
Abstract
Large-scale deforestation in the tropics, triggered by logging and subsequent agricultural monoculture has a significant adverse impact on biodiversity due to habitat degradation. Here, we measured the diversity of butterfly species in three agricultural landscapes, agroforestry orchards, oil palm, and rubber tree plantations. Butterfly species were counted at 127 sampling points over the course of a year using the point count method. We found that agroforestry orchards supported a greater number of butterfly species (74 species) compared to rubber tree (61 species) and oil palm plantations (54 species) which were dominated by generalist (73%) followed by forest specialists (27%). We found no significant difference of butterfly species composition between agroforestry orchards and rubber tree plantation, with both habitats associated with more butterfly species compared to oil palm plantations. This indicates butterflies were able to persist better in certain agricultural landscapes. GLMMs suggested that tree height, undergrowth coverage and height, and elevation determined butterfly diversity. Butterfly species richness was also influenced by season and landscape-level variables such as proximity to forest, mean NDVI, and habitat. Understanding the factors that contributed to butterfly species richness in an agroecosystem, stakeholders should consider management practices to improve biodiversity conservation such as ground vegetation management and retaining adjacent forest areas to enhance butterfly species richness. Furthermore, our findings suggest that agroforestry system should be considered to enhance biodiversity in agricultural landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The large-scale conversion of tropical forests into agricultural areas raised concerns among conservationists due to its major detrimental impacts on biodiversity (Barnes et al. 2014; Ahrends et al. 2015; Azhar et al. 2015a; Warren-Thomas et al. 2020; Alroy 2017; Giam 2017; Pashkevich et al. 2020; Potapov et al. 2020). Most land-use change and intensification lead to landscape simplification and fragmentation, which result in significant biodiversity losses due to habitat destruction (Ewers et al. 2009), as well as disruption of ecosystem services such as pollination, pest control, and nutrient cycling (Landis 2017; Power 2010). With the loss of ecosystem services, human well-being will be in jeopardy (Diaz et al. 2006). Nevertheless, agroforestry system that includes productive and protected areas can reduce the impact of tropical deforestation by promoting a mosaic of different land-use types (Stanturf et al. 2019; Schwarz et al. 2021).
In Malaysia, about 33.1% of the total land area has been converted for agricultural purposes (MPIC 2015); agriculture sectors account for approximately 23% of total exports and 7.2% of Malaysia's GDP (Rozhan 2015). The land conversion motivated by agricultural intensification in Malaysia began during the British colonial era with the introduction of monoculture rubber tree (Hevea brasilensis) and oil palm (Elaeis guineensis) plantations from Brazil and South Africa, respectively (Athukorala and Loke 2009; Nath and Chaudhuri 2010; Sayer et al. 2012). To date, approximately 5,865,290 ha of land have been planted with oil palm trees and 1,106,861 ha with rubber trees (MPIC 2015). Furthermore, a mix of native fruit trees is planted in agroforestry orchards to provide farmers with additional income (Salma et al. 2006; Abdullah 2011), covering approximately 116,369 ha of land (DOSM 2019). Thus, changes in habitat structure (both at the local and landscape scale) caused by different agricultural practices in each respective farmlands may harbor different composition of associated wildlife (Pogue and Schnell 2001; Azhar et al. 2015b).
Butterflies are well-documented taxa for agricultural biodiversity research and are frequently paired with other taxa such as birds (Posa and Sodhi 2006; Koh 2008; Azhar et al. 2015b; Salek et al. 2018), spiders (Salek et al. 2018), bats (Azhar et al. 2015b), and ants (Lucey and Hill 2012). Besides offering a wide range of ecosystem services, butterflies are good indicators for environmental changes as their diversity is related to other taxa such as birds and spiders (Ekroos et al. 2013; Salek et al. 2018). Butterflies play major roles in the ecosystem as food resources to other animals, pollinators by transferring pollen to flowers, and defoliators on numerous plants (Stokes et al. 1991; Cleary 2004; Karen-Chia 2014).
Previous research has shown that the richness of butterfly species in oil palm agroecosystems is influenced by landscape and local scale-level habitat attributes (Azhar et al. 2015b; Asmah et al. 2017). At the landscape level, mixed-crop agriculture supports more diverse arthropod communities compared to monoculture (Azhar et al. 2015b; Ghazali et al. 2016; Ashraf et al. 2018). The polyculture system's heterogeneity provides more food resources and habitat for various butterfly species (Collinge et al. 2003). Besides, the proximity of agricultural areas to forests increases the diversity of butterfly species (Lucey and Hill 2012; Koh 2008). On a local scale, butterfly species richness has been linked to ground vegetation cover and height (Azhar et al. 2015b; Asmah et al. 2017). This ground vegetation provides adults and immature butterflies with food and a breeding ground (Koh 2008).
Until now, butterfly research in Malaysian agroecosystems has been limited to oil palm plantations (Koh 2008; Lucey and Hill 2012; Azhar et al. 2015b; Asmah et al. 2017). No research has included butterfly species survey in rubber tree plantations or agroforestry orchards. As a result, farmland biodiversity in agroecosystems is poorly understood, and the role they play in agricultural landscapes in combination with either monoculture or polyculture farming systems, as well as encapsulating protected areas or forest reserves, is not well known.
In this study, we quantified butterfly presence and species richness in three different agricultural landscapes (agroforestry orchards, rubber tree plantations, and oil palm plantations) using point count methods. We also measured local- and landscape-level parameters and butterfly species distribution to answer three major questions;
-
1.
Does the diversity of butterfly species differ between agroforestry orchards, rubber tree plantations, and oil palm plantations?
-
2.
What local- and landscape-level factors are important in determining butterfly species richness in diverse agricultural landscapes?
-
3.
Are there any changes in butterfly community structure across agricultural landscapes?
This study is critical for promoting associated biodiversity in agricultural ecosystems, with a focus on butterflies as they play important ecological roles in the ecosystem (Brown and Freitas 2003) and are potentially affected by agricultural intensifications (Koh 2008; Lucey and Hill 2012). The study's findings will inform farmers and conservationists about the necessity of enhancing habitat quality at local- and landscape-level to increase biodiversity within agricultural landscapes.
