
Abstract
The aim of the present study was to estimate the heat transfer through cutaneous and respiratory evaporation of dairy cows raised in tropical ambient conditions using simple environmental and physiological measures. Twenty-six lactating crossbred cows (7/8 Holstein-Gir) were used, 8 predominantly white and 18 predominantly black. The environmental variables air temperature, relative humidity, black globe temperature, and wind speed were measured. Respiratory rate and coat surface temperature were measured at 0700, 0900, 1100, 1300, and 1500 h. The environmental and physiological data were used to estimate heat loss by respiratory (ER) and cutaneous evaporation (EC). Results showed that there was variation (P < 0.01) for respiratory rate depending on the times of the day. The highest values were recorded at 1100, 1300, and 1500 h, corresponding to 66.85 ± 10.20, 66.98 ± 7.80, and 65.65 ± 6.50 breaths/min, respectively. Thus, the amount of heat transferred via respiration ranged from 19.21 to 29.42 W/m2. There was a variation from 31.6 to 38.8 °C for coat surface temperature; these values reflected a range of 55.52 to 566.83 W/m2 for heat transfer via cutaneous evaporation. However, throughout the day, the dissipation of thermal energy through the coat surface accounted for 87.9 % total loss of latent heat, and the remainder (12.1 %) was via the respiratory tract. In conclusion, the predictive models based on respiratory rate and coat surface temperature may be used to estimate the latent heat loss in dairy cows kept confined in tropical ambient conditions.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Ruminants with excess thermal load use evaporation as the main thermoregulatory mechanism, especially under high air temperatures. Thus, the ability of an animal to withstand hot environments is proportional to its ability to dissipate the latent heat by evaporation, either by sweat on the skin (McLean 1963) or from the respiratory system (Silva et al. 2002; Maia et al. 2005b).
Several studies have evaluated the effects of the thermal environment on heat dissipation in cattle (McLean 1963; Finch 1985; Silva 1999; Maia et al. 2005a, b; Maia et al. 2008), sheep (Hofmeyr et al. 1969; Silva et al. 2002; Silva and Starling 2003) and goats (Ligeiro et al. 2006). Most of these experiments were conducted under controlled conditions, using direct methods in the evaluation of heat loss by respiratory and cutaneous evaporation, which involve the use of masks and respirometry chambers, ventilated capsules, among other methods. However, estimating the animal heat balance in field has been a challenge for studies in animal biometereorology.
The use of indirect methods to estimate the thermal energy transfer rate would be interesting especially for application in field studies. Maia et al. (2005a) and Silva and Maia (2011) developed equations that are able to predict the latent heat transfer in Holstein cows in a tropical environment, through simple measures such as coat surface temperature and respiratory rate. Such measures are relatively easy to obtain in field conditions with the use of infrared thermometer and visual observation of respiration (Gaughan 2000). Moreover, it is a non-invasive method, without the need for animal containment, which ensures greater confidence in the physiological variables measured.
Research focused on estimating heat balance in Holstein cattle in tropical environments is scarce, especially in the field. Furthermore, the use of these prediction equations may be validated to estimate the latent heat transfer in Holstein cows raised in tropical semiarid regions of Brazil. Thus, the aim of this study was to estimate the heat transfer through cutaneous and respiratory evaporation of dairy cows raised in tropical ambient conditions using simple environmental and physiological measures.
Materials and methods
Animals and handling
Twenty-six lactating crossbred cows (7/8 Holstein-Gir) were used, 18 predominantly black (days in milk (DIM) 65–160 days) and eight predominantly white (DIM 72–163 days), with an average body weight of 556 ± 21.23 kg, being all multiparous (between 2 nd and 5 nd parity). The study was conducted in Caturité, Paraiba, Brazil (07°22′45.1″ south latitude, 36°31′47.2″ west longitude and 405 m altitude), during the period of April–May 2014.
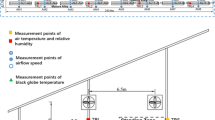
The cows were housed in open areas (50 m long × 25 m wide) with collective drinkers and feeders, and natural shadows of Prosopis juliflora (Fig. 1). The feed consisted of tifton grass hay, corn silage, foraging palm, soybean meal, corn meal, and wheat meal, supplied as complete diet at 0500, 1200, and 1700 h. The mineral and vitamin supplementation was offered ad libitum.

