
Abstract
Purpose
The transcription factor Fos-related antigen-1 (Fra-1) has been described to affect the morphology, motility and invasive potential of breast cancer cells. Since tumor cell adhesion plays an essential role in the metastatic process, especially for extravasation from blood vessels, we investigated the influence of Fra-1 on breast cancer cell interactions with the endothelium.
Methods
Using Fra-1-overexpressing MCF7 [weakly invasive, estrogen receptor (ER)-positive] and MDA MB231 (strongly invasive, ER-negative) cells, we performed dynamic cell flow adhesion assays on surfaces coated with E-selectin or with human pulmonary microvascular endothelial cells.
Results
We found a significant increased adhesion of Fra-1-overexpressing MCF7 cells to E-selectin but also to activate endothelial cells, whereas the MDA MB231 cell line showed moderate enhanced cell rolling and tethering on both coated surfaces. These different adhesion behaviors corresponded to an up-regulation of various adhesion-related proteins such as CD44 and integrin α5 in Fra-1-overexpressing MCF7 cells measured by microarray analysis and flow cytometry in comparison with no deregulation of key adhesion molecules observed in Fra-1-overexpressing MDA MB231 cells. In line with these results and based on cDNA microarray data of breast cancer patients (n = 197), high Fra-1 expression significantly correlates with shorter overall survival and higher rate of lung metastasis in ER-positive breast cancer patients (n = 130), but has no impact on the prognosis of patients with ER-negative tumors.
Conclusion
Thus, in addition to its pro-invasive and pro-migratory effect, Fra-1 might influence the metastatic potential of breast cancer cells by changing the expression of adhesion molecules, resulting in increased adherence to endothelial cells under flow conditions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the leading causes of death in women is breast cancer. It is a heterogeneous disease, and the most important behavior of breast cancer cells is the early lymphogenous and hematogenous metastasis. On the one hand, metastatic dissemination is an inefficient process as only a small subset of cancer cells in a primary tumor has the potential to form metastases; on the other hand, metastasis is responsible for around 90 % of cancer patient mortality. The process of metastasis includes many steps including invasion of the basal membrane and neighboring tissues by cancer cells, intravasation into the vasculature or infiltration of lymphatic vessels, survival in the circulation, extravasation, distant organ infiltration and colonization. These mechanisms require a precise coordination of various signaling pathways and strategies used by the cancer cells to acquire the ability to change their adhesive properties. Specially, the exit of tumor cells from capillary beds into the parenchyma of an organ is strongly affected by their interactions with vascular endothelial cells. Cancer cells have to slow down and then form stable attachment. These processes are mediated by a wide range of ligands and receptors, including selectins, integrins, cadherins, CD44 and immunoglobulin (Ig) superfamily receptors.
The activator protein-1 complex, AP-1, is a key regulator of transcriptional responses induced by various cancer-associated signaling pathways. It consists either of homodimers of Jun family members (c-Jun, JunB and JunD) or heterodimers of Jun proteins with Fos family members (c-Fos, FosB, Fra-1 and Fra-2) or members of the ATF and MAF family (Zhao et al. 2014). Unlike the members of the Jun family, the Fos family proteins need to heterodimerize with members of the Jun family to form transcriptionally active complexes. After dimerization, AP-1 complexes bind to specific DNA consensus sequences as TRE (TPA responsive elements) or CRE (cAMP responsive elements) in the promoter and enhancer regions of several target genes (Angel and Karin 1991; Milde-Langosch et al. 2004). In vitro studies have shown that Jun–Fos heterodimers generate more stable complexes than Jun–Jun homodimers and therefore display a stronger DNA-binding activity and subsequent are more efficient in transcriptional control (Bamberger et al. 1999; Milde-Langosch et al. 2004). Accumulating evidence has implicated AP-1 in the regulation of a variety of cellular processes, including proliferation, differentiation, apoptosis, cell migration, invasion, transformation and adhesion (Hess et al. 2004; Milde-Langosch 2005; Shaulian 2010; Shaulian and Karin 2002; Wagner and Eferl 2005).
