
Abstract
Key message
‘Newhall’ grafted onto xiangcheng rootstock with Glomus versiforme or without displayed different responses, and genes related to photosystem II and alpha-linolenic acid metabolism pathways were involved in the responses.
Abstract
Previous studies have shown that there are significant differences in the physiological responses of ‘Newhall’ (Citrus sinensis) scions grafted onto trifoliate orange (Poncirus trifoliata) to arbuscular mycorrhizal (AM) fungi inoculation under normal and stress conditions. However, little is known about the transcriptomic responses of C. sinensis to AM fungi inoculation. In this study, we investigated the effects of inoculating the AM fungus Glomus versiforme on the biomass, pigment content, magnesium (Mg) content and distribution, net photosynthesis rate, and global transcriptome profile of ‘Newhall’ scions grafted onto xiangcheng (Citrus junos) rootstock. The results showed that AM inoculation significantly increased plant growth, Mg concentration, and photosynthesis, but not pigment contents. More than 68,299,008 transcripts were examined in spring-flush leaves, and 29 genes were identified as being differentially expressed in response to mycorrhizal colonization. The differentially expressed genes encoded proteinase inhibitors, transporters, and products related to chlorophyll and disease resistance. Genes encoding proteins related to chlorophyll and transport were up-regulated by AM inoculation while genes encoding proteinase inhibitors were down-regulated. Gene Ontology and KEGG database analyses revealed that genes related to photosystem II and alpha-linolenic acid metabolism pathways were involved in the response to AM inoculation. Comparative expression profiling revealed that the enhancement of photosynthesis after AM inoculation was due to activation of the light-harvesting complex family of proteins in photosystem II. Our results provide new insights into plant–mycorrhizal fungi interactions and their effects on plant growth.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
‘Newhall’ (Citrus sinensis) navel orange is an important citrus cultivar that is usually grafted onto trifoliate orange (Poncirus trifoliata) or xiangcheng (Citrus junos) rootstocks, the most common rootstocks in the citrus-producing areas of China. However, P. trifoliata and C. junos have short root hairs, or few root hairs, in field conditions. Thus, they rely on arbuscular mycorrhizal (AM) fungi for optimal growth (Wang et al. 2012; Wu and Xia 2006). The mutualistic symbiosis between plant roots and AM fungi is a complex trait resulting in cooperative interactions between the two symbionts. The effectiveness of AM colonization in terms of nutrient acquisition and photosynthesis differs markedly among different AM fungi and host plant genotypes (Ortas and Ustuner 2014; Talaat and Shawky 2014). Our previous study showed that an AM fungus enhanced growth, photosynthesis and the absorption of Mg and Zn in ‘Newhall’ scions grafted onto trifoliate orange rootstock under normal and stress conditions (Chen et al. 2014; Xiao et al. 2014). It is well established that the AM symbiosis improves plant nutrition as a direct effect of nutrients being taken up via the mycorrhizal pathway, and as an indirect effect of morphological and physiological changes to the roots (Cavagnaro 2008). AM fungi may also influence nutrient availability via their effects on the physicochemical properties of the soil, nutrient cycling and microbial communities (Cavagnaro et al. 2015). Thus, we hypothesized that the growth, photosynthesis and Mg absorption were also higher in mycorrhizal ‘Newhall’ scions grafted onto xiangcheng rootstock than in non-mycorrhizal plants, and the responses of grafted C. sinensis to AM colonization can be explained using relevant transcriptomic and physiological studies.
The interactions between plant roots and AM fungi include the bi-directional exchange of signalling molecules and metabolic resources. The formation of the mycorrhizal association in the roots leads to plant-wide modulation of gene expression. To understand the systemic effect of the fungal symbiosis on the host, the global transcriptome profilings on tomato fruits (Zouari et al. 2014) have been investigated using RNA-Seq technologies. However, the knowledge about transcriptional diversity and the mechanisms of citrus–mycorrhizal fungi interactions remains largely unstudied. Here, we evaluated the effects of mycorrhization by Glomus versiforme on ‘Newhall’ scions grafted onto xiangcheng rootstocks by comparing plant growth, photosynthesis, Mg concentration and distribution and RNA-seq data between AM-inoculated plants and non-inoculated (non-AM) plants.
Materials and methods
Plant culture and experimental procedures
‘Newhall’ (C. sinensis) navel orange grafted onto 2-year-old xiangcheng (C. junos) seedlings with a uniform rootstock stem diameter (1.3–1.4 cm) were selected for these experiments. The initial shoots (about 4–5 cm above the grafting zone) of the rootstock were removed. All the materials were virus-free (rootstocks and scions) and were harvested from the Fine Varieties Breeding Center of Citrus in Zigui County, Hubei Province, China.
