, then declined to presumably baseline levels in metamorphic and adult frogs. The consistency of these results with the observations of Jordan and Speidel (1922, 1924) and the data from urodels (Ussing and Rosenkilde 1995) speaks to its universal nature among all amphibians. These data also lend support to the idea that these cells act to further the process of tissue lysis during metamorphosis, since they appear most abundant at the time when tissues are being heavily remodeled (metamorphic climax). Jordan and Speidel believed that this cell produced substances that aided in the breakdown of tissues, thereby acting as a precursor to phagocytosis by monocytes. Indeed, the current study did find evidence that monocytes are involved in the final stages of metamorphosis (Fig. 2e) and presumably act to rid the body of cellular debris, which is consistent with Jordan and Speidel’s suggestion. However, today, our understanding of the function of eosinophils has broadened, and we now know that it produces a number of chemicals to initiate and modulate the immune and inflammation response (Adamko et al. 2005; Rothenberg and Hogan 2006). It may be that the lysis of tissues during metamorphosis behaves in a way similar to the inflammation response, and the eosinophil acts to modulate this process via secretion of chemicals from its granules.
Data from this study suggest that basophils do not appear to be involved in metamorphosis per se but, instead, show a gradual increase in abundance over time during development, with highest numbers evident among adults. This trend perhaps reflects the timing of their formation and entrance in circulation of developing larvae more so than a direct association with metamorphosis. The role of these cells in the innate immune system is not clear (Turner 1988), although for reasons unknown, this cell does vary greatly in abundance among amphibian species from as much as 57% of leukocytes in the Japanese newt (Cynops pyrrhogaster; Pfeiffer et al. 1990) to 1% in the Colorado river toad (Bufo alvarius; Cannon and Cannon 1979).
A final observation from this work that is of interest is the fact that four of the five leukocyte types appeared to increase in abundance, either slightly or very pronounced, between stages 28 and 33 for some unknown reason. During this point of development, the larva is not only slowly growing hind legs but also experiencing rapid growth in overall body size (Fig. 1). Are these increases a response to the rapid growth? Or are they a bolstering of the innate immune system during a time of increased food consumption and presumably increased activity? Such questions clearly warrant further study, and they highlight the large gaps that still remain in our knowledge of amphibian metamorphosis, especially as it relates to blood cells.
References
Adamko DJ, Odemuyiwa SO, Vethanayagam D, Moqbel R (2005) The rise of the phoenix: the expanding role of the eosinophil in health and disease. Allergy 60:13–22 doi:10.1111/j.1398-9995.2005.00676.x
Barni S, Boncompagni E, Grosso A, Bertone V, Freitas I, Fasola M et al (2007) Evaluation of Rana esculenta blood cell response to chemical stressors in the environment during the larval and adult phases. Aquat Toxicol 81:45–54 doi:10.1016/j.aquatox.2006.10.012
Cabagna MC, Lajmanovich RC, Stringhini G, Sanchez-Hernandez JC, Peltzer PM (2005) Hematological parameters of health status in the common toad Bufo arenarum in agroecosystems of Santa Fe Province, Argintina. Appl Herpetology 2:373–380 doi:10.1163/157075405774483085
Cannon MS, Cannon AM (1979) The blood leukocytes of Bufo alvarius: a light, phase-contrast, and histochemical study. Can J Zool 57:314–322
Cathers T, Lewbart GA, Correa M, Stevens JB (1997) Serum chemistry and hematology values for anesthetized American bullfrogs (Rana catesbeiana). J Zoo Wildl Med 28:171–174
Davis AK, Maerz JC (2008a) Comparison of hematological stress indicators in recently captured and captive paedomorphic mole salamanders, Ambystoma talpoideum. Copeia 2008:613–617
Davis AK, Maerz JC (2008b) Sex-related differences in hematological stress indices of breeding, paedomorphic mole salamanders. J Herpetol 42:197–201 doi:10.1670/06–277R2.1