In this study, we focused on the transcription factor Fos-related antigen-1 (Fra-1), which is, beside c-Fos, the best studied member of the Fos family and encoded by the fos-like-1 gene (fosl1). The c-fos and fra-1 genes are best characterized as immediate early genes, and the transcriptional activity of Fra-1 is regulated both transcriptionally (Casalino et al. 2003; Young and Colburn 2006; Young et al. 2002) and posttranslationally (Smith et al. 1999). The expression of Fra-1 is deregulated in many tumors (Milde-Langosch 2005): its overexpression has been reported in proliferative disorders such as breast, brain, lung, colon, esophageal and thyroid cancer (Belguise et al. 2005; Chiappetta et al. 2000, 2007; Debinski and Gibo 2005; Kustikova et al. 1998; Logullo et al. 2011; Nakajima et al. 2007; Song et al. 2006; Usui et al. 2012; Young and Colburn 2006; Zajchowski et al. 2001). In addition, Fra-1 expression has been shown as a feature of hyperplastic and neoplastic breast epithelium (Chiappetta et al. 2007; Nakajima et al. 2007; Song et al. 2006) with capacity to influence proliferation, migration and invasiveness of breast cancer cells (Belguise et al. 2005; Kustikova et al. 1998). These findings implicate that Fra-1 activity might be functionally involved in breast cancer metastasis. In an immunohistochemical study, Logullo et al. (2011) found a positive correlation between Fra-1 expression and an aggressive phenotype in invasive breast ductal carcinoma.
Since tumor cell adhesion to endothelial cells has been shown to be crucial for hematogenic metastasis of various tumor types, we were interested in the potential influence of Fra-1 on the adhesive properties of breast cancer cells. Therefore, we performed dynamic adhesion assays using two breast cancer cell lines [ER(−) and ER(+)] with stable Fra-1 overexpression. Our results point to a strong influence of Fra-1 on cell adhesion to endothelia in ER-positive tumor cells and provide an explanation for the unfavorable prognostic impact of this protein in this molecular breast cancer subtype.
Materials and methods
Breast cancer cell lines and generation of stable transfectants
MDA MB231 and MCF7 breast cancer cells were cultivated as described before (Bamberger et al. 2001; Milde-Langosch et al. 2001, 2004). MDA MB231 cells express Fra-1 inherently high, while MCF7 cells do not show any detectable Fra-1 protein expression without transfection. To generate stable clones with increased Fra-1 expression, the full Fra-1 cDNA that had been cloned before in pcDNA3.1+ plasmids was supplemented with a C-terminal FLAG epitope by PCR and subcloned in the bicistronic vector pIRES-P (Genbank no. Z75185) using the NheI and EcoR1 restriction enzymes (Nandy et al. 2003). Transfection of MCF7 and MDA MB231 cells with this plasmid and the empty vector was performed with LipofectAMINE PLUS reagent (Life Technologies, Karlsruhe, Germany). After selection with puromycin (1 µg/ml), a single-cell Fra-1 clone (MCF7 Fra-1#8 and MDA MB231 Fra-1#2) and negative control clones (MCF7 NC and MDA MB231 NC) were chosen for further analysis. The differences in Fra-1 expression were repeatedly confirmed by Western blots during the experiments.
Cultured endothelial cells
Human pulmonary microvascular endothelial cells (HPMEC, Promocell, Heidelberg, Germany) were cultured in endothelial cell growth medium MV (PromoCell), supplemented with Supplement Mix (PromoCell) in vitro under standard culture conditions (37 °C, 100 % relative humidity, 5 % CO2). For the cell flow assay, HPMEC cells from passage one to twelve were stimulated with 10 ng/ml TNFα for 4 h.
Western blots
Western blot analyses were used to confirm Fra-1 overexpression in both cell lines as described (Milde-Langosch et al. 2004). Total protein extraction was performed using RIPA lysis buffer. For protein detection, the following antibodies were used: rabbit anti-Fra-1 antibody R20 (1:200; Santa Cruz Biotechnologies, Heidelberg, Germany) and goat anti-actin I19 (1:4000, Santa Cruz).
Cell flow assay
To study the influence of Fra-1 on adhesion of breast cancer cells to the endothelium during extravasation, we performed cell flow assays as described before (Dippel et al. 2013; Richter et al. 2011).
Flow cytometry
For assessment of certain adhesion molecules or selectin glycoligand structures on the surface of MCF7 and MDA MB231 transfectants, tumor cells were incubated with the following conjugated antibodies: anti-integrin α5 AF488 clone eBioSAM-1 (eBioscience, Frankfurt, Germany), anti-CD44v6 FITC clone VFF-7 (Invitrogen, Karlsruhe, Germany) and anti-CD 66c APC (R&D Systems, Minneapolis, MN 55413). Respective isotype controls, IgG1 AF488 (eBioscience), IgG2a APC (Dako, Hamburg, Germany) and IgG1 FITC (Miltenyi Biotec, Bergisch-Gladbach, Germany), were used. The stained cells were analyzed in the FACS Calibur (Becton/Dickinson, Heidelberg, Germany) or the CUBE 8 (Partec, Muenster, Germany) and visualized by FCS Express.