According to Papadakis et al. (2004), the major part of the root system of the plants were severely pruned to maintain highly similar root systems before the treatments and to stimulate new root development. A small number of the roots may have been infected with native AM fungal species, which should not have differed significantly among these plants because they had so few roots. Subsequently, all the plants were washed with deionized water to remove surface contamination, and then transplanted into pots (one plant per pot) containing 3.0 kg quartz sand and perlite (1:1 v/v) medium that had been sterilized by autoclaving (0.11 MPa, 121 °C, 2 h). Before transplantation, the medium had been washed with 3 % (v/v) hydrochloric acid (HCl) overnight and then rinsed thoroughly with deionized water.
The mycorrhizal inoculum, provided by the Institute of Plant Nutrition and Resources at the Beijing Academy of Agriculture and Forestry Sciences, consisted of spores, soil, hyphae and infected root fragments from a stock culture of Glomus versiforme (No. BGC HUN02B). The inoculation dose was 120 g inoculum (containing ~600 spores) per pot. The inocula were placed 5 cm below roots at the time of transplantation. In the non-AM treatment, the same weight of autoclaved mixture was added to the medium.
The plants were initially irrigated with distilled water every 2 days, until the shoots sprouting from the scions were about 10 cm long. Then, the plants were supplied with ½-strength Hoagland’s No. 2 nutrient solution (half-strength macronutrients and full-strength micronutrients) every 2 days. The modified full-strength nutrient solution contained 4 mmol L−1 Ca(NO3)2·4H2O, 6 mmol L−1 KNO3, 1 mmol L−1 NH4H2PO4, 2 mmol L−1 MgSO4·7H2O, 46 μmol L−1 H3BO3, 9 μmol L−1 MnCl2·4H2O, 0.3 μmol L−1 CuSO4·5H2O, 0.1 μmol L−1 H2MoO4 and 50 μmol L−1 Fe-EDTA.
Each scion was grown in a greenhouse for 6 months under natural daylight from April to September. The average day/night temperature was 25/20 °C and the average air relative humidity was 80 %. The site was located at 31°20′N latitude, 119°21′E longitude, at 10 m above sea level.
Analyses of mycorrhizal colonization, plant biomass and magnesium
At harvest time, plant height was measured with a measuring tape. A portion of fresh roots was carefully washed and cut into 1-cm pieces to fix in formalin–acetic acid–alcohol solution. These root pieces were cleaned with 10 % (w/v) KOH and stained with 0.05 % (w/v) trypan blue in lactophenol (Phillips and Hayman 1970), and microscopically detected for root colonization. The mycorrhizal colonization was counted by the following formula: mycorrhizal colonization (%) = 100 × root length infected/root length detected. Mycorrhizal dependency was defined as the ratio of the dry weight of the AM-inoculated plants to that of the non-AM-inoculated plants (Graham and Syvertsen 1985).
For each sample, the leaves, scion stems, rootstock stems, and roots were separated. For the leaves, basal leaves (spring-flush) and upper leaves (summer and autumn-flush) were collected based on different phases of shoot growth. The plant materials were dried at 75 °C for 48 h and the dry weights were measured. Then, samples were ground to a fine powder and wet-digested in HNO3–HClO4 (4:1 v/v) before Mg analysis by inductively coupled plasma optical emission spectrometry (ICP-OES, Perkin Elmer Optimal 2100 DV, Perkin Elmer, USA).
Pigment and photosynthesis analyses
Chlorophylls and carotenoids were extracted from spring-flush leaves and quantified as described by Wang (2006). The net photosynthetic rate (P n) of spring-flush leaves was determined using a CI-340 portable photosynthesis system (CID Bio-Science, Inc., Camas, WA, USA) at an ambient CO2 concentration under a controlled light intensity of 1000 μmol m−2 s−1 between 10:00 and 11:00 on a clear day.
Experimental design and statistical analysis
The experiment was made up of G. versiforme-inoculated treatments (AMF) and non-AM-inoculated treatments (non-AMF). Three replications (two plants in each replicate) were designated for each treatment, making a total of 12 pots.
The data were subjected to analysis of variance (ANOVA) using Statistical Product and Service Solutions software (SPSS Institute Inc. Chicago, IL, USA) and differences were compared by Duncan’s test with a significance level of p < 0.05.
RNA-seq library preparation and sequencing
To examine the transcriptome of mycorrhizal ‘Newhall’ plants, we collected spring-flush leaves from AM plants and non-AM plants at harvest time. The leaves from groups of three plants per treatment were pooled and frozen in liquid nitrogen and stored at −80 °C for RNA extraction. Total RNA was extracted from the above-cited leaf samples using Trizol reagent. Then, RNA degradation and contamination was monitored by electrophoresis on 1 % agarose gels. For each sample, 3 μg RNA was used as input material to prepare RNA samples. The enrichment of mRNAs, fragment interruption, addition of adapters, size selection, PCR amplifications and RNA-seq were performed by staff at WuXi AppTec (Shanghai, China). Poly(A) mRNA was isolated using oligo dT beads and then cleaved into short fragments. The RNA-seq libraries were prepared from two samples of AM and non-AM plants, and sequenced on the Illumina HiSeq 2000 platform.