Materials and methods
Study sites
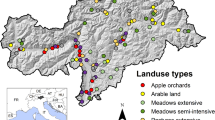
We conducted our research between Rembau District (2° 35′ 25.8900'' N, 102° 5′ 34.7532'' E), Negeri Sembilan, on the west coast of Peninsular Malaysia (Fig. 1). The study area (approximately 41,512 ha) was converted from lowland dipterocarp forests to agricultural areas at least 60 years ago (Denan et al. 2019). We selected this area because it has a diverse land use that includes oil palm plantations, rubber tree plantations, and agroforestry orchards. Both oil palm plantations and rubber tree plantations are monoculture system planted uniformly with a single species. For this study, oil palm and rubber tree plantations with an average age of 10 years old were selected for the sampling. Both monoculture plantations have rigorous management practices which involved systematic applications of chemical pesticides to control weeds, pests, and diseases (Azhar et al. 2015a). On the other side, agroforestry orchards were mainly made up of native fruit trees, with an average of five species of fruit trees planted at each orchard consisted mainly of durian (Durio zibethinus L.), rambutan (Nephelium lappaceum L.), langsat (Lansium domesticum Corr.), mangosteen (Garcinia mangostana L.), jering (Archidendron pauciflorum (Benth.) I. C. Nielsen), petai/ stink bean (Parkia speciosa Hassk.), and cempedak (Artocarpus integer Merr.). Typically, agroforestry orchards practice less intensive management approaches including mechanical removal of understory vegetation one or twice a year before fruiting seasons.

Map of study location in Pedas, Negeri Sembilan, Peninsular Malaysia. The left map indicates the location of the study site at Negeri Sembilan and the right map indicates the location of sampling point: yellow circle = agroforestry orchards, red square = oil palm plantation, and blue triangle = rubber tree plantations
Sampling design
Systematic sampling with a random starting point was used in this experiment to ensure better spatial coverage and lower variance (Morrison et al. 2008; Thomas et al. 2010). The butterfly and vegetation structure data were collected from 49 sampling points in oil palm plantations, 41 in rubber tree plantations, and 37 in agroforestry orchards. The distance among sampling points was at least 300 m apart to reduce the chances of counting the same individual at the sites. Each sampling point was visited once.
Butterflies sampling
Butterflies were observed using a point count method (Reynold et al. 1980) from February 2019 to February 2020. At each point, we recorded butterflies for 10 min in a 30 m plot radius from 9.00 a.m. to 3.00 p.m. using binoculars and sweep nets. Binocular (PENTAX Papilio 8.5 × 21) was used to detect the butterfly wing patterns. We identified the species of each butterfly using guidebooks by Kirton (2014) and Khoon (2015). Butterfly species that could not be identified were captured using a net and taken to the laboratory for species-level identification using a book by Corbet and Pandlebury (1992, 2020). Equal number of sites from each type of treatment were sampled each month. We counted the species richness of butterflies and categorized them according to their taxonomy (family, genus, and species) and distribution range (1- coastal mangrove, 2- secondary plant growth below 760 m, 3- primary forest below 760 m, and 4- all area above 760 m (Corbet and Pandlebury 2020)). We grouped butterfly species based on their distribution range into generalist or specialist category. Butterflies encountered in area 1 and 2 as well as area 2, 3 and 4 were classified as generalists, while those specifically found in area 3 and 4 were classified as specialists.
Measurement of local-level variables
In-situ habitat quality variables such as vegetation structure were recorded for each sampling within the area of 1000 m2. Variables measured at the local-level were: (i) tree height; (ii) number of tree species; (iii) diameter at breast height (dbh); (iv) canopy cover; (v) undergrowth coverage; (vi) undergrowth height; and (vii) elevation. Variables (iv), (v), and (vi) were measured in 1 m radius in four directions (east, north, west and south). We measured tree height and dbh using a laser range finder (Nikon LASER 350) and dbh meter, respectively. Tree species with more than 10 cm dbh were identified and recorded. We used HabitApp version 1.1 mobile application to measure canopy cover and Canopeo version 1.1.7 for undergrowth coverage, respectively. The undergrowth height was measured using measuring tape. We used GARMIN Global Positioning System (GPS) to provide the georeferenced location and determine the elevation at each sampling point.
Measurement of landscape-level variables
We measured the normalized difference vegetation index (NDVI) of the crops since butterfly species richness is related to the vegetation structure and productivity (Bailey et al. 2004). The NDVI was determined using ServEO version 1.3 mobile application. We also measured the distance between the sampling point and the nearest forest because butterfly species were affected by forest proximity especially for forest specialist (Lucey and Hill 2012). The nearest forests to our sites were Angsi Forest Reserve (12,435 ha), Sungai Menyala Forest Reserve (1,280 ha), and Gunung Tampin Forest Reserve (5,541 ha). The distance to the nearest forest was measured using Google Earth Pro measuring tools.
Measurement of seasonal variation
We measured the effect of seasonality (wet and dry) on butterfly species richness. We assigned sampling months into the wet and dry seasons based on data provided by MET Malaysia; wet seasons (southwest monsoon and inter-monsoon) between May and November, and dry seasons between December and April (MET 2019).
Statistical analysis
To compare butterfly species richness in agricultural landscapes, we used iNEXT online which a non-asymptotic approach based on interpolation and extrapolation (Colwell et al. 2012; Chao et al. 2016). Using the diversity order q = 0, we created sample-size-based rarefaction and extrapolation sampling curves (species richness). The default settings were used, with the number of bootstraps set to 50 and the level of confidence interval set to 95%.
The similarity percentage (SIMPER) analysis was used to determine the contribution of each species of butterfly assemblage in three different agricultural landscapes (i.e., agroforestry orchard, oil palm plantation and rubber tree plantation). The result showed the most persistence species in the pattern of similarity percentage. We used Bray–Curtis distance method to calculate the similarity metric between each landscape. The cutoff was set at 90% of the species accumulations.
We compared butterfly species composition between oil palm plantations, rubber tree plantations and fruit orchards using the analysis of similarity (ANOSIM). A resemblance matrix between samples was calculated using Bray–Curtis similarity. Percentages of similarity were used to determine the contribution of each butterfly species assemblage in three separate land uses. All multivariate analyses were performed in PRIMER Version 6 (PRIMER-E Ltd, Plymouth) (Clarke and Gorley 2006).