Patients cohort
We analyzed the mRNA expression data obtained from primary breast cancer tissue samples from our hospital (n = 194). Patients were treated at the University Medical Center Hamburg-Eppendorf, Germany, Department of Gynecology between 1991 and 2002 and selected on the basis of tissue availability. One hundred six patients received anthracycline-based adjuvant chemotherapy regimens [mainly epirubicin/cyclophosphamide (EC) or cyclophosphamide/methotrexate/fluorouracil (CMF)]. Seventy-four patients received endocrine therapy only, eight patients were treated by radiation without any systemic therapy and four patients remained untreated after surgery (no information: two patients). The median follow-up time was 132 months. Informed consent for the scientific use of tissue materials, which was approved by the local ethics committees (HH 05/04/2004), was obtained from all patients. The study was performed in accordance to the principles of the Declaration of Helsinki and REMARK criteria (Altman et al. 2012). No radiotherapy, neoadjuvant chemotherapy or endocrine therapy had been administered before surgery. Adjuvant therapy was applied according to international recommendations.
RNA isolation and microarray analysis
The Affymetrix (Santa Clara, CA, USA) HG-U133A array and GeneChip System™ were used to quantify the relative transcript abundance in the breast cancer tissues (n = 197) as described (Ihnen et al. 2008). Starting from 5 µg total RNA, labeled cRNA was prepared using the Roche Microarray cDNA Synthesis, Microarray RNA Target synthesis (T7) and Microarray Target Purification Kit, according to the manufacturer’s instructions. In the analysis settings, the global scaling procedure was chosen, which multiplied the output signal intensities of each array to a mean target intensity of 500. Samples with suboptimal average signal intensities (i.e., scaling factors >25) or GAPDH 3′/5′ ratios >5 were relabeled and rehybridized on new arrays.
Total RNA from MCF7 cells which were cultured to 70 % confluence was extracted by lysing the cells in Trizol reagent (Invitrogen) according to the manufacturer’s instructions and further purified using the RNeasy Mini Kit (Qiagen, Hilden, Germany). Procedures for cDNA synthesis, labeling and hybridization were carried out according to the manufacturer’s protocol (Affymetrix). The experiments were performed using Affymetrix Human Genome GeneChip U133 Plus 2.0 as described (Schroder et al. 2010). To compare samples and experiments, the trimmed mean signal of each array was scaled to a target intensity of 100. Absolute and comparison analyses were performed with Affymetrix GCOS (version 1.4, Affymetrix) software using default parameters. To assist in the identification of genes that were positively or negatively regulated in the experiment, we selected genes that were increased or decreased at least 1.9-fold compared to the control.
Statistical analysis
For evaluation of the prognostic significance of Fra-1 mRNA expression in breast cancer patients (n = 197), the cohort was first divided into three groups of similar size (n = 65–66) according to their expression values representing weak, moderate and high expression, and Kaplan–Meier analyses and log-rank tests were performed. Since cases with weak or moderate expression behaved similarly in survival analysis (Fig. 1a), both groups were combined for further stratified analyses in ER-positive and ER-negative groups. Associations of Fra-1 expression with clinical and histological variables were calculated by Chi-square tests using the following groups: nodal involvement (positive vs. negative), tumor stage (T1, T2, T3, T4), histological grading (G1/G2 vs. G3), estrogen receptor (ER) status and progesterone receptor (PR) status (positive vs. negative), histological type (ductal vs. lobular), differences in adherent cell numbers were calculated by two-sided T tests. All analyses were performed using the SPSS 21 software, and p values of <0.05 were considered to indicate a significant result.

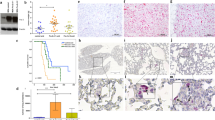
Prognostic impact of Fra-1 expression in breast cancer patients. Kaplan–Meier curves showing the prognostic value of Fra-1 in a 197 patients, b 148 ER(+) patients and c 41 ER(−) patients included in our collective. Broken line low and moderate Fra-1 expression; solid line high Fra-1 expression
Results
Fra-1 mRNA expression significantly correlates with shorter overall survival and lung metastasis in ER-positive breast cancer patients
Fra-1 mRNA expression was evaluated in 197 tumor samples of breast cancer patients using microarray data. Characteristics of patients included in this collective are shown in Table 1. In general, mRNA expression data for the Affymetrix probeset 204420_at (fosl1) in the tumors were low, ranging from 6.5 to 558.8 (mean 78.5 and median 62.2). According to these values, the cohort was first divided into three groups of similar size displaying low, moderate or high Fra-1 expression. High Fra-1 expression was associated with significantly shorter overall survival by Kaplan–Meier analysis (p = 0.027), and the same trend could be observed for progression-free survival (p = 0.227). Groups containing patients with low and moderate Fra-1 expression levels showed similar overall and progression-free survival times and were combined in a unique group for further analysis. As expected, Fra-1 retained its negative prognostic significance using these two groups as shown in Fig. 1a (OAS: p = 0.008; DFS: p = 0.085).