Scheme of the area where the animals were kept during data collection and distance of measurement of coat surface temperature and respiratory rate
Experimental design
Animals were distributed in a completely randomized design, in a factorial arrangement 2 × 2 (coat color × milk yield levels). According to the predominance of the coat color, the animals were divided into two groups: predominantly black (>70 % black coat color) and predominantly white (>70 % white coat color). The proportion of black and white spots on a body surface area was determined directly by visual assessment on both sides of each animal. The tail, head, and limbs were not taken in consideration for the color assessment (Maia et al. 2003). Two months before starting and during experimental period, dairy control were performed at each 2 weeks. By means of dairy controls, cows were divided in two groups according to milk yield, which for group 1, yield levels between 18 and 26 kg milk/day (average of 20 ± 2.55 kg/day) and, for group 2, animals yielding between 10 and 16 kg milk/day (average of 14 ± 1.75 kg/day). The treatment groups were balanced according to DIM and coat color, which in the group 1 were assigned four predominantly white cows and group 2 four predominantly white cows. For predominantly black cows, ten animals were assigned to the group 1 and eight cows were assigned to the group 2.
Environmental data collection
The environmental variables were collected daily from 0500 to 1700 h during the trial period, at intervals of 2 h. Air temperature (TA), relative air humidity (RH), black globe temperature (BGT), and wind speed (Ws) were measured in the holding pen and at two different points in the confinement area: in the shade and in full sun (Fig. 1). TA and RH and WS and BGT were obtained using a set of termohigrometers (model 766401000 Incoterm®; accuracy ±0.35 °C temperature and ±0.25 % relative humidity), anemometer (model LM-8000 Lutron®; accuracy ± 0.1 m/s) and black globe thermometers (a standard 0.15 m diameter hollow copper painted matt black was used), respectively, at the height of 1.20 m from the ground. From the climatic, data was calculated the black globe temperature and humidity index (BGHI) according to Buffington et al. (1981):
where BGT is the black globe temperature, (°C), and DPT is the dew point temperature (°C).
Mean radiant temperature (TMR; °C) was calculated by the equation proposed by Vilella (1983):
where BGT is the black globe temperature (°C); WS is the wind speed (m/s); and TA is the air temperature (°C).
Physiological variables collection
Six days before the beginning of the experimental period, the animals were accustomed to the physiological variables collection routine. Observers walked among the animals simulating the data collections in order to reduce the flight reaction and thus decrease the distance of measurement of the variables. The habituation period has been completed when it was managed to measure the physiological variables at a distance of 3 m from the animal (Fig. 1).
The observations were made 2 or 3 days per week, totaling 10 days of observations per animal. Respiratory rate (F R; breaths/min) and coat surface temperature (Ts; °C) were measured at the times of 0700, 0900, 1100, 1300, and 1500 h. The F R was measured by the number of breaths obtained visually by counting the movements in the animal’s flank, with the aid of a digital timer for 20 s. Subsequently, the value was multiplied by three to obtain the number of breaths per minute. The T S was measured using an infrared thermometer (model 568 Fluke®; accuracy ±1.0 °C), positioned perpendicularly at a distance of approximately 3 m from the animal (Fig. 1), taking average of scanning, following of the scapula to the femur, in the position of 20 cm below the dorsal line of the animal, with automatic calibration and considering the default emissivity of 0.98 for biological tissues (Steketee 1973). At the hottest times of the day (1100, 1300, and 1500 h), physiological variables (F R and T S) were taken when the cows were in the shade, while at 0700 and 0900 h, measurements were taken in the sun.
Prediction equations
Heat transfer through the respiratory tract
Obtaining the physiological data, it was estimated the latent heat transfer (W/m2) through respiratory tract and coat surface, using the equations described below. The rate of heat loss by evaporation from the respiratory tract was estimated according to Silva et al. (2012):
where E R is the heat flow through the respiratory tract; λ is the latent heat of water vaporization (J/g); Ψ EXP is the absolute humidity of the expired air (g/m3); Ψ ATM is the absolute humidity of the atmosphere (g/m3); and r VR is the water vapor resistance to heat loss through the respiratory tract (s/m).