We used generalized linear mixed models (GLMMs) (Schall 1991) to examine the relationships between the richness of butterfly species and habitat quality characteristics (local and landscape-level variables). Spearman’s rank correlation coefficient tests were used to assess the multicollinearity of the explanatory variables. To prevent bias in model estimation, strongly correlated variables (|r|> 0.7) were checked (Dormann et al. 2013). However, none of the variables was strongly correlated. In the modelling process, Poisson distribution and log-link function were used with the number of butterfly species at each sampling point as the response variable and sampling month as a random model. Models with the lowest Mallow's Cp were identified as the most parsimonious models. The modeling was conducted using Genstat (VSNI Hemel, UK).
Results
General patterns of butterfly biodiversity
We recorded 1567 butterfly individuals comprising five families (Hespiriidae, Lycaenidae, Nymphalidae, Papilionidae, and Pieridae), 68 genera, and 104 species (Supplementary Information). Out of 104 butterfly species, 28 (27%) were specialists, while the remaining 76 (73%) were generalists. We also recorded two protected species in Malaysia under the Wildlife Conservation Act 2010 (Act 716), namely the banded peacock (Papilio palinurus) and common birdwing (Troides helena).
In agroforestry orchards, we recorded 472 butterfly individuals from 74 species (mean observation per point = 7.838) with 58 (78%) generalists and 16 (22%) forest specialists. For oil palm plantations, 615 butterflies from 54 species (7.102) were recorded with 44 (81%) generalists and 10 (19%) forest specialists while, for the rubber tree plantations we recorded 480 butterflies comprising 61 species (6.610) with 44 (72%) generalists and 17 (28%) forest specialists (Fig. 2). Of the total butterfly species observed, 31 were found across all agricultural landscapes, 25 exclusively in agroforestry orchards, 17 in rubber tree plantations, and only nine in oil palm plantations (Fig. 3). The rarefaction curves showed no overlap between three agricultural landscapes, suggesting that the observed differences in species diversity and number of individuals were not caused by varying sampling coverages (Fig. 4). Based on rarefaction and extrapolation results, our sampling effort managed to achieve 95% coverage of the butterfly communities across three agricultural landscapes (Table 1).

The percentages of butterfly species richness of generalist and specialist observed at three agricultural landscapes

Butterfly species richness and species overlap observed at three agricultural landscapes. The number in parenthesis represents the total number of species, and the square box represents species overlap in each habitat

Rarefaction curves of butterfly species at three different agricultural landscapes
SIMPER analysis (Supplementary Information) indicated 13 species of butterflies contributed to 90% of species composition for agroforestry orchards (average similarity = 22.46%) leading by Yptima baldus (26.71%), Appias libythea (10.09%), Eurema hecabae (9.10%), Junonia almana (7.94%) and Eurema sari (6.34%). For the oil palm plantations (average similarity = 26.29%), 11 species of butterflies contributed to 90% of species accumulation including Elymnias hypermnestra (16.23%), Leptosia nina (14.57%), Appias libythea (13.48%), Ypthima baldus (13.33%) and Eurema hecabe (10.60%). Eleven species of butterflies contributed to 90% of species accumulation in rubber tree plantations (average similarity = 21.34%) with Ypthima baldus (31.42%), Leptosia nina (10.78%), Appias libythea (8.72%), Eurema hecabe (8.54%) and Eurema sari (7.13%).
Overall, there was a significant difference of butterfly species composition with a high overlapping pattern (ANOSIM: number of permutations = 999; Global R = 0.064; p = 0.001). We found no significant difference in terms of butterfly species composition in agroforestry orchards and rubber tree plantations (ANOSIM R = 0.003; p = 0.387). Butterfly species composition in oil palm plantations was significantly different compared to agroforestry orchards (ANOSIM R = 0.075, p = 0.001) and rubber tree plantations (ANOSIM R = 0.099, p = 0.003).
Determinants of butterfly species richness
We found that eight predictor variables of tree height, undergrowth coverage, undergrowth height, elevation, season, NDVI, habitat, and distance to forest influenced butterfly species richness in three focal tropical agricultural landscapes. For the local level, a model with five predictor variables (tree height, undergrowth coverage, undergrowth height, elevation, and season) was the most parsimonious combination with the lowest Mallow's Cp of 5.55 (Table 2; Fig. 5). The model explained 30.23% of the variation in butterfly species richness. Our data revealed that butterfly species richness increased with increasing undergrowth coverage (slope = 0.007681) and undergrowth height (slope = 0.4057) (Table 3). On the other hand, we found out that butterfly species richness decreased with increasing tree height (slope = – 0.03052) and elevation (slope = – 0.002891) (Table 3). Butterfly species richness was lower during the wet season compared to the dry season (slope = – 0.2038) (Table 3).

Different responses to local-level parameters by butterfly species richness. Scatter plots have 95% confidence intervals (blue line) on the regression (red line)
At the landscape level, a model with four predictor variables (distance to forest, NDVI, habitat, and season) was the most parsimonious combination with the lowest Mallow's Cp of 22.52 (Table 2; Fig. 6). The model explained 20.41% of the variation in butterfly species richness of butterfly. We found that butterfly species richness increased as the distance to the forest decreased (slope = – 0.042) (Table 3). The species richness had a negative relationship with the NDVI value (slope = – 0.5194). Oil palm plantations (slope = – 0.3109) and rubber tree plantations (slope = – 0.1988) supported lower species richness in comparison to agroforestry orchards (Table 3). During the wet season (slope = – 0.2341), butterfly species richness was lower than in the dry season (Table 3).

Different responses to landscape level parameters by butterfly species richness. Scatter plots have 95% confidence intervals (blue line) on the regression (red line)
Discussion
General patterns of butterfly diversity
There are about 1051 species of butterflies recorded in Peninsular Malaysia (Corbet and Pendelbury 2020) and in this study we recorded a total of 104 species of butterflies, which representing approximately 10% of the total butterfly species reported. This finding is consistent with previous research indicating that the richness of butterfly species is impoverished in tropical agricultural landscapes (Koh 2008; Azhar et al. 2015b; Asmah et al. 2017) compared to forest areas (Willott et al. 2000; Benedick et al. 2006). This may indicate that only a small proportion of butterfly species is able to persist well in agricultural landscapes, especially generalist species, which represent about 73%, followed by forest specialists at 27%.