Since Fra-1 expression has been found to be mainly overexpressed in carcinomas with high metastatic potential such as in receptor-negative breast cancer, we were interested in the prognostic value of this transcription factor within the subgroups of ER-positive and ER-negative patients. Interestingly, high Fra-1 expression significantly correlates with shorter overall survival in ER-positive breast cancer patients (n = 150; p = 0.023; DFS: p = 0.149), but has no impact on the prognosis of patients with ER-negative tumors (n = 41; Fig. 1b–c). By Chi-square tests, we found significant associations of high Fra-1 expression levels with nodal involvement (p = 0.019) and with ER- and PR-negative status (p = 0.006 and p = 0.040), whereas no correlation with age, histological type, grading or stage could be found (Table 1). We further investigated whether the expression of Fra-1 correlated with distant metastasis in general or with metastasis to specific organ sites during recurrence. Thus, we found no association of Fra-1 expression with distant metastasis in general, but a significant correlation with metastasis to the lung (p = 0.021) and a weak association of high Fra-1 expression with brain metastasis (p = 0.056; Table 2).
Fra-1 regulates tumor cell adhesion to E-selectin and endothelial cells
Tumor cell interactions with the vascular endothelium are crucial for cell extravasation and therefore play an essential role in tumor progression and metastasis. Particularly, E-selectin, which is expressed in activated endothelial cells, has been implicated in metastatic spread in different tumor entities (Barthel et al. 2007). In order to evaluate the influence of Fra-1 on breast cancer cell adhesion, we established stable transfectants with forced Fra-1 overexpression in two human breast cancer cell lines: MCF7 (weakly invasive, ER-positive) and MDA MB231 (strongly invasive, ER-negative). High Fra-1 expression levels compared to the negative control cells (NC), which were generated after transfection with the empty vector, could be confirmed in monoclonal cell populations of either MCF7 (Fra-1#8) or MDA MB231 (Fra-1#2) cells by Western blot analysis (Fig. 2a).

Dynamic adhesion assays with Fra-1-overexpressing MCF7 and MDA MB231 cells. a Western blot analysis showing Fra-1 expression in a stably transfected clone derived from either MCF7 or MDA MB231 cells compared with untransfected cells and cells transfected with empty vector (NC). b Percent of adherent or rolling cells on E-selectin (upper side) and HPMECs (lower side) relative to control ± SEM are shown. *p < 0.05, **p < 0.005
We further analyzed the effect of Fra-1 overexpression on tumor cell adhesion to endothelial cells and to E-selectin under dynamic conditions. Using a laminar flow chamber assay with E-selectin- or HPMEC-coated surfaces, firm adhesion events as well as rolling behavior were evaluated. Cell adhesion or tethering was not observed when capillaries were coated with fetal calf serum, used as negative control for each experiment (data no shown). Interestingly, we observed a different adhesive behavior of MCF7 and MDA MB231 cells in these experiments: MCF7 exhibited firm, irreversible adhesion to endothelial monolayers or E-selectin-coated surfaces, whereas MDA MB231 cells rather demonstrated rolling behavior.
According to current knowledge, tumor cell interactions with endothelial E-selectin consist of a transient binding that facilitates cell tethering and rolling. Once tumor cells slowed down, firm adherence to different adhesion molecules of the endothelium and extravasation take place. In order to mimic the physiological situation, we measured adhesive events of tumor cells on E-selectin-coated capillaries at a shear stress rate of 0.5 dyn/cm2 and firm adhesion to endothelial cells at a lower rate of 0.25 dyn/cm2.
We observed significantly more adhesion events on E-selectin- and HPMEC-coated surfaces for Fra-1-overexpressing MCF7 cells (Fig. 2b; upper left: p = 0.012; lower left: p = 0.002) compared to NC cells, whereas the influence of Fra-1 on MDA MB231 tethering on E-selectin- (p = 0.067) or endothelial-coated surfaces (p = 0.557) was less prominent (Fig. 2b, right).