Davis AK, Cook KC, Altizer S (2004) Leukocyte profiles of House Finches with and without mycoplasmal conjunctivitis, a recently emerged bacterial disease. EcoHealth 1:362–373 doi:10.1007/s10393-004-0134-2
Forbes MR, McRuer DL, Shutler D (2006) White blood cell profiles of breeding American toads (Bufo americanus) relative to sex and body size. Comp Clin Pathol 15:155–159 doi:10.1007/s00580-006-0623-5
Forson D, Storfer A (2006) Atrazine increases ranavirus susceptibility in the tiger salamander, Ambystoma tigrinum. Ecol Appl 16:2325–2332 doi:10.1890/1051-0761(2006)016[2325:AIRSIT]2.0.CO;2
Gervasi SS, Foufopoulos J (2008) Costs of plasticity: responses to desiccation decrease post-metamorphic immune function in a pond-breeding amphibian. Funct Ecol 22:100–108
Gosner KL (1960) A simplified table for staging anuran embryos and larvae. Herpetologica 16:183–190
Hadji-Azimi I, Coosemans V, Canicatti C (1987) Atlas of adult Xenopus laevis laevis hematology. Dev Comp Immunol 11:807–874 doi:10.1016/0145-305X(87)90068-1
Hayes TB (1997) Amphibian metamorphosis: an integrative approach. Am Zool 37:121–123
Hollyfield JG (1966) Erythrocyte replacement at metamorphosis in the frog Rana pipiens. J Morphol 119:1–6 doi:10.1002/jmor.1051190102
Jain NC (1993) Essentials of veterinary hematology. Blackwell, Philadelphia
Jordan HE, Speidel CC (1922) Leukocytes in relation to the mechanism of thyroid-accelerated metamorphosis in the larval frog. Proc Soc Exp Biol Med 20:380–383
Jordan HE, Speidel CC (1923) Blood cell formation and distribution in relation to the mechanism of thyroid-accelerated metamorphosis in the larval frog. J Exp Med 38:529–543 doi:10.1084/jem.38.5.529
Jordan HE, Speidel CC (1924) The behavior of the leucocytes during coincident regeneration and thyriod-induced metamorphosis in the frog larva, with a consideration of growth factors. J Exp Med 40:1–11 doi:10.1084/jem.40.1.1
Kiesecker JM (2002) Synergism between trematode infection and pesticide exposure: a link to amphibian deformities in nature? Proc Natl Acad Sci U S A 99:9900–9904 doi:10.1073/pnas.152098899
Maniatis GM, Ingram VM (1971) Erythropoiesis during amphibian metamorphosis 1. Site of maturation of erythrocytes in Rana catesbeiana. J Cell Biol 49:372 doi:10.1083/jcb.49.2.372
Pfeiffer CJ, Pyle H, Asashima M (1990) Blood cell morphology and counts in the Japanese newt (Cynops pyrrhogaster). J Zoo Wildl Med 21:56–64
Raffel TR, Rohr JR, Kiesecker JM, Hudson PJ (2006) Negative effects of changing temperature on amphibian immunity under field conditions. Funct Ecol 20:819–828 doi:10.1111/j.1365-2435.2006.01159.x
Rollins-Smith LA, Barker KS, Davis AT (1997) Involvement of glucocorticoids in the reorganization of the amphibian immune system at metamorphosis. Dev Immunol 5:145–152 doi:10.1155/1997/84841
Rothenberg ME, Hogan SP (2006) The eosinophil. Annu Rev Immunol 24:147–174 doi:10.1146/annurev.immunol.24.021605.090720
Statistica. 2003. Statistica version 6.1, Statsoft Inc.
Thrall MA (2004) Hematology of amphibians. In: Thrall MA, Baker DC, Lassen ED (eds) Veterinary hematology and clinical chemistry: text and clinical case presentations. Lippincott Williams & Wilkins, Philadelphia, PA
Turner RJ (1988) Amphibians. In: Rawley AF, Ratcliffe NA (eds) Vertebrate blood cells. Cambridge University Press, Cambridge, pp 129–209
Ussing AP, Rosenkilde P (1995) Effect of induced metamorphosis on the immune system of the Axolotl, Ambystoma mexicanum. Gen Comp Endocrinol 97:308–319 doi:10.1006/gcen.1995.1031
Wakahara M, Yamaguchi M (2001) Erythropoiesis and conversion of RBCs and hemoglobins from larval to adult type during amphibian development. Zool Sci 18:891–904 doi:10.2108/zsj.18.891
Wright KM (2001) Amphibian hematology. In: Wright KM, Whitaker BR (eds) Amphibian medicine and captive husbandry. Krieger, Malabar, FL, pp 129–146
Acknowledgments
Jennifer Yates and Stephanie Kern helped with catching adult bullfrogs. John Maerz provided logistic support, and financial support came from the D.B. Warnell School of Forestry and Natural Resources, at the University of Georgia.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Davis, A.K. Metamorphosis-related changes in leukocyte profiles of larval bullfrogs (Rana catesbeiana). Comp Clin Pathol 18, 181–186 (2009). https://doi.org/10.1007/s00580-008-0773-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00580-008-0773-8