We found that the richness of butterfly species was higher in agroforestry orchards in comparison to rubber tree and oil palm plantations. This finding showed that agroforestry orchards with polyculture practices would offer better habitats for butterflies compared to rubber tree and oil palm plantations that are monocultures. In our case, agroforestry orchards were integrated with more than 5 species of fruit tree species consisting mainly of large-sized trees (e.g., durian, jering, petai), as well as other medium-sized trees (e.g., rambutan, mangosteen) forming a more complex habitat compared to monocultures. As such, polyculture systems improve habitat heterogeneity and vegetation structure (Horak et al. 2014; Elings et al. 2017) and are expected to provide more resources for an insect (Jones and Gillet 2005; Elba et al. 2014). Agroforestry systems maintain biodiverse habitat and landscape heterogeneity (Bhagwat et al. 2008; Yahya et al. 2022), in contrast to large-scale monoculture plantations, which have a lower potential for biodiversity conservation due to lower heterogeneity (Azhar et al. 2015a). Native plants provide unlimited food resources for butterflies (Burghardt et al. 2009) as such, integrating multiple species of fruit trees would facilitate more butterfly species within an agriculture landscape (Asmah et al. 2017). We also observed the Painted Jezebel butterfly (Delias hyparete) feeding on rambutan fruit in the agroforestry orchards.
However, the species composition of butterflies between agroforestry orchard and rubber plantations are similar but different to oil palm plantations. The similarity of species richness is likely related to management practices, as most of the crops belonged to small-scale growers, except for a few rubber tree sites owned by the plantation company. On the other hand, oil palm plantations are mainly owned by plantation companies. Small-scale farmers typically practice less intensive management and are less dependent on modern machinery (Azhar et al. 2015a). This means that practices such as chemical weeding and agrochemical applications are rarely consistent compared to large-scale plantations. As a result, it will enhance the growth of understory vegetation and provide suitable habitats and food resources for the butterflies. Butterflies utilized (i.e., feed and breed) on naturally grown understory vegetation across the agricultural landscape, regardless of land use. Broadleaved shrubs and tree (whiteweed—Ageratum conyzoides, Chinese violet—Asystasia gangetica, rhododendron—Melastoma malabathricum, Soapbush—Clidemia hirta) and grasses (wiregrass- Eleusine indica, congongrass—Imperata cylindrica) are the common weed species in the Malaysian agricultural landscape in (Barnes and Chan 1990; Maziatul-Suriza and Idris 2012; Fee et al. 2017). Broadleaved shrubs and tree provide a source of nectar for low-flying butterflies such as Psyche, Rings (Ypthima), Grass yellows and others. Grass is an important breeding host for Rings (Ypthima) since their caterpillars feed on grass (Poaceace) (Corbet and Pandelbury 2020). Thus, this contributed to the higher abundance of the Y. baldus in agricultural landscapes. On the other hand, the abundance of E. hypermnestra, a common species in oil palm plantations (Kirton 2014) which is associated with various palm species (Corbet and Pandelbury 2020). Oil palm plantations support fewer animal species than forest and other tree crops (Fitzherbert et al. 2008) due to intensive management practices such as the use of agrochemicals to increase yields (Azhar et al. 2015a).
Oil palm and rubber tree plantations support lower butterfly species richness compared to agroforestry orchards. This finding implies that agroforestry orchards would provide the butterfly with better habitat and food resources than oil palm and rubber tree plantations. Rubber tree and oil palm plantations are primarily renowned for a monoculture system with uniform stand age, which is unfavorable to farmland biodiversity (Ghazali et al. 2016; Yahya et al. 2022). The structure and composition of vegetation influence the diversity of butterflies (Kremen et al. 1993; New et al. 1995). The uniform canopy strata in monoculture plantations are unfavorable to butterflies due to their natural behavior of inhibiting different canopy strata of trees (Asmah et al. 2017). Butterflies of the Morphinae and Satyrinae subfamilies only fly in the understory on the lower vegetation layer (Schulze et al. 2001) while Apaturinae, Charaxinae and Nymphalinae subfamilies can be found flying on the upper canopy layer (DeVries and Walla 2001). However, Apaturinae and Charaxinae were not detected by our sampling method (i.e., point count), as both are forest specialists and attracted to rotten fruit (Corbet and Pandlebury 2020). Aside from that, ground cover vegetation in fruit orchards may provide a food source and breeding ground for butterflies. The majority of butterflies in Southeast Asia rely on monocotyledonous host plants for breeding and feeding (Schulze et al. 2001; Corbet and Pandlebury 2020). Aside from that, agroforestry orchards exhibit different phenology during the flowering season providing a constant supply of nectar for butterflies and other nectivorous insects (Elings et al. 2017).
Butterfly species richness and influencing attributes
Our data suggested that the tree height negatively affects the richness of the butterfly species. A similar result was also recorded in previous studies, which reported a negative association between tree height and butterfly diversity (e.g., Kumar et al. 2009; Wettstein and Schmid 1999; Thomas and Mallorie 1985). The biology and physiology of butterflies may have contributed to this negative relationship since butterflies are poikilothermic species. The solar radiation needed by butterflies for thermoregulation may be restricted by the shading of high trees (Kumar et al. 2009).
Our findings indicate that undergrowth coverage and undergrowth height had a positive impact on the richness of the butterfly species. This positive association is expected since undergrowth vegetation provide resources for the butterflies. Similar findings were reported by Azhar et al. (2015b), who discovered that decreases in ground vegetation structure and coverage reduced butterfly species richness. Both adult and larvae of butterflies require a certain host plant that serves as food source and their richness is closely linked to host plant distribution and nectar availability (Collinge et al. 2003). The diversity of butterflies is also affected by the structure and composition of ground vegetation (Kremen et al. 1993; New et al. 1995). Most of the butterflies, particularly those in the Morphinae and Satyrinae subfamilies, are flying at low to moderate height (Schulze et al. 2001; Kumar et al. 2009) to find their resources (e.g., host plants, mates and foods) (Vane-Wright 2015).
A negative relationship was observed between butterfly species richness and elevation. This negative association was expected since butterfly diversity, species richness, and species abundance are generally higher in low-elevation habitats than in high-elevation habitats (Lien and Yuan 2003). Several factors contribute to this negative relationship, including a decreasing trend in plant species richness and habitat heterogeneity, which decreases with elevation and eventually leads to lower insect diversity (Lieberman et al. 1996; Grytnes and Beaman 2006; Achrya and Vijayan 2015). Moreover, butterfly distributions in Malaysia are constrained by altitude and plant associations, with approximately half of all butterfly species occurring below 750 m above sea level (asl) (Corbet and Pandelbury 2020). In addition, at elevations above 1000 m (asl), there are fewer butterfly species, but butterfly species that exist at higher elevations do not live at lower levels (Kirton 2014).