Results of microarray analysis and validation of candidate genes using flow cytometry
Since our analysis showed a prognostic impact of Fra-1 only for ER-positive breast cancer patients, and in vitro dynamic adhesion assays corroborated a strong influence of Fra-1 on the adhesive behavior of MCF7 (ER+) but not in MDA MB231 cells (ER−), we have chosen the breast cancer cell line MCF7 (MCF7Fra−1 versus MCF7NC) for further microarray analysis. In order to identify Fra-1 target genes that could be involved in metastatic properties of breast cancer cells, cDNA arrays were performed with mRNA isolated from the stably transfected MCF7 Fra-1# 8 and control transfectants (MCF7 NC). For evaluation of gene expression data, a threshold expression value of 50 and a signal log ratio (SLR) of ≥0.9 or ≤−0.9 compared to the control were used. Using these criteria, 1,041 probesets (664 genes) with known function (i.e., signal transduction, immune response, proteolysis or metabolism) were up-regulated, and 535 genes (390 probesets) were down-regulated in MCF7 pIRES Fra-1 cells. Our special attention in this study prevailed in genes, which might influence the adhesive properties of the Fra-1-overexpressing cells (selected genes are shown in Table 3).
Interestingly, overexpression of Fra-1 in MCF7 cells results in up-regulation of several adhesion molecules, notably CD44, CEACAM6 and integrin α5. In addition, Fra-1 overexpression correlates with higher levels of some components of the ECM, i.e., fibronectin (FN1), radixin (RDX) and different collagen isoforms. Furthermore, we found a couple of up-regulated genes participating in O- and N-glycosylation processes, namely UDP-GlcNac:betaGal beta-1,3-N-acetylglucosaminyltransferase3 (B3GNT3), heparan sulfate 6-O-sulfotransferase2 (HS6ST2) and ST6alpha-N-acetyl-neuraminyl-2,3-beta-galactosyl-1,3-N-acetylgalactosaminide-alpha-2,6-sialyl-transferase (ST6GALNAC4). On the other hand, diverse cell adhesion molecules or glycosylation enzymes, such as L1CAM, claudin 1 or NCAM1/2, FUCA1 and MAN1A1 (Table 3), were found to be down-regulated in Fra-1-overexpressing MCF7 cells in comparison with MCF7 NC cells.
The regulation of some Fra-1 target genes was validated on protein level by FACS analysis. In comparison with the original cell line MCF7 and the control transfectants MCF7 NC, we could show an obvious up-regulation of integrin α5 [arith. mean 37.55 (NC) to 131.35 (Fra-1#8)] and CD44v6 [arith. mean 34.10 (NC) to 144.59 (Fra-1#8)] on the cell surface of stably transfected MCF7 Fra-1#8 (Fig. 3) and a weak increase in CEACAM6 (arith. mean 9.01) in MCF7 Fra-1 cells compared to control MCF7 cells (arith. mean 5.0) (data not shown). The expression level of further cell adhesion-related proteins such as integrins α1, α2, α4, α6, αV, β1, β3, β4 and β7, ICAM, ALCAM, PSGL-1 and L1CAM was analyzed by FACS. Here, we did not detect deregulation of the mentioned adhesion proteins on MCF7 or MDA MB231 Fra-1-overexpressing cells in comparison with their respective controls.

FACS analysis. Results of flow cytometry showing increased ITGA5 and CD44v6 binding in MCF7 Fra-1#8 cells in comparison with MCF7 and MCF7 NC control cells. Black curves displayed isotype control, and gray curves show specific mAb binding, respectively. Each experiment repeated three times, and histograms of one representative experiment are displayed
Discussion
Several studies have described the effect of increased Fra-1 expression in several tumor entities (Chiappetta et al. 2000, 2007; Debinski and Gibo 2005; Kustikova et al. 1998; Logullo et al. 2011; Nakajima et al. 2007; Song et al. 2006; Usui et al. 2012; Zajchowski et al. 2001). In breast cancer cell lines, overexpression of this transcription factor leads to morphological changes, increased motility and invasive behavior in vitro (Belguise et al. 2005; Milde-Langosch 2005; Milde-Langosch et al. 2004). Here, we described for the first time the effect of Fra-1 expression on the adhesive properties of breast cancer cells in vitro and further showed, using microarray data from 194 breast cancer tumor samples, the prognostic relevance of this factor, particularly in the subgroup of ER-positive breast cancer patients.