Quality control
Clean data were obtained by removing reads containing adapters and low-quality reads from the raw data. All subsequent analyses were based on clean, high-quality data.
Read mapping to reference genome
The reference genome and gene model annotation files were downloaded directly from the citrus genome website (http://citrus.hzau.edu.cn/orange/). An index of the reference genome was built using Bowtie software. Raw reads were filtered to obtain high-quality reads by removing low-quality reads. The high-quality reads were mapped onto the C. sinensis reference genome using TopHat. Differential expression was estimated and tested with R software. Gene expression was quantified in terms of RPKM values. Transcripts with p < 0.05 were considered to be significantly differentially expressed between AM and non-AM plants.
Quantification of gene expression levels
The R package cummeRbund was used to count the number of reads mapped to each gene. Then, the reads per kilobase of exon model per million mapped reads (RPKM) value for each gene was calculated based on the length of the gene and the number of reads mapped to it.
Differential expression analysis
Before the differential gene expression analysis, for each sequenced library, the read counts were adjusted using the R program package through one scaling normalized factor. A corrected p value of 0.05 was set as the threshold for significant differential expression.
GO and KEGG enrichment analysis of differentially expressed genes
We performed a gene ontology (GO) functional analysis to identify the biological processes that were most strongly represented by the differentially expressed genes (DEGs). We also conducted a GO enrichment analysis for DEGs. GO enrichment analyses of mycorrhizal-responsive genes were implemented with InterPro software. GO terms with a corrected p value <0.05 were considered to be significantly enriched.
A KEGG pathway (http://www.genome.jp/kegg/pathway.html) enrichment analysis was conducted to identify significantly enriched metabolic or signal transduction pathways in DEGs compared with the whole genome background.
Results
Plant growth and photosynthesis
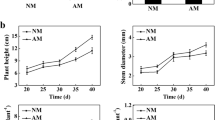
The absence of mycorrhizae from roots was confirmed for the non-AM ‘Newhall’ plants. Compared with non-AM plants, the AM plants showed a 15.95 % increase in plant height, a 20.87 % increase in leaf number, and a 22.91 % increase in dry weight per plant (Table 1).
There were no significant differences between non-AM and AM plants in leaf chlorophyll a, chlorophyll b, chlorophyll a + b, carotenoid concentrations. However, the P n was 48.56 % higher in AM plants than in non-AM plants (Table 2).
Magnesium concentrations in different plant parts
The AM plants showed significantly increased Mg concentrations in various parts of the plant. The Mg concentrations in the upper leaves, basal leaves, scion stems, rootstock stems and roots were 15.93, 10.34, 23.56, 26.09 and 18.25 % higher, respectively, in AM plants than in non-AM plants. Regardless of the presence of the AM fungus, the highest Mg concentrations were in the roots and the lowest were in the rootstock stems (Fig. 1).

Mg concentrations of upper leaves, basal leaves, scion stems, rootstock stems and roots of ‘Newhall’ (C. sinensis) scions grafted onto xiangcheng (C. junos) rootstock with an arbuscular mycorrhizal fungi (AMF) or without (Non-AMF). Values represent the mean ± SE of three replicates (n = 3), samples from two plants were collected for each replicate. Bars with asterisk indicate significant differences at p < 0.05
Read number, transcriptome coverage and total expressed genes
To identify DEGs between AM and non-AM plants, we filtered the sequences, estimated the transcriptome coverage and then identified the expressed genes. The raw reads obtained from the Illumina HIseq were filtered and checked for sequence contaminants with the fastQC application. Contaminant-free, filtered reads (120–130 million for each sample; Table 3) were mapped with Bowtie/TopHat to the navel orange genome sequence that was obtained from the website. The raw read counts were obtained using HTSeq software. A RPKM cut-off value of 0.1 was set to detect the expression of a particular locus.
To obtain an overview of the transcriptional information for navel orange leaves and prepare the reference database for the RNA-Seq analysis, a normalized cDNA pool was constructed from AM and non-AM plants using the Illumina HiSeq 2000 platform. First, we obtained a total of about 124.7 million raw reads with an average length of 125 nt (Table 3). After data filtering and quality checks, more than 124.5 million clean reads (99.78 % of the raw reads) were generated with a Q30 (base quality more than 30) of 89.0 %. The total length of the reads was more than 31.2 Gb. Paired-end sequencing was performed on the two samples using the Illumina Hiseq 2000 platform, generating 139,684,647 (non-AM plants) and 124,790,090 (AM plants) 125-bp reads. We pooled the short reads and aligned them against the citrus reference genome using TopHat.