During the wet season, butterfly diversity is lower as rainfall affects humidity, temperature, and local microclimate (Speight et al. 2008). Most of the butterflies tend to be inactive during this period since low temperatures and precipitation may limit their activities. Butterflies are cool-blooded organisms. Even small changes in ambient temperature affect their abundance, foraging activity, mating, and physiology (Kearns 2012; Klok 2013). Aside from that, heavy rain can impact the survival of butterfly larvae and pupae (Hill et al. 2003). Thus, lower butterfly species richness during the wet season is caused by environmental factors that are not measured in this study.
The mean NDVI and butterfly species richness were found to have a negative relationship. In this study, the mean NDVI in oil palm plantations was the highest followed by the rubber plantations, but the lowest when it came to butterfly species richness. Such results are expected, chemical fertilizers are systematically used in conventional plantations, especially in oil palm plantations to boost yield productions where more than 85% of the operation cost went to fertilizers alone (Goh 2005). Fertilizer increases the chlorophyll content of plants, influencing the NDVI reading (Gómez et al. 2019). Aside from that, the phenological difference in rubber tree plantations may influence the NDVI reading especially during foliation stages resulting in higher NDVI reading (Yeang 2007; Janatul et al. 2018; Hazir et al. 2020). Rubber trees produce new leaves during this period, and the canopy recovers from defoliation, increasing the NDVI value (Dong et al. 2013; Janatul et al. 2018).
Our study also showed that species richness of the butterfly increased with decreasing distance to the nearest forest as the majority of the butterfly species observed in this study are associated with forest habitat. This finding aligned with Lucey and Hill (2012), and Panjaitan (2020), which reported that the spillover of butterflies of forest specialists from the nearest forest will improve diversity in the agriculture areas. Natural forests and secondary forest remnants adjacent to plantations serve as an important source of complementary breeding and feeding for butterfly species, especially forest dwellers, due to the availability of host plants (Koh 2008), and contribute to conservation on the landscape level (Veddeler et al. 2005).
Our findings suggested that ground vegetation cover and proximity to a forest were important in supporting butterfly species richness in agricultural landscapes. To improve butterfly diversity, the common practices in mainstream agriculture such as using synthetic herbicides for controlling overgrown shrubs and weeds should be reduced. Alternatively, livestock grazing which is more environmentally friendly can be integrated with the existing agricultural landscapes (Tohiran et al. 2017, 2019; Azhar et al. 2017, 2021). Aside from that, preserving forest areas within agricultural landscapes is critical for facilitating butterfly dispersal and mitigating species loss due to land conversion (Lucey and Hill 2012; Lucey et al. 2014; Panjaitan et al. 2020). Small-scale farmers should practice alley-cropping system to improve biodiversity on their farmlands. This strategy could enhance the floristic composition and stand structural complexity for the arthropods, which eventually reduce the microclimate effects in the farmlands (Ashraf et al. 2018, 2019). Based on our data, agroforestry orchards showed the significance of habitat quality to boost biodiversity in agricultural landscapes and served as an example for key stakeholders in Southeast Asia's oil palm and rubber tree sector to improve biodiversity and ecosystem services in conventional plantations.
Our observational data are restricted to point counts of butterfly species. Other sampling methods, such as bait trapping, should be considered to obtain a comprehensive picture of the butterfly species community in future studies (Checa et al. 2019; Hebel et al. 2022). However, due to disturbance from long-tailed macaques (Macaca fascicularis), bait trapping is not appropriate for use in our study area. Aside from that, future research on butterfly species communities in agricultural landscapes should include other factors that may influence butterflies, such as the number of blossoms, water availability, host plants, and rotten fruits.
Conclusions
Based on the evidence presented in this study, tropical agricultural landscapes, particularly agroforestry orchards are instrumental for biodiversity conservation outside protected areas. Conservationists must consider the conservation value of agroforestry orchards for biodiversity to make conservation programs more effective. Because agroforestry orchards affect a substantial portion of the terrestrial region in Southeast Asia amid monoculture plantations, their contribution to biodiversity is critical for future conservation efforts.
Data availability
Data generated from this study will be provided upon reasonable request by the corresponding author.
References
Abdullah SA (2011) The characteristics of the cultural landscape in malaysia: concept and perspective. In: Hong SK., Kim JE., Wu J., Nakagoshi N (Eds) Landscape ecology in Asian cultures. Ecological research monographs. Springer, Tokyo. https://doi.org/10.1007/978-4-431-87799-8_4
Acharya BK, Vijayan L (2015) Butterfly diversity along the elevation gradient of Eastern Himalaya, India. Ecol Res 30:909–919. https://doi.org/10.1007/s11284-015-1292-0
Ahrends A, Hollingsworth PM, Ziegler AD, Fox JM, Chen H, Su Y, Xu J (2015) Current trends of rubber plantation expansion may threaten biodiversity and livelihoods. Glob Environ Chang 34:48–58
Alroy J (2017) Effects of habitat disturbance on tropical forest biodiversity. Proc Natl Acad Sci 114:6056–6061
Ashraf M, Zulkifli R, Sanusi R, Tohiran KA, Terhem R, Moslim R et al (2018) Alley-cropping system can boost arthropod biodiversity and ecosystem functions in oil palm plantations. Agr Ecosyst Environ 260:19–26
Ashraf M, Sanusi R, Zulkifli R, Tohiran KA, Moslim R, Ashton-Butt A, Azhar B (2019) Alley-cropping system increases vegetation heterogeneity and moderates extreme microclimates in oil palm plantations. Agric for Meteorol 276:107632