The association between Fra-1 expression and a more malignant and aggressive phenotype in breast cancer has been evidenced by a variety of descriptive and functional studies (Belguise et al. 2005; Milde-Langosch et al. 2004; Zajchowski et al. 2001), whereas only few reports on the prognostic role of Fra-1 in this entity are known (Logullo et al. 2011). Logullo et al. (2011) investigated Fra-1 expression by immunohistochemistry in ductal carcinomas in situ (DCIS) and invasive ductal carcinomas (IDC) and could corroborate a clear correlation between Fra-1 expression and a more aggressive phenotype in IDC, but no association with overall survival. Our results, based on microarray data of 197 breast tumor samples, showed in contrast to a significant association of high Fra-1 mRNA expression with shorter patient outcome. Interestingly, when analyzing the ER-positive and ER-negative populations separately, we found that the prognostic impact of Fra-1 is restricted to the ER(+) population. This was an unexpected result, since the biological relevance of Fra-1 has been demonstrated in vitro for ER(+) as well as for ER(−) breast cancer cells, and enhanced proliferation rates, invasivity and motility correlate with high Fra-1 expression levels regardless of ER status (Belguise et al. 2005). Our statistical data, however, suggest a different role for Fra-1 in ER(+) versus ER(−) tumors. In line with our observations, Philips et al. demonstrated an inverse Fra-1-dependent regulation of AP-1 activity by estradiol in ER(+) versus ER(−) breast cancer cell lines and proposed that expression of AP-1-controlled genes might differ in ER(+) and ER(−) breast cancer cells (Philips et al. 1998).
The implication of AP-1 proteins in carcinogenesis has been described shortly after their discovery (Lamph et al. 1988). AP-1-regulated genes include important regulators of invasion, proliferation, differentiation, survival and genes associated with hypoxia and angiogenesis (Milde-Langosch 2005). Moreover, we have recently shown the effect of two AP-1 components, Fra-2 and c-Fos, on the adhesive properties and the metastatic potential of breast and ovarian cancer cells, respectively (Schroder et al. 2010; Oliveira-Ferrer et al. 2014). Regarding the functional role of Fra-1 in breast cancer, there are several experimental studies indicating an association between enhanced Fra-1 expression and proliferation, invasion and motility (Belguise et al. 2005). In addition, Fra-1 has been suggested to participate in the molecular switch during epithelial–mesenchymal transition (EMT) (Stinson et al. 2011). Nevertheless, nothing is known about Fra-1-mediated regulation of breast cancer cell adhesion features. Tumor cell adhesion and particularly interactions with the endothelium are critical for tissue colonization and metastasis development. The attachment of cancer cells to the endothelial layer is the first step of the extravasation process and requires the expression of adequate cell adhesion molecules. Mimicking the in vivo situation, we analyzed the effect of Fra-1 on the adhesive behavior of two breast cancer cell lines (MCF7 and MDA MB231) under dynamic conditions. MCF7 showed strong adhesion to endothelial monolayers or E-selectin-coated surfaces, whereas MDA MB231 cells rather demonstrated rolling behavior. By coating the flow channel surface with E-selectin or endothelial cells, we were able to discriminate between two different steps in the adhesion cascade: initial rolling or tethering events mediated by selectins (Tremblay et al. 2008) and firm cell adhesion on stimulated endothelial cells. Fra-1 led to a significant increase in MCF7 cell adhesion events on E-selectin as well as on stimulated endothelial cells under these conditions, whereas the effect on MDA MB231 rolling on both surfaces was less prominent and not significant. Since these two cell lines differ in their ER status [MCF7 is ER(+) and MDA MB231 is ER(−)], one could postulate, in line with our previous results from patient tumor samples, an ER-dependent influence of Fra-1 on the tumor cell adhesion features.
Our present data show that E-selectin binding is significantly increased in Fra-1-overexpressing MCF7 cells under flow conditions. Although the microarray data revealed up-regulation of some enzymes involved in O-glycosylation and the biosynthesis of Lewis antigens (B3GNT3 and B4GALT1), others such as FUT1 and FUT9 (Potapenko et al. 2010) were down-regulated in MCF7Fra−1. In fact, we could not detect higher expression levels of the minimal selectin-binding epitopes sialyl LewisX (CD15 s) and sialyl LewisA (CA19-9) by FACS analysis (data not shown), indicating that for MCF7 cells alternative E-selectin ligands have to be still postulated.