The latent heat of water vaporization can be described by a regression function based on ambient air temperature (TA; °C), according to Maia et al. (2005a):
The absolute humidity of the expired air (Ψ EXP) can be obtained based on the equation described by Silva et al. (2012):
where P VEXP is the saturation pressure of the expired air. To estimate the expired air temperature (T exp), it was used the regression equation described by Maia et al. (2005b) as a function of air temperature (TA):
The absolute humidity of the atmospheric air (Ψ ATM) can be obtained based on the equation described by Silva et al. (2012):
where P VATM is the vapor pressure of the atmospheric air (kPa) at ambient air temperature (Maia et al. 2005b). The water vapor resistance to heat loss through the respiratory tract (r VR) was obtained according to Silva et al. (2012):
where F R is the respiratory rate (breaths/min).
Cutaneous evaporation
To estimate the heat loss by cutaneous evaporation (Ec), it was used the equation developed by Silva and Maia (2011) depending on the animal’s coat surface temperature (Ts):
Statistical analyses
All data were analyzed using SAS System, Version 9.3 (SAS Inst. Inc., Cary, NC). Exploratory analyses were conducted to characterize the data distribution. The normality distribution was checked by skewness and kurtosis coefficients. Data were subjected to analysis of variance by the procedure “general linear model” and means compared by the f test at 5 % confidence level according to the model:
where Y ijlk—variable in analysis; μ—overall mean; H i—effect of time of the day; N k—effect of the yield levels; C l—coat color effect; (HN)ik—interaction between time of the day and level of yield; (HC)il—effect of the interaction between time of the day and coat color; and e ikl—experimental error. Regression models were tested for respiratory and cutaneous evaporation based on respiratory rate and coat surface temperature, respectively.
Results
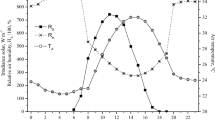
The air temperature ranged from 21 to 30.2 °C, the relative humidity from 87.7 to 52.6 %, and the BGHI ranged from 69.9 to 82.9 in the shade. Notwithstanding, the amplitude of these variables was greater when recorded in full sun, which temperature and relative humidity reached values ranging from 21.1 to 32.3 °C and 92.5 to 44.1 %, respectively, and from 69.8 to 90.2 for the BGHI; the most critical values recorded at 1300 h (Fig. 2).

Climate variables air temperature, relative humidity and black globe temperature and humidity index (mean ± SE) in the shade and in the sun, at coordinates 7°22′45,1″ S and 36°31′47,2″ W in the Brazilian semiarid
There was variation (P < 0.01) for respiratory rate depending on the time of day. For this trait, there was no interaction (P > 0.05) between time of day and coat color categories. Similarly, in both experimental group (Milk yield 20 ± 2.55 vs. 14 ± 1.75 kg) respiratory rate was similar (P > 0.05) in all time of day. The highest values were recorded at 1100, 1300, and 1500 h, corresponding to 66.85 ± 10.20, 66.98 ± 7.80, and 65.65 ± 6.50 breaths/min, respectively. The lowest frequency (54.05 ± 5.09 breaths/min) was observed at 0700 h, followed by 61.30 ± 4.05 breaths/min at 0900 h (P < 0.01; Fig. 3). To estimate the heat loss through the respiratory tract, the model of better adjustment was the quadratic (R 2 = 0.73; Er = 0.5966 + 0.815 F R − 0.0059 F R 2). Thus, the amount of heat transferred via respiration ranged from 19.21 to 29.42 W/m2 (Fig. 4). The respiratory rate and respiratory evaporation showed no variation depending on the milk yield level (P > 0.05; Fig. 5a), as well as there was no variation regarding coat color predominance (P > 0.05; Fig. 5b).