Asmah S, Ghazali A, Syafiq M, Yahya MS et al (2017) Effects of polyculture and monoculture farming in oil palm smallholdings on tropical fruit-feeding butterfly diversity. Agric for Entomol 19:70–80
Athukorala P-C, Loke W-H (2009) Agricultural incentives in Malaysia: trends, patterns and policy implications. Malays J Econ Stud 46(2):151–173
Azhar B, Saadun N, Puan CL, Kamarudin N, Aziz N, Nurhidayu S, Fischer J (2015a) Promoting landscape heterogeneity to improve the biodiversity benefits of certified palm oil production: evidence from Peninsular Malaysia. Glob Ecol Conserv 3:553–561. https://doi.org/10.1016/j.gecco.2015.02.009
Azhar B, Puan CL, Aziz N et al (2015b) Effects of in situ habitat quality and landscape characteristics in the oil palm agricultural matrix on tropical understory birds, fruit bats and butterflies. Biodivers Conserv 24:3125–3144. https://doi.org/10.1007/s10531-015-1005-6
Azhar B, Saadun N, Prideaux M, Lindenmayer DB (2017) The global palm oil sector must change to save biodiversity and improve food security in the tropics. J Environ Manage 203:457−466
Azhar B, Nobilly F, Lechner AM, Tohiran KA, Maxwell T et al (2021) Mitigating the risks of indirect land use change (ILUC) related deforestation from industrial palm oil expansion by sharing land access with displaced crop and cattle farmers. Land Use Policy 107:105498. https://doi.org/10.1016/j.landusepol.2021.105498
Bailey SA, Horner-Devine MC, Luck G, Moore LA, Carney KM, Anderson S et al (2004) Primary productivity and species richness: relationships among functional guilds, residency groups and vagility classes at multiple spatial scales. Ecography 27(2):207–217
Barnes DE, Chan LG (1990) Common weeds of Malaysia and their control. Ancom Berhad
Barnes AD, Jochum M, Mumme S, Haneda NF, Farajallah A, Widarto TH, Brose U (2014) Consequences of tropical land use for multitrophic biodiversity and ecosystem functioning. Nat Commun 5:5351. https://doi.org/10.1038/ncomms6351
Benedick H, Hill K, Mustaffa N, Chey VK, Maryati M, Searle B, Schilthuizen M, Hamer KC (2006) Impacts of rain forest fragmentation on butterflies in northern Borneo: species richness, turnover and the value of small fragments. J Appl Ecol 43:967–977. https://doi.org/10.1111/j.1365-2664.2006.01209.x
Bhagwat SA, Willis KJ, Birks HJB, Whittaker RJ (2008) Agroforestry: a refuge for tropical biodiversity? Trends Ecol Evol 23:261–267
Brown KS, Freitas AVL (2003) Butterfly communities of urban forest fragments in Campinas, São Paulo, Brazil: structure, instability, environmental correlates, and conservation. J Insect Conserv 6:217–231
Burghardt KT, Tallamy DW, Gregory SW (2009) Impact of native plants on bird and butterfly biodiversity in suburban landscapes. Conserv Biol 23(1):219–224
Checa MF, Donoso DA, Rodriguez J, Levy E, Warren A, Willmott K (2019) Combining sampling techniques aids monitoring of tropical butterflies. Insect Conserv Diversity 12(4):362–372
Chao A, Ma KH, Hsieh TC (2016) User’s guide for iNEXT online: Software for interpolation and Extrapolation of species diversity. Code 30043(September):1–14
Clarke KR, Gorley RN (2006) User Manual/Tutorial. PRIMER‐E Ltd, U.K
Cleary DFR (2004) Assessing the use of butterflies as indicators of logging in Borneo at three taxonomic levels. J Econ Entomol 97(2):429–435
Collinge SK, Prudic KL, Oliver JC (2003) Effects of local habitat characteristics and landscape context on grassland butterfly diversity. Conserv Biol 1:178–187
Colwell RK, Chao A, Gotelli NJ, Lin SY, Mao CX, Chazdon RL, Longino JT (2012) Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J Plant Ecol 5(1):3–21
Corbet AS, Pendlebury HM (1992) The butterflies of the Malay Peninsula, 4th revised edition by Eliot JN. Malayan Nature Society, Kuala Lumpur
Corbet AS, Pendlebury HM (2020) The Butterflies of the Malay Peninsula, 5th edition, revised by George van der Poorten & Nancy van der Poorten. Malayan Nature Society, Kuala Lumpur
Díaz S, Fargione J, Chapin FS III, Tilman D (2006) Biodiversity loss threatens human well-being. PLoS Biol 4(8):e277. https://doi.org/10.1371/journal.pbio.0040277
Denan N, Wan Zaki WM, Norhisham AR, Sanusi R et al (2019) Predation of potential insect pests in oil palm plantations, rubber tree plantations, and fruit orchards. Ecol Evol. https://doi.org/10.1002/ece3.5856
Department of Statistics Malaysia (DOSM) (2019) Key data-fruit crop statistics Malaysia 2019. https://www.dosm.gov.my. Accessed 20 July 2020
De Vries PJ, Walla TR (2001) Species diversity and community structure in neotropical fruit-feeding butterflies. Biol J Lin Soc 74(1):1–15
Dong J, Xiao X, Chen B, Torbick N, Zhang G, Biradar C, Jin C (2013) Mapping deciduous rubber plantations through integration of PALSAR and multi-temporal Landsat imagery. Remote Sens Environ 134:392–402
Dormann CF, Elith J, Bacher S et al (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Elba B, Suárez SA, Lenardis AE, Poggio SL (2014) Intercropping sunflower and soybean in intensive farming systems: evaluating yield advantage and effect on weed and insect assemblages. NJAS-Wageningen J Life Sci 70:47–52
Elings J, Kirimbo G, Li X, Mandal P (2017) Enhancing biodiversity in traditional fruit orchards. Ecological Consultancy Bureau. Wageningen University, Red Apple
Ekroos J, Kuussaari M, Tiainen J, Heliölä J, Seimola T, Helenius J (2013) Correlations in species richness between taxa depend on habitat, scale and landscape context. Ecol Ind 34:528–535. https://doi.org/10.1016/j.ecolind.2013.06.015
Ewers RM, Scharlemann JPW, Balmford A, Green RE (2009) Do increases in agricultural yield spare land for nature? Glob Chang Biol 15:1716–1726
Fee CG, Tui LC, Bin CS, Hoy CK (2017) Pictorial guide to common weeds of plantations and their control (No. L-0987), 2nd ed. Agricultural Crop Trust, ACT, Bonn
Fitzherbert EB, Struebig MJ, Morel A, Danielsen F, Brühl CA, Donald PF, Phalan B (2008) How will oil palm expansion affect biodiversity? Trends Ecol Evol 23(10):538–545
Ghazali A, Asmah S, Syafiq M et al (2016) Effects of monoculture and polyculture farming in oil palm smallholdings on terrestrial arthropod diversity. J Asia-Pacific Entomol 19:415–421
Giam X (2017) Global biodiversity loss from tropical deforestation. Proc Natl Acad Sci USA 114:5775–5777. https://doi.org/10.1073/pnas.1706264114