With a false discovery rate (FDR) <0.001, a total of 29 genes were identified as DEGs; 15 DEGs had reduced expression levels in AM plants, and 14 DEGs had increased expression levels in AM plants (Fig. 2). The expression levels of most genes were unaffected by AM inoculation, suggesting that AM colonization targets only a limited number of genes.

Differentially expressed genes (DEGs) between non-AM plants and AM plants: the up- and down-regulated represent the genes that had an increased and decreased expression levels in navel orange plants
Differentially expressed genes and overrepresented categories
To identify DEGs, we used CummeRbund in the R package to evaluate the significance of differences in expression and to control for the FDR. Pearson’s correlation coefficients among biological replicates were subjected to the same treatment and were all greater than 0.95. The FDR threshold was set to 0.1 and gene dispersion values were calculated by fitting a curve using the DESeq “fit-only” mode. Using these criteria, we identified 29 DEGs (Table 4), including 14 up-regulated genes and 15 down-regulated genes in AM plants, compared with non-AM plants. We also determined the degree of differential expression by comparing the log-transformed fold-change in expression levels between control and AM plants. Among the DEGs, the three most highly expressed genes in AM plants were those encoding a chlorophyll a/b-binding protein, a pentatricopeptide repeat-containing protein, and the chloroplastic pumilio homolog 6. These gene products are related to important processes in the chloroplast.
With the CummeRbund package in the statistics environment R, we set the adjusted p value to 0.05 as the threshold to select DEGs between non-AM and AM plants (Supplementary Fig. S1). To estimate the gene expression levels from the sequencing data, the fragments per kilobase of exon model per million mapped fragments (FPKM) values were applied to normalize and evaluate each unigene. Statistics of the distribution of the FPKM values showed that the expression level of most unigenes was between 0 and 100.
Gene ontology annotation of differentially expressed genes
We conducted a gene ontology (GO) enrichment analysis to identify the major gene groups affected by mycorrhizal inoculation. As in all RNA-Seq experiments, the variable length of transcripts may produce biases in the data. The functional classifications were defined using GO terms, which provide broad functional classifications for genes and gene products based on their corresponding biological processes, cellular components and molecular functions (Fig. 3). The largest category was molecular function, and ‘endopeptidase inhibitor activity’ was the largest group in this category, followed by ‘protein binding’ and ‘RNA binding’. Among the ten functional groups in the biological process category, GO terms were predominantly associated with ‘protein phosphorylation’, ‘signal transduction’, and ‘defence response’. The groups of GO terms suggested that a large part of the navel orange transcriptome is devoted to the control of the main metabolic processes (i.e. photosynthesis) and to ion transport. For example, genes involved in photosynthesis, transport process, and catalytic activity accounted for 21.4, 28.57 and 50 %, respectively, of the core set of regulated genes.

Gene ontology (GO) enrichment in differentially expressed genes (DEGs). The 32 enriched subcategories are shown. The y-axis indicates the different subcategories, and the x-axis indicates the number of DEGs in a subcategory
Pathway definitions were obtained from the KEGG database (Supplementary Fig. S2). Genes involved in photosystem II and the alpha-linolenic acid metabolism pathway showed marked changes in expression between AM and non-AM orange plants. The very high proportions of trienoic fatty acids in the chloroplast membranes of all higher plants suggest that these lipids might be essential for photosynthesis. The chlorophyll-binding subunits of photosystems I and II are internal antenna light-harvesting proteins involved in oxygenic photosynthesis. These antenna proteins in the light-harvesting chlorophyll protein complexes in green plants act as peripheral antenna systems, enabling more efficient absorption of light energy. These annotations provide a valuable resource for investigating specific processes, functions and pathways that are activated or repressed in navel orange inoculated with an AM fungus.
Exploration of gene expression
We conducted a hierarchical clustering analysis of the DEGs to observe the overall gene expression patterns. We generated a heatmap of RPKM-normalized transcript counts to illustrate the DEGs between AM plants and non-AM plants. As shown in Fig. 4, 23 DEGs were divided into five groups: chlorophyll, disease resistance protein, proteinase inhibitor, transporter and unknown. The genes in the chlorophyll, disease resistance protein and transporter groups were up-regulated in AM plants. For example, genes encoding the chlorophyll a/b-binding protein, a TIR-NBS-LRR protein, and a zinc transporter were up-regulated in AM plants. On the other hand, some genes in the proteinase inhibitor group were down-regulated in AM plants, for example, genes encoding a Kunitz-type protease inhibitor and a latex serine protease inhibitor.