Respiratory rate (mean ± SE) of dairy cows in the period from 0700 to 1500 h

Heat loss by respiratory evaporation according to the respiratory rate of dairy cows

Respiratory rate and evaporation (Mean ± SE) depending on milk yield level (a) and coat color predominance (b) of dairy cows in semiarid ambient
A quadratic function was fitted for cutaneous evaporation as a function of coat surface temperature (R 2 = 0.90; E c = 7191–465.42Ts + 7.5309Ts2; P < 0.01; Fig. 6). There was a variation from 31.6 to 38.8 °C for coat surface temperature, which these values reflected a range of 55.52 to 566.83 W/m2 for heat transfer via cutaneous evaporation. The higher values for heat transfer via surface evaporation (224.57 vs. 275.29 W/m2; Fig. 7b) were observed in times with higher levels of mean radiant temperature in the shade (1100 and 1300 h; 49.6 and 51.0 °C, respectively; Fig. 7a).

Heat loss by cutaneous evaporation of dairy cows in relation to coat surface temperature

Mean radiant temperature (a) and heat loss by cutaneous evaporation (b) (mean ± SD) of dairy cows in semiarid ambient according to times of the day
Coat surface temperature depending on coat color, where predominantly black cows had a higher value compared with white cows (35.49 ± 0.07 °C vs. 35.41 ± 0.08 °C; P < 0.01; Table 1). Consequently, heat loss by cutaneous evaporation followed the same trend; predominantly, black cows lost more heat via cutaneous surface than white cows (186.32 ± 4.09 W/m2 vs. 158.24 ± 6.04 W/m2; P < 0.01; Table 1). There was no variation (P > 0.05) in the coat surface temperature with respect to the milk yield level of cows. Nonetheless, there was effect on cutaneous evaporation regarding the level of yield; thus, animals with an average yield of 20 ± 2.55 kg/day dissipated higher amount of thermal energy in relation to the cows that yielded an average of 14 ± 1.75 kg/day (185.27 ± 5.55 W/m2 vs. 169.40 ± 4.86 W/m2, respectively; P < 0.02; Table 1).
Regarding the percentage distribution of latent heat loss, it was observed that at 0700 h, approximately 80 % of the latent heat loss occurred through the cutaneous evaporation, and the remainder was given through the respiratory tract, while at times with higher air temperatures (1100 to 1300 h) the cutaneous surface heat dissipation reached 90 %. Throughout the day, overall mean for dissipation of thermal energy through the cutaneous surface accounted 87.9 % of total latent heat loss, and the remainder (12.1 %) was through the respiratory tract (Fig. 8).

Participation of the heat loss by cutaneous evaporation and respiratory evaporation of dairy cows in semiarid ambient
Discussion
The use of predictive functions to evaluate the animal heat balance based on simple physiological measures such as coat surface temperature and respiratory rate can be a useful tool in the absence of more sophisticated methods such as masks and respirometric chambers and ventilated capsules. According to Maia et al. (2005b), the use of these mathematical functions should be done taking into account the type of animal and the environment in which these functions have been developed. In the present study, it was estimated from the measurements of respiratory rate and coat surface temperature the heat loss through the cutaneous surface and respiratory tract throughout the day, respectively, for crossbred cows (7/8 Holstein-Gir) raised in confined condition in tropical environment.
According to our results, the increase in the respiratory rate of the animals happened gradually, stabilizing in the afternoon (1100 to 1500 h), times when the air temperature, and the BGHI reached upper critical values of 30.2 and 87.7 °C (shade) and 32.3 and 92.5 °C (full sun), respectively. At times with higher ambient temperature (1100 to 1500 h), the maximum value for respiratory rate was approximately 69 breaths/min. This result may not have reached higher values due to the physical conditions of the environment, since most of the animals at times of high temperatures and intense solar radiation were under shaded areas, provided by P. juliflora.
The increase in the respiratory rate is usually associated with heat exposure, a fact that promotes change in the respiratory behavior of cattle (Gaughan et al. 2000). In studies of crossbred Holstein x Zebu cows, Ferreira et al. (2006) found that the respiratory rate, at morning, was within the physiological limits (17.50 breaths/min), notwithstanding, in the afternoon, this frequency increased by about seven times (134.02 breaths/min), which this situation was triggered by changes in the thermal environment throughout the day, and our results followed the same trend throughout the day. Accordingly, Gaughan et al. (2000) points out that the respiratory rate has been widely used as a heat stress indicator in cattle.