Enhanced adhesion of MCF7 cells to endothelial cells after Fra-1 overexpression might not only be explained to increased binding to E-selectin but also to other receptors contributing to stable cancer cell adhesion to ECs, i.e., integrins, CD44 and MUC1 (Miles et al. 2008; Kobayashi et al. 2007; Petruzzelli et al. 1999). In MCF7Fra−1 cells, up-regulation of integrin alpha5 and CD44 was detected at mRNA as well as at protein level using microarray and FACS analysis, respectively. This is particularly interesting since integrins follow selectins in the adhesion cascade of leukocytes, and cancer cells follow similar adhesion mechanisms (Strell and Entschladen 2008). CD44 expression on cancer cells strongly correlates with tumor cell adhesion to endothelial cells and metastasis particularly in prostate cancer and breast cancer cells (Draffin et al. 2004; Orian-Rousseau 2010). Interestingly, specific glycosylated forms of CD44 can interact with selectins (Zen et al. 2008). In addition, we detected higher mRNA expression levels of the cell adhesion molecule CEACAM6, which has been previously described to modulate tumor cell adhesion to endothelial cells (Blumenthal et al. 2005).
In contrast to these findings on MCF7 cells, in Fra-1-overexpressing MDA MB231 cells, we could not detect deregulation neither of those CAMs mentioned before (CD44, integrin alpha5 and CEACAM6) nor of other classical mediators of tumor cell adhesion such as integrins alpha 1/2/4/6 and V, integrins beta 1/3/4/7, ICAM-1, ALCAM, PSGL-1 or L1CAM. This underlines the hypothesis of a different regulative function of Fra-1 in ER(+) and ER(−) breast cancer cells.
Conclusion
In conclusion, our data show the prognostic relevance of the transcription factor Fra-1 for patients with ER(+) breast cancer and suggest different roles of Fra-1 in breast tumor cells depending on the ER status. In addition to its known pro-proliferative and pro-invasive effect, Fra-1 might influence breast cancer progression by modulating the adhesion of tumor cells to endothelial surfaces. Single Fra-1 deregulated genes and their relevance in mediating cell adhesion to endothelial cells should be further analyzed in order to identify new target molecules for therapeutic purposes.
References
Altman DG, McShane LM, Sauerbrei W, Taube SE (2012) Reporting recommendations for tumor marker prognostic studies (REMARK): explanation and elaboration. BMC Med 10:51. doi:10.1186/1741-7015-10-51
Angel P, Karin M (1991) The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim Biophys Acta 1072:129–157
Bamberger AM, Methner C, Lisboa BW, Stadtler C, Schulte HM, Loning T, Milde-Langosch K (1999) Expression pattern of the AP-1 family in breast cancer: association of fosB expression with a well-differentiated, receptor-positive tumor phenotype. Int J Cancer 84:533–538. doi:10.1002/(SICI)1097-0215(19991022)84:5<533:AID-IJC16>3.0.CO;2-J
Bamberger AM, Milde-Langosch K, Rossing E, Goemann C, Loning T (2001) Expression pattern of the AP-1 family in endometrial cancer: correlations with cell cycle regulators. J Cancer Res Clin Oncol 127:545–550
Barthel SR, Gavino JD, Descheny L, Dimitroff CJ (2007) Targeting selectins and selectin ligands in inflammation and cancer. Expert Opin Ther Targets 11:1473–1491. doi:10.1517/14728222.11.11.1473
Belguise K, Kersual N, Galtier F, Chalbos D (2005) FRA-1 expression level regulates proliferation and invasiveness of breast cancer cells. Oncogene 24:1434–1444. doi:10.1038/sj.onc.1208312
Blumenthal RD, Hansen HJ, Goldenberg DM (2005) Inhibition of adhesion, invasion, and metastasis by antibodies targeting CEACAM6 (NCA-90) and CEACAM5 (Carcinoembryonic Antigen). Cancer Res 65:8809–8817. doi:10.1158/0008-5472.CAN-05-0420
Casalino L, De Cesare D, Verde P (2003) Accumulation of Fra-1 in ras-transformed cells depends on both transcriptional autoregulation and MEK-dependent posttranslational stabilization. Mol Cell Biol 23:4401–4415
Chiappetta G et al (2000) FRA-1 expression in hyperplastic and neoplastic thyroid diseases. Clin Cancer Res 6:4300–4306
Chiappetta G et al (2007) FRA-1 protein overexpression is a feature of hyperplastic and neoplastic breast disorders. BMC Cancer 7:17. doi:10.1186/1471-2407-7-17