Goh KJ (2005) Fertilizer recommendation systems for oil palm: estimating the fertilizer rates. In: Proceedings of MOSTA best practices workshops-agronomy and crop management. Malaysian Oil Scientists’ and Technologists’ Association, pp 1–37
Gómez MI, Barragán A, Magnitskiy S, Rodríguez LE (2019) Normalized difference vegetation index, and K+ in stem sap of potato plants (Group Andigenum) as affected by fertilization. Exp Agric 55(6):945–955
Grytnes JA, Beaman JH (2006) Elevational species richness patterns for vascular plants on Mount Kinabalu, Borneo. J Bio- Geogr 33:1838–1849
Habel JC, Ulrich W, Eberle J, Schmitt T (2022) Species community structures of Afrotropical butterflies differ depending on the monitoring method. Biodiversity Conserv 31:1–15
Hazir MHM, Kadir RA, Gloor E, Galbraith D (2020) Effect of agroclimatic variability on land suitability for cultivating rubber (Hevea brasiliensis) and growth performance assessment in the tropical rainforest climate of Peninsular Malaysia. Climate Risk Manag. https://doi.org/10.1016/j.crm.2019.100203
Hill J, Hamer K, Dawood MM, Tangah J, Chey V (2003) Rainfall but not selective logging affect changes in abundance of a tropical forest butterfly in sabah. Borneo J Trop Ecol 19(1):35–42
Horak J (2014) Fragmented habitats of traditional fruit orchards are important for dead wood-dependent beetles associated with open canopy deciduous woodlands. Naturwissenschaften 101:499–504
Janatul AAR, Abdul RMS, Noordin A, Maher IS (2018) Mapping rubber trees based on phenological analysis of Landsat time series data-sets. Geocarto Int 33(6):627–650. https://doi.org/10.1080/10106049.2017.1289559
Jones GA, Gillett JL (2005) Intercropping with sunflowers to attract beneficial insects in organic agriculture. Florida Entomol 88:91–96
Karen-Chia HM (2014) Exploring the diversity of butterflies (lepidoptera) at different elevations in Genting Highlands and the validity of graphium species in Peninsular Malaysia. Doctoral Dissertation, University of Malaya
Kearns P, Stevenson RD (2012) Effect of decreasing temperature on Arthropod diversity and abundance in horse dung decomposition communities of South-Eastern Massachusetts. Psyche 2012:1–12
Khoon KS (2015) A Field Guide to the Butterflies of Singapore, 2nd edn. Ink On Paper Communications Pte Ltd., Singapore
Kirton LG (2014) A Naturalist’s Guide to the Butterflies of Peninsular Malaysia. John Beaufoy Publishing Limited, Oxford, Singapore and Thailand
Klok CJ, Harrison JF (2013) The temperature size rule in arthropods: independent of macro-environmental variables but size dependent. Integr Comp Bio 53:55–70
Koh LP (2008) Can oil palm plantations be made more hospitable for forest butterflies and birds? J Appl Ecol 45:1002–1009
Kremen C, Colwell RK, Erwin TL, Murphy DD, Noss RF, Sanjayan MA (1993) Terrestrial arthropod assemblages: Their use in conservation planning. Conserv Biol 7:796–808
Kumar S, Simonson SE, Stohlgren TJ (2009) Effects of spatial heterogeneity on butterfly species richness in Rocky Mountain National Park, CO, USA. Biodivers Conserv 18:739–763
Landis DA (2017) Designing Agricultural Landscapes for Biodiversity-Based Ecosystem Services. Basic Appl Ecol 18:1–12
Lieberman D, Lieberman M, Peralta R, Hartshorn GS (1996) Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. J Ecol 84:137–152
Lien VV, Yuan D (2003) The differences of butterfly (Lepidoptera, Papilionoidea) communities in habitats with various degrees of disturbance and altitudes in tropical forests of Vietnam. Biodivers Conserv 12:1099–1111. https://doi.org/10.1023/A:1023038923000
Lucey JM, Hill JK (2012) Spillover of Insects from Rain Forest into Adjacent Oil Palm Plantations. Biotropica 44(3):368–377. https://doi.org/10.1111/j.1744-7429.2011.00824.x
Lucey JM, Tawatao N, Senior MJ, Chey M et al (2014) Tropical forest fragments contribute to species richness in adjacent oil palm plantations. Biol Cons 169:268–276. https://doi.org/10.1016/j.biocon.2013.11.014
Maizatul-Suriza M, Idris AS (2012) Occurrence of common weeds in immature plantings of oil palm plantations in Malaysia. The Planter 88(1037):537–547
MET (2019) Malaysian Meteorological Department Annual Report 2019 https://www.met.gov.my/content/pdf/penerbitan/laporantahunan/laporantahunan2019.pdf (accessed 14.09.2022)
Ministry of Primary Industry of Malaysia (MPIC) (2015) MPIC Annual Report 2015. www.mpic.gov.my. Accessed 20 July 2020
Morrison LW, Smith DR, Young CC, Nichols DW (2008) Evaluating sampling designs by computer simulation: a case study with the Missouri bladderpod. Popul Ecol 50:417–425
Nath S, Chaudhuri PS (2010) Human – induced biological invasions in rubber (Hevea brasiliensis) plantations of Tripura (India) – Pontoseolex corethrurus as a case study. Asian J Experim Biol Sci 1:360–369
New TR, Pyle RM, Thomas JA, Thomas CD, Hammond PC (1995) Butterfly conservation management. Annu Rev Entomol 40:57–83
Panjaitan R, Drescher J, Buchori D, Peggie D, Harahap IS, Scheu S, Hidayat P (2020) Diversity of butterflies (Lepidoptera) across rainforest transformation systems in Jambi, Sumatra, Indonesia. Biodiversitas 21:5119–5127
Pashkevich MD, Aryawan AAK, Luke SH, Dupérré N et al (2020) Assessing the effects of oil palm replanting on arthropod biodiversity. J Appl Ecol. https://doi.org/10.1111/1365-2664.13749
Pogue DW, Schnell GD (2001) Effects of agricultural on habitat complexity in a prairie-forest ecotone in the Southern Great Plains of North America. Agric Ecosyst Environ 87:287–298
Posa MRC, Sodhi NS (2006) Effects of anthropogenic land use on forest birds and butterflies in Subic Bay, Philippines. Biol Conserv 129:256–270
Potapov AM, Dupérré N, Jochum M, Dreczko K, Klarner B et al (2020) Functional losses in ground spider communities due to habitat structure degradation under tropical land-use change. Ecology. https://doi.org/10.1002/ecy.2957
Power AG (2010) Ecosystem services and agriculture: tradeoffs and synergies. Phil Trans r Soc B 365:2959–2971
Reynolds RT, Scott JM, Nussbaum RA (1980) A Variable Circular-Plot Method for Estimating Bird Numbers. The Condor Ornithol Appl 82(3):309–313. https://doi.org/10.2307/1367399
Rozhan AD (2015) Transformation of agricultural sector in Malaysia through agricultural policy. FFTC Agricultural Policy. www.ap.fftc.agnet.org.