The effect of ambient air temperature on the respiratory rate is not constant and is subject to a number of factors that influence the process (Gaughan et al. 2000). Thus, the effect of air temperature on the respiratory rate is influenced by age, sex, genotype (Azevedo et al. 2005), performance level, nutrition, feeding time, body condition of animals, as well as previous exposure to hot conditions (Gaughan et al. 2000), type of confinement, imposed cooling strategies (Almeida et al. 2011), and other environmental factors. Nonetheless, in this study, it was not observed changes in the respiratory rate regarding the milk yield level, as well as in relation to the prevalence in the animal coat color. Possibly the closeness in the milk yield average (group 1 = 14 ± 1.75 kg; and group 2 = 20 ± 2.55 kg) contributed so that no variations were found in the number of breaths. Regarding the color of the fur, our findings corroborate Pocay et al. (2001), who found no difference in the respiratory rate of predominantly white (93.45 breaths/min) and black (93.88 breaths/min) dairy cows.
In relation to the heat loss by evaporation in animals, breathing is one of the primary mechanisms that utilize the latent heat of vaporization of water to increase the thermal energy dissipation (Rodrigues et al. 2010). It was observed in our study a quadratic response to respiratory evaporation depending on the respiratory movements, verifying that, when the animal reached a rate of 72 breaths/min occurred maximum evaporation through the respiratory tract (29.42 W/m2). Notwithstanding, from this point, even with increase in the respiratory rate, the heat loss by evaporation was not increased. This can be explained by the decrease in the tidal volume as the respiratory rate rises up. According to Maia et al. (2005b), the mass flow through the respiratory tract is also a function of the tidal air volume. Thus, the heat loss through respiratory movement is decreased. In this context, volume of inspired air, associated with body temperature and air humidity, are the most important factors that affect evaporation through the respiratory tract (Silva et al. 2002; Starling et al. 2002).
In our findings, it was observed an average value of about 26.3 W/m2 for heat loss via respiratory evaporation, under average air temperature of 27.3 °C; this corresponded in percentage terms to about 12.1 % of the total latent heat loss of Holstein cows, in open conditions in the semiarid region. In similar semiarid conditions, Silva et al. (2012) found that the evaporation via respiratory tract accounted for 27.3 % of the total latent heat loss, under air temperature conditions of 33.1 °C.
The surface temperature depends mainly on the environmental conditions of humidity and temperature of air and wind, and on the physiological conditions such as vascularization and evaporation through sweat (Ferreira et al. 2006). According to our findings, it was observed that the heat transfer considerably increased due to the increase in coat temperature. This increase in the heat dissipation via cutaneous surface occurred from 0700 to 1300 h and ranged from 117.89 to 275.49 W/m2, respectively, along with the increase, in parallel of the mean radiant temperature (TMR), which reached maximum value (51 °C) at 1300 h. However, the prediction model used was able to estimate the heat loss by cutaneous evaporation when the interval for the coat surface temperature was from 28 to 39 °C. Values outside this range could not be used as indirect estimators and therefore were removed from the database. So, given the limitations imposed by simple regression models, we suggested that more reliable models are developed, as in semiarid conditions of Brazil, in most of the year, high air temperatures and intense solar radiation are common; easily, the temperature of the animal surface exceeds 39 °C (Silva et al. 2014).
In a study with Holstein cows, Hillman et al. (2001) found a skin evaporation rate of about 240 W/m2 in the shade, but when the animals were exposed to direct solar radiation, that rate increased to 500 W/m2, depending on the surface temperature elevation. Maia et al. (2005a) have reported a close relationship between cutaneous evaporation and coat surface temperature, suggesting that it is the physiological activation mechanism for sweating.
In studies with Holstein cows, Maia et al. (2005a) found that the heat loss by skin evaporation accounted for 20–30 % of the total heat loss when the air temperature remained between 10 and 20 °C, the remainder being lost as sensible heat. Under high ambient temperatures (36 °C) and elevated TMR (160.8 °C), animals start gaining heat by sensible ways; then, these temperatures are close to or higher than the body temperature, as is frequently the case in semiarid regions in the intertropical zone. Under this condition, the cutaneous evaporation becomes the primary way of heat loss, accounting for approximately 85 % (350 W/m2) of the total heat loss, while the rest is lost by respiratory evaporation. According to our findings, the loss of heat through the cutaneous surface accounted for approximately 87 % (190.7 W/m2) of the total loss of latent heat of crossbred Holstein cows.