Clustering and heatmap of expression values for differentially expressed genes (DEGs). Light and dark colors indicate higher and lower expression values, respectively. The heatmap was generated from CummeRbund normalized counts and further processed with custom scripts based on heatmap function
Discussion
Differences in growth between AM and non-AM citrus plants
Previous studies have shown that AM fungal colonization could exert significant influence on the root system architecture, which were linked to the improved uptake of nutrients from soil, and then affected plant growth (Padilla and Encina 2005; Yao et al. 2009). In this study, AM inoculation increased the P n in the leaves and the dry weight per plant of ‘Newhall’ navel orange plants. This may be related to changes in endogenous hormone levels, and increased uptake and translocation of water and nutrients by AM plants (Goicoechea et al. 1997).
It is widely accepted that the AM association increases the biomass production of the plant. Similarly, the growth of prairie grasses was strongly correlated with AM colonization (Wilson et al. 2012). In acacia, root and shoot weights were significantly higher in AM plants than in non-AM plants (Giri et al. 2003). However, there were no significant differences in the dry weights of organs of Trifolium repens between AM and non-AM plants (Wright et al. 1998). In our study, we found that the biomass of C. sinensis plants was markedly increased in AM plants, both for above-ground and below-ground organs. These results indicated that this specific AM fungus positively affected the growth of grafted navel orange plants. However, the effects of AM fungi on growth differ among plant species. The production and distribution of biomass can involve significant energy inputs, consuming photosynthetic products and nutrients. In another study, the specific leaf area was greater in AM orange plants than in non-AM orange plants, leading to an increased photosynthetic rate in the AM plants (Ruotsalainen and Kytöviita 2004; Wright et al. 1998). Also, the acquisition of nutrients (e.g. phosphorus and nitrogen) was greater in AM plants than in non-AM plants, and this may have contributed to the greater biomass of AM plants (Jha et al. 2012; Liu et al. 2014).
Genes related to photosynthesis
Many studies have shown that AM fungi positively affect photosynthesis, consistent with our findings. Previous studies have reported that chlorophyll a, chlorophyll b and total chlorophyll concentrations were significantly higher in AM plants than in non-AM plants (Wu et al. 2015). An AM fungus may stimulate plant growth through increasing the total chlorophyll concentration. However, we observed that the total photosynthetic pigment (chlorophyll and carotenoid) concentrations were not significantly different between AM and non-AM navel orange plants. Therefore, the observed increase in P n did not result from increased chlorophyll concentrations in this case. It was shown that AM plants exhibited increased efficiency for photosynthetic energy conservation and increased photosynthetic performance (Zubek et al. 2009, 2010). Additionally, AM infection led to more intense electron flow and a higher productive photosynthetic activity at several sites of the photosynthetic electron-transport chain. Also, AM fungi were shown to have a stabilizing effect on the donor site of photosystem II and on the electron-transport chain between photosystems I and II (Pinior et al. 2005). In all plants, photosynthesis uses solar energy collected by the light-harvesting complex (LHC) associated with photosystem II. The results of the RNA-Seq analysis revealed that several genes encoding products related to the LHC were up-regulated in AM navel orange plants, in agreement with Cicatelli et al. (2012), who showed that genes encoding the chlorophyll A-B-binding protein/LHC1 type III are up-regulated in AM white poplar plants.
Transport-related genes
The AM fungus establishes a symbiosis with roots and improves the mineral nutrition of the plant. One of the most dramatic effects of colonization by AM fungi on the host plant is the increase in phosphorus uptake. Phosphorus has been shown to play important role in the tolerance of mycorrhizal plants to pathogens and alleviates the deleterious effects of saline soils on plant growth (Giri et al. 2007).
Recent research has identified the networks of genes involved in phosphate and nitrogen acquisition in plants with mycorrhizal symbioses. Analyses of the phylogenetic relationships of transport proteins in different plants revealed that they were non-orthologous (Paszkowski et al. 2002). Some genes encoding Pi- and N-transporters have been cloned from pea (Grønlund et al. 2013), maize (Tian et al. 2013) and Sorghum (Koegel et al. 2013), but few genes involved in Mg transport have been shown to be involved in mycorrhizal function. Burleigh et al. (2003) reported that a Zn transporter in Medicago truncatula was regulated by a mycorrhizal fungus. The association of AM fungi with the roots of navel orange significantly enhanced Mg accumulation in all parts of the plant, especially the roots and leaves. Analyses of the transcriptomic profiles revealed that a set of genes that potentially encode Mg-, Zn- and Co- transporters were up-regulated in AM plants. These molecular data suggest that AM navel orange translocated more Mg into the leaves to increase the photosynthetic rate, consistent with the physiological data.