Salma I, Masrom AM, Raziah ML (2006) Diversity and use of traditional fruit species in selected home gardens or fruit orchards in Malaysia. J Trop Agric Food Sci 34(1):149–164
Sayer J, Ghazoul J, Nelson P, Boedhihartono AK (2012) Oil palm expansion transforms tropical landscapes and livelihoods. Glob Food Sec 1(2):114–119
Šálek M, Hula V, Kipson M, Daňková R, Niedobová J, Gamero A (2018) Bringing diversity back to agriculture: smaller fields and non-crop elements enhance biodiversity in intensively managed arable farmlands. Ecol Indic 90:65–73
Schall R (1991) Estimation in generalized linear-models with random effects. Biometrika 78:719–727
Schulze CH, Linsenmair KE, Fiedler K (2001) Understory versus canopy: patterns of vertical stratification and diversity among Lepidoptera in a Bornean rain forest. In: Linsenmair KE, Davis AJ, Fiala B, Speight MR (Eds) Tropical Forest Canopies: Ecology and Management. Forestry Sciences 69. Springer, Dordrecht. https://doi.org/10.1007/978-94-017-3606-0_11
Schwarz J, Schnabel F, Bauhus J (2021) A conceptual framework and experimental design for analysing the relationship between biodiversity and ecosystem functioning (BEF) in agroforestry systems. Basic Appl Ecol 55:133–151. https://doi.org/10.1016/j.baae.2021.05.002
Speight MR, Hunter MD, Watt AD (2008) Ecology of Insects: Concepts and Applications. Blackwell Science Ltd, Oxford
Stanturf JA, Kleine M, Mansourian S et al (2019) Implementing forest landscape restoration under the Bonn Challenge: a systematic approach. Ann for Sci 76:50. https://doi.org/10.1007/s13595-019-0833-z
Stokes DW, Stokes LQ, Williams E (1991) The butterfly book: an easy guide to butterfly gardening, identification, and behavior. Little, Brown and Company, Boston
Thomas CD, Mallorie HC (1985) Rarity, species richness and conservation: butterflies of the Atlas Mountains in Morocco. Biol Conserv 33(2):95–117
Thomas L, Buckland ST, Rexstad EA, Laake JL et al (2010) Distance software: design and analysis of distance sampling surveys for estimating population size. J Appl Ecol 47(1):5–14
Tohiran KA, Nobilly F, Zulkifli R, Maxwell T, Moslim R, Azhar B (2017) Targeted cattle grazing as an alternative to herbicides for controlling weeds in bird-friendly oil palm plantations. Agron Sustain Dev 37:62. https://doi.org/10.1007/s13593-017-0471-5
Tohiran KA, Nobilly F, Zulkifli R, Ashton-Butt A, Azhar B (2019) Cattle-grazing in oil palm plantations sustainably controls understory vegetation. Agric Ecosyst Environ 278:54–60. https://doi.org/10.1016/j.agee.2019.03.021
Vane-Wright D (2015) Butterflies a complete guide to their biology and behaviour. Natural History Museum, London
Veddeler D, Schulze CH, Steffan-Dewenter I, Buchori D, Tscharntke T (2005) The contribution of tropical secondary forest fragments to the conservation of fruit-feeding butterflies: effects of isolation and age. Biodivers Conserv 14:3577–3592. https://doi.org/10.1007/s10531-004-0829-2
Warren-Thomas E, Nelson L, Juthong W, Bumrungsri S, Brattström O, Stroesser L, Dolman PM (2020) Rubber agroforestry in Thailand provides some biodiversity benefits without reducing yields. J Appl Ecol 57:17–30
Wettstein W, Schmid B (1999) Conservation of arthropod diversity in montane wetlands: effect of altitude, habitat quality and habitat fragmentation on butterflies and grasshoppers. J App Ecol 36(3):363–373
Willott SJ, Lim DC, Compton SG, Sutton SL (2000) Effects of selective logging on the butterflies of a Bornean rainforest. Conserv Biol 14(4):1055−1065
Yahya MS, Atikah SN, Mukri I, Sanusi R, Norhisham AR, Azhar B (2022) Agroforestry orchards support greater avian biodiversity than monoculture oil palm and rubber tree plantations. For Ecol Manage 513:120177
Yeang HY (2007) Synchronous flowering of the rubber tree (Hevea brasiliensis) induced by high solar radiation intensity. New Phytol 175(2):283–289
Acknowledgements
We thank the anonymous plantation company and smallholders in Rembau, Negeri Sembilan, for granting permission to conduct the research on their farms. We also would like to express our gratitude to Rutger Sterenborg, Mizan Mat Nawi, Syafiq Salehhuddin, Arief Aiman Arman, Nuradilah Denan, Ku Noor Khalidah Ku Halim, Nuurfarahani Zulkifli, Sharifah Nur Atikah, and Kamaliah Eddy for their assistance during the fieldwork.
Funding
This research was funded by Universiti Putra Malaysia under Putra Young Initiative Gant (IPM) (Project No: 9637800).
Author information
Authors and Affiliations
Contributions
W.Z.W.M., M.S.Y., A.R.N., R.Z., P.J.v.d.M., and B.A. conceived the ideas, designed methodology, and administered the research; W.Z.W.M. collected the data in the field and identified the samples, W.Z.W.M., M.S.Y. and B.A. analyzed the data, and drew the figures; W.Z.W.M., M.S.Y., A.R.N., R.Z., P.J.v.d.M., and B.A. led the writing of the manuscript. All authors contributed critically to the manuscript and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Communicated by Sebastian Seibold.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Wan Zaki, W.M., Yahya, M.S., Norhisham, A.R. et al. Agroforestry orchards support greater butterfly diversity than monoculture plantations in the tropics. Oecologia 201, 863–875 (2023). https://doi.org/10.1007/s00442-023-05348-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-023-05348-3