In this study, it was observed that black coat cows showed higher surface temperature (35.49 ± 0.08 °C) than predominantly white cows (35.05 ± 0.08 °C), where this difference in fur temperature can be attributed to the characteristics of absorbance of the coat surface. In studies with Holstein cows, Silva et al. (2003), using a spectro-radiometer with wavelength between 300 and 850 nm, determined absorptance values in the fur of Holstein cows at the level of 0.902 (93 %) for black and 0.518 (35 %) for white. Similar results were found by Façanha et al. (2010), which studying coat characteristics of Holstein cows in the semiarid region, found that the surface temperature of white coat cows was lower (34.05 ± 0.073 °C) than that presented by black coat cows (37.77 ± 0.073 °C). Pocay et al. (2001) observed of 44.59 and 38.18 °C for surface temperature of the black and white coats, respectively. According to our results, heat dissipation by cutaneous evaporation followed the same trend of the surface temperature with respect to the predominance in coat color. Thus, it was observed that cows with black color dissipated greater amount of thermal energy (186.32 ± 4.09 W/m2) than white coat cows (158.24 ± 6.04 W/m2), taking into account the direct relationship between the rate of skin sweating and the coat surface temperature (Hillman et al. 2001; Maia et al. 2005a; Silva and Maia 2011).
The present study demonstrated that the cows that yielded an average of 20 ± 2.55 kg/day eliminated greater amount of heat when compared to the cows that yielded 14 ± 1.75 kg/day, which the values being represented by 185.27 ± 5.55 and 169.40 ± 4.86 W/m2, respectively. Thus, individually, high yielding cows are more vulnerable to heat stress than cows of lower milk yield. This is because milk yield leads to increased production of metabolic heat due to the higher nutrient contribution for synthesis of milk components (Kadzere et al. 2002).
Conclusions
The predictive models based on respiratory rate and coat surface temperature could be used to estimate the latent heat loss in dairy cows kept confined in tropical ambient conditions. Nonetheless, some simple regression models have limitations on their use.
Latent heat dissipation through respiratory tract and cutaneous surface correspond, on average, to 12.1 and 87.9 % of dairy cows bred in semiarid tropical environment.
References
Almeida GL, Pandorfi H, Guiselini C, Henrique HM, Almeida GA (2011) Uso do sistema de resfriamento adiabático evaporativo no conforto térmico de vacas da raça girolando. R Bras Eng Agríc Ambiental 15:754–760
Azevedo M, Pires MDFA, Saturnino HM, Lana AMQ, Sampaio IBM, Monteiro JBN, Morato LE (2005) Estimativa de níveis críticos superiores do índice de temperatura e umidade Para vacas leiteiras 1/2, 3/4 e 7/8 Holandês-zebu em lactação. Rev Bras Zootec 34:2000–2008
Buffington DE, Collazo-Arocho A, Canton GH, Pitt D, Thatcher WW, Collier RJ (1981) Black globe-humidity index (BGHI) as comfort equation for dairy cows. Trans ASAE, Mich 24:711–714
Façanha DAE, Silva RD, Maia ASC, Guilhermino MM, Vasconcelos AD (2010) Variação anual de características morfológicas e da temperatura de superfície do pelame de vacas da raça Holandesa em ambiente semiárido. Rev Bras Zootec 39:837–844
Ferreira F, Pire MF, Martinez ML, Coelho SG, Carvalho AU, Ferreira PM, Facury Filho EJ, Campos WE (2006) Parâmetros fisiológicos de bovinos cruzados submetidos ao estresse calórico. Arq Bras Med Vet Zootec 58:732–738
Finch VA (1985) Comparison of non-evaporative heat transfer in different cattle breeds. Aust J Agric Res 36:497–508. doi:10.1071/AR9850497