Defence-related regulation
The successful colonization of plant roots by beneficial microbes depends on the ability of the microbe to manipulate the defence responses of the host. AM inoculation leads to increased disease resistance and higher activities/increased abundance of antioxidant enzymes, stress-responsive proteins, defensive enzymes, chitinase, specific lectins and increased concentrations of some plant hormones (Foo et al. 2014; Zhao et al. 2014; Xiao et al. 2015).
In our study, AM plants showed repressed expression of some genes encoding pathogenesis-related proteins. TIR-NBS-LRR proteins include a nucleotide-binding site (NBS), a leucine-rich repeat (LRR) region and a motif homologous to the cytoplasmic domains of the Drosophila Toll protein and the mammalian interleukin-1 receptor (TIR). These proteins are a type of disease resistance (R) protein. Resistant plant genotypes can prevent pathogen entry via a defence mechanism, which, in the simplest model, is initiated through a direct or indirect interaction between an R gene product and a specific biotrophic pathogen avirulence gene product (Miller et al. 2008). The observation in tomato that defense-related PR-1 protein accumulates around arbuscules has been reported for many other mycorrhizal plants. Gianinazzi-Pearson (1996) suggested that the resistance of arbuscule-containing cells is linked to the low priming of defense genes in these cells. We believe that the expression of defense genes may both affect arbuscule-containing and the stage of AM development. Krajinski et al. (1998) reported that RNA accumulation of the gene psam2, which could belong to a group of genes related to plant defence or pathogenesis, was repressed at late stages of AM development. In the present investigation, we found that the expression levels of the TIR-NBS-LRR disease resistance protein are decreased, which may be beneficial to AM development in navel orange plants.
Concomitantly, many genes encoding protease inhibitors were induced in the AM plants. Protease inhibitors are widely distributed in plants. They function in biochemical defence responses, as storage proteins, and in the regulation of endogenous protease activity. We observed that genes encoding Kunitz-type proteinase inhibitors were activated in the AM navel orange plants. These proteins are a prominent feature of local and systemic defence responses in plants induced by insect feeding or mechanical wounding (Philippe et al. 2009).
In conclusion, our results show that an AM fungus significantly increased the growth, net photosynthetic rate and Mg concentrations in grafted C. sinensis plants, and also affected the distribution of Mg in the plant tissues. Genes encoding proteinase inhibitors, transporters, disease resistance proteins and chlorophyll-binding proteins were differentially expressed between AM and non-AM plants. Our results suggest that AM fungi increase the growth of C. sinensis plants by enhancing photosynthesis and nutrient absorption. The AM plants strongly expressed genes encoding transporters and showed increased Mg concentrations. Also, their increase in photosynthesis was accompanied by increased expression of genes encoding members of the LHC family, which form a part of photosystem II. Nevertheless, the regulation and function of some genes are still poorly understood. Therefore, further research is required to clarify the mechanisms by which AM fungi promote plant growth.
Author contribution statement
Study design, data acquisition and analysis, drafting the manuscript: X. Gao and S. Zhao. Study design, manuscript preparation and editing: Q.-L. Xu. Manuscript revision/view, and final version approval: J.-X. Xiao.
References
Burleigh SH, Kristensen BK, Bechmann IE (2003) A plasma membrane zinc transporter from Medicago truncatula is up-regulated in roots by Zn fertilization, yet down-regulated by arbuscular mycorrhizal colonization. Plant Mol Biol 52:1077–1088. doi:10.1023/A:1025479701246
Cavagnaro TR (2008) The role of arbuscular mycorrhizas in improving plant zinc nutrition under low soil zinc concentrations: a review. Plant Soil 304:315–325. doi:10.1007/s11104-008-9559-7
Cavagnaro TR, Bender SF, Asghari HR, van der Heijden MGA (2015) The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci 20(5):283–290. doi:10.1016/j.tplants.2015.03.004
Chen Y-Y, Hu C-Y, Xiao J-X (2014) Effects of arbuscular mycorrhizal inoculation on the growth, zinc distribution and photosynthesis of two citrus cultivars grown in low-zinc soil. Trees 28:1427–1436. doi:10.1007/s00468-014-1046-6
Cicatelli A, Lingua G, Todeschini V, Biondi S, Torrigiani P, Castiglione S (2012) Arbuscular mycorrhizal fungi modulate the leaf transcriptome of a Populus alba L. clone grown on a zinc and copper-contaminated soil. Environ Exp Bot 75:25–35. doi:10.1016/j.envexpbot.2011.08.012
Foo E, Ferguson BJ, Reid JB (2014) Common and divergent roles of plant hormones in nodulation and arbuscular mycorrhizal symbioses. Plant Signal Behav 9:e29593. doi:10.4161/psb.29593
Gianinazzi-Pearson V (1996) Plant cell responses to arbuscular mycorrhizal fungi: getting to the roots of the symbiosis. Plant Cell 8:1871–1883. doi:10.1105/tpc.8.10.1871
Giri B, Kapoor R, Mukerji K (2003) Influence of arbuscular mycorrhizal fungi and salinity on growth, biomass, and mineral nutrition of Acacia auriculiformis. Biol Fertil Soils 38:170–175. doi:10.1007/s00374-003-0636-z
Giri B, Kapoor R, Mukerji K (2007) Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum may be partly related to elevated K/Na ratios in root and shoot tissues. Microb Ecol 54:753–760. doi:10.1007/s00248-007-9239-9
Goicoechea N, Antolin M, Sánchez-Díaz M (1997) Gas exchange is related to the hormone balance in mycorrhizal or nitrogen-fixing alfalfa subjected to drought. Physiol Plant 100:989–997. doi:10.1111/j.1399-3054.1997.tb00027.x
Graham J, Syvertsen J (1985) Host determinants of mycorrhizal dependency of citrus rootstock seedlings. New Phytol 101:667–676. doi:10.1111/j.1469-8137.1985.tb02872.x
Grønlund M, Albrechtsen M, Johansen IE, Hammer EC, Nielsen TH, Jakobsen I (2013) The interplay between P uptake pathways in mycorrhizal peas: a combined physiological and gene-silencing approach. Physiol Plant 149:234–248. doi:10.1111/ppl.12030
Jha A, Kumar A, Saxena R, Kamalvanshi M, Chakravarty N (2012) Effect of arbuscular mycorrhizal inoculations on seedling growth and biomass productivity of two bamboo species. Indian J Microbiol 52:281–285. doi:10.1007/s12088-011-0213-3
Koegel S, Ait Lahmidi N, Arnould C, Chatagnier O, Walder F, Ineichen K, Boller T, Wipf D, Wiemken A, Courty PE (2013) The family of ammonium transporters (AMT) in Sorghum bicolor: two AMT members are induced locally, but not systemically in roots colonized by arbuscular mycorrhizal fungi. New Phytol 198:853–865. doi:10.1111/nph.12199
Krajinski F, Martin-Laurent F, Gianinazzi S, Gianinazzi-Pearson V, Franken P (1998) Cloning and analysis of psam2, a gene from Pisum sativum L. regulated in symbiotic arbuscular mycorrhiza and pathogenic root-fungus interactions. Physiol Mol Plant Pathol 52:297–307
Liu H, Tan Y, Nell M, Zitter-Eglseer K, Wawscrah C, Kopp B, Wang S, Novak J (2014) Arbuscular mycorrhizal fungal colonization of Glycyrrhiza glabra roots enhances plant biomass, phosphorus uptake and concentration of root secondary metabolites. J Arid Land 6:186–194. doi:10.1007/s40333-013-0208-5
Miller RN, Bertioli DJ, Baurens FC, Santos CM, Alves PC, Martins NF, Togawa RC, Souza MT, Pappas GJ (2008) Analysis of non-TIR NBS-LRR resistance gene analogs in Musa acuminata Colla: isolation, RFLP marker development, and physical mapping. BMC Plant Biol 8:15. doi:10.1186/1471-2229-8-15
Ortas I, Ustuner O (2014) The effects of single species, dual species and indigenous mycorrhiza inoculation on citrus growth and nutrient uptake. Eur J Soil Biol 63:64–69. doi:10.1016/j.ejsobi.2014.05.007
Padilla IMG, Encina CL (2005) Changes in root morphology accompanying mycorrhizal alleviation of phosphorus deficiency in micropropagated Annona cherimola Mill. plants. Sci Hortic 106:360–369. doi:10.1016/j.scienta.2005.05.001
Papadakis IE, Dimassi KN, Bosabalidis AM, Therios IN, Patakas A, Giannakoula A (2004) Effects of B excess on some physiological and anatomical parameters of ‘Navelina’ orange plants grafted on two rootstocks. Environ Exp Bot 51:247–257. doi:10.1016/j.envexpbot.2003.11.004
Paszkowski U, Kroken S, Roux C, Briggs SP (2002) Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proc Natl Acad Sci 99:13324–13329. doi:10.1073/pnas.202474599
Philippe RN, Ralph SG, Külheim C, Jancsik SI, Bohlmann J (2009) Poplar defense against insects: genome analysis, full-length cDNA cloning, and transcriptome and protein analysis of the poplar Kunitz-type protease inhibitor family. New Phytol 184:865–884. doi:10.1111/j.1469-8137.2009.03028.x