Gaughan JB, Holt SM, Hahn GL, Mader TL, Eigenberg R (2000) Respiration rate—is it a good measure of heat stress in cattle? Asian-Aus J Anim Sci 13:329–332
Hillman PE, Lee CN, Parkhurst A (2001) Impact of hair color on thermoregulation of dairy cows to direct sunlight. The Annual International Meeting of the ASAE 94:14–20
Hofmeyr HS, Guidry AJ, Waltz FA (1969) Effects of temperature and wool length on surface and respiratory evaporative losses of sheep. J Appl Physiol 26:517–523
Kadzere CT, Murphy MR, Silanikove N, Maltz E (2002) Heat stress in lactating dairy cows: a review. Livest Prod Sci 77:59–91. doi:10.1016/S0301-6226(01)00330-X
Ligeiro EC, Maia ASC, Silva RG, Loureiro CMB (2006) Perda de calor por evaporação cutânea associada às características morfológicas do pelame de cabras leiteiras criadas em ambiente tropical. Rev Bras Zootec 35:544–549
Maia ASC, Silva RG, Bertipaglia ECA (2003) Características do pelame de vacas Holandesas em ambiente tropical: um estudo genético e adaptativo. Rev Bras Zootec 32:843–853
Maia ASC, Silva RG, Loureiro CMB (2005a) Sensible and latent heat loss from the body surface of Holstein cows in a tropical environment. Int J Biometeorol 50:17–22
Maia ASC, Silva RG, Loureiro CMB (2005b) Respiratory heat loss Holstein cows in a tropical environment. Int J Biometeorol 49:332–336.
Maia ASC, Silva RG, Loureiro CMB (2008) Latent heat loss of Holstein cows in a tropical environment: a prediction model. Rev Bras Zootec 37:1837–1843
McLean JA (1963) Measurement of cutaneous moisture vaporization from cattle by ventilated capsules. J Physiol v.167, p.417–426. doi: 10.1113/jphysiol.1963.sp007159
Pocay PLB, Pocay VG, Starling JMC, Silva RG (2001) Respostas fisiológicas de vacas Holandesas predominantemente brancas e predominantemente negras sob radiação solar direta. Ars Veterinária 17:155–161
Rodrigues NEB, Zangeronimo MG, Fialho ET (2010) Adaptações fisiológicas de suínos sob estresse térmico. Rev Nutr 7:1197–1211
Silva RG (1999) Estimativa do balanço térmico por radiação em vacas Holandesas expostas ao sol e à sombra em ambiente tropical. Rev Bras Zootec 28:1403–1411
Silva RG, LaScala Junior N, Lima Filho AE, Catharin MC (2002) Respiratory heat loss in the sheep: a comprehensive model. Int J Biometeorol 46:136–140
Silva RG, LaScala Junior N, Tonhati H (2003) Radiative properties of the body surface of cattle and other animals. Transassion of the ASEA 46:913–918.
Silva RG, Maia ASC (2011) Evaporative cooling and cutaneous surface temperature of Holstein cows in tropical conditions. Rev Bras Zootec 40:1143–1147
Silva RG, Maia ASC, Costa LLM, Queiroz JPAF (2012) Latent heat loss of dairy cows in an equatorial semi-arid environment. Int J Biometeorol 56:927–932.
Silva RG, Starling JMC (2003) Evaporação cutânea e respiratória em ovinos sob altas temperaturas ambientais. Rev Bras Zootec 32:1–6
Starling JMC, Silva RG, Muñoz M, Barbosa GSSC, Costa MJRP (2002) Análise de algumas variáveis fisiológicas Para avaliação do grau de adaptação de ovinos submetidos ao estresse por calor. Rev Bras Zootec 31:2070–2077
Steketee J (1973) Spectral emissivity of skin and pericardium. Phys Med Biol 18:686
Vilella EC (1983) Confort térmico - Método de Fanger para su evaluación. NTP-74.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Santos, S.G.C.G.d., Saraiva, E.P., Pimenta Filho, E.C. et al. The use of simple physiological and environmental measures to estimate the latent heat transfer in crossbred Holstein cows. Int J Biometeorol 61, 217–225 (2017). https://doi.org/10.1007/s00484-016-1204-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-016-1204-1