Phillips J, Hayman D (1970) Improved procedures for clearing roots and staining parasitic and vesicular–arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br Mycol Soc 55:158–IN118. doi:10.1016/S0007-1536(70)80110-3
Pinior A, Grunewaldt-Stöcker G, von Alten H, Strasser RJ (2005) Mycorrhizal impact on drought stress tolerance of rose plants probed by chlorophyll a fluorescence, proline content and visual scoring. Mycorrhiza 15:596–605. doi:10.1007/s00572-005-0001-1
Ruotsalainen AL, Kytöviita M-M (2004) Mycorrhiza does not alter low temperature impact on Gnaphalium norvegicum. Oecologia 140:226–233. doi:10.1007/s00442-004-1586-3
Talaat NB, Shawky BT (2014) Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity. Environ Exp Bot 98:20–31. doi:10.1016/j.envexpbot.2013.10.005
Tian H, Drijber RA, Li X, Miller DN, Wienhold BJ (2013) Arbuscular mycorrhizal fungi differ in their ability to regulate the expression of phosphate transporters in maize (Zea mays L.). Mycorrhiza 23:507–514. doi:10.1007/s00572-013-0491-1
Wang X (2006) Experimental principle and technique for plant physiology and biochemistry. Higher Education Press, Beijing, pp 118–283
Wang P, Zhang J, Shu B, Xia R (2012) Arbuscular mycorrhizal fungi associated with citrus orchards under different types of soil management, southern China. Plant Soil Environ 58:302–308
Wilson GW, Hickman KR, Williamson MM (2012) Invasive warm-season grasses reduce mycorrhizal root colonization and biomass production of native prairie grasses. Mycorrhiza 22:327–336. doi:10.1007/s00572-011-0407-x
Wright D, Scholes J, Read D (1998) Effects of VA mycorrhizal colonization on photosynthesis and biomass production of Trifolium repens L. Plant Cell Environ 21:209–216. doi:10.1046/j.1365-3040.1998.00280.x
Wu Q-S, Xia R-X (2006) Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J Plant Physiol 163:417–425. doi:10.1016/j.jplph.2005.04.024
Wu Q-S, Lou YG, Li Y (2015) Plant growth and tissue sucrose metabolism in the system of trifoliate orange and arbuscular mycorrhizal fungi. Sci Hortic 181:189–193. doi:10.1016/j.scienta.2014.11.006
Xiao JX, Hu CY, Chen YY, Yang B, Hua J (2014) Effects of low magnesium and an arbuscular mycorrhizal fungus on the growth, magnesium distribution and photosynthesis of two citrus cultivars. Sci Hortic 177:14–20. doi:10.1016/j.scienta.2014.07.016
Xiao JX, Hu CY, Chen YY, Hua J, Yang B (2015) Growth and nutrient content of trifoliate orange seedlings influenced by arbuscular mycorrhizal fungi inoculation in low magnesium soil. J Plant Nutr 38:1516–1529. doi:10.1080/01904167.2014.957400
Yao Q, Wang LR, Zhu HH, Chen JZ (2009) Effects of arbuscular mycorrhizal fungal inoculation on root system architecture of trifoliate orange (Poncirus trifoliata L. Raf.) seedlings. Sci Hortic 121:458–461. doi:10.1016/j.scienta.2009.03.013
Zhao X, Zhang J, Chen C, Yang J, Zhu H, Liu M, Lv F (2014) Deep sequencing-based comparative transcriptional profiles of Cymbidium hybridum roots in response to mycorrhizal and non-mycorrhizal beneficial fungi. BMC Genom 15:747. doi:10.1186/1471-2164-15-747
Zouari I, Salvioli A, Chialva M, Novero M, Miozzi L, Tenore GC, Bagnaresi P, Bonfante P (2014) From root to fruit: RNA-Seq analysis shows that arbuscular mycorrhizal symbiosis may affect tomato fruit metabolism. BMC Genom 15:221. doi:10.1186/1471-2164-15-221
Zubek S, Turnau K, Tsimilli-Michael M, Strasser RJ (2009) Response of endangered plant species to inoculation with arbuscular mycorrhizal fungi and soil bacteria. Mycorrhiza 19:113–123. doi:10.1007/s00572-008-0209-y
Zubek S, Stojakowska A, Anielska T, Turnau K (2010) Arbuscular mycorrhizal fungi alter thymol derivative contents of Inula ensifolia L. Mycorrhiza 20:497–504. doi:10.1007/s00572-010-0306-6
Acknowledgments
This work was supported by the National Natural Science Foundation of China (31372014), Anhui Provincial Natural Science Foundation (1308085MC37) and the program of Collaborative Innovation Center of Recovery and Reconstruction of Degraded Ecosystem in Wanjiang City Belt.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by P. Courty.
X. Gao and S. Zhao contributed equally as first author.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Gao, X., Zhao, S., Xu, QL. et al. Transcriptome responses of grafted Citrus sinensis plants to inoculation with the arbuscular mycorrhizal fungus Glomus versiforme . Trees 30, 1073–1082 (2016). https://doi.org/10.1007/s00468-015-1345-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-015-1345-6