Debinski W, Gibo DM (2005) Fos-related antigen 1 modulates malignant features of glioma cells. Mol Cancer Res 3:237–249. doi:10.1158/1541-7786.MCR-05-0004
Dippel V et al (2013) Influence of L1-CAM expression of breast cancer cells on adhesion to endothelial cells. J Cancer Res Clin Oncol 139:107–121. doi:10.1007/s00432-012-1306-z
Draffin JE, McFarlane S, Hill A, Johnston PG, Waugh DJ (2004) CD44 potentiates the adherence of metastatic prostate and breast cancer cells to bone marrow endothelial cells. Cancer Res 64:5702–5711. doi:10.1158/0008-5472.CAN-04-0389
Hess J, Angel P, Schorpp-Kistner M (2004) AP-1 subunits: quarrel and harmony among siblings. J Cell Sci 117:5965–5973. doi:10.1242/jcs.01589
Ihnen M et al (2008) Predictive impact of activated leukocyte cell adhesion molecule (ALCAM/CD166) in breast cancer. Breast Cancer Res Treat 112:419–427. doi:10.1007/s10549-007-9879-y
Kobayashi H, Boelte KC, Lin PC (2007) Endothelial cell adhesion molecules and cancer progression. Curr Med Chem 14:377–386
Kustikova O, Kramerov D, Grigorian M, Berezin V, Bock E, Lukanidin E, Tulchinsky E (1998) Fra-1 induces morphological transformation and increases in vitro invasiveness and motility of epithelioid adenocarcinoma cells. Mol Cell Biol 18:7095–7105
Lamph WW, Wamsley P, Sassone-Corsi P, Verma IM (1988) Induction of proto-oncogene JUN/AP-1 by serum and TPA. Nature 334:629–631. doi:10.1038/334629a0
Logullo AF et al (2011) Role of Fos-related antigen 1 in the progression and prognosis of ductal breast carcinoma. Histopathology 58:617–625. doi:10.1111/j.1365-2559.2011.03785.x
Milde-Langosch K (2005) The Fos family of transcription factors and their role in tumourigenesis. Eur J Cancer 41:2449–2461. doi:10.1016/j.ejca.2005.08.008
Milde-Langosch K, Bamberger AM, Goemann C, Rossing E, Rieck G, Kelp B, Loning T (2001) Expression of cell-cycle regulatory proteins in endometrial carcinomas: correlations with hormone receptor status and clinicopathologic parameters. J Cancer Res Clin Oncol 127:537–544
Milde-Langosch K et al (2004) The role of the AP-1 transcription factors c-Fos, FosB, Fra-1 and Fra-2 in the invasion process of mammary carcinomas. Breast Cancer Res Treat 86:139–152. doi:10.1023/B:BREA.0000032982.49024.71
Miles FL, Pruitt FL, van Golen KL, Cooper CR (2008) Stepping out of the flow: capillary extravasation in cancer metastasis. Clin Exp Metastasis 25:305–324. doi:10.1007/s10585-007-9098-2
Nakajima H et al (2007) Aberrant expression of Fra-1 in estrogen receptor-negative breast cancers and suppression of their propagation in vivo by ascochlorin, an antibiotic that inhibits cellular activator protein-1 activity. J Antibiot (Tokyo) 60:682–689. doi:10.1038/ja.2007.87
Nandy A, Jenatschke S, Hartung B, Milde-Langosch K, Bamberger AM, Gellersen B (2003) Genomic structure and transcriptional regulation of the human NAD+-dependent 15-hydroxyprostaglandin dehydrogenase gene. J Mol Endocrinol 31:105–121
Oliveira-Ferrer L et al (2014) c-FOS suppresses ovarian cancer progression by changing adhesion. Br J Cancer 110:753–763. doi:10.1038/bjc.2013.774
Orian-Rousseau V (2010) CD44, a therapeutic target for metastasising tumours. Eur J Cancer 46:1271–1277. doi:10.1016/j.ejca.2010.02.024
Petruzzelli L, Takami M, Humes HD (1999) Structure and function of cell adhesion molecules. Am J Med 106:467–476
Philips A, Teyssier C, Galtier F, Rivier-Covas C, Rey JM, Rochefort H, Chalbos D (1998) FRA-1 expression level modulates regulation of activator protein-1 activity by estradiol in breast cancer cells. Mol Endocrinol 12:973–985
Potapenko IO et al (2010) Glycan gene expression signatures in normal and malignant breast tissue; possible role in diagnosis and progression. Mol Oncol 4:98–118. doi:10.1016/j.molonc.2009.12.001
Richter U et al (2011) Adhesion of small cell lung cancer cells to E- and P-selectin under physiological flow conditions: implications for metastasis formation. Histochem Cell Biol 135:499–512. doi:10.1007/s00418-011-0804-4
Schroder C et al (2010) The transcription factor Fra-2 promotes mammary tumour progression by changing the adhesive properties of breast cancer cells. Eur J Cancer 46:1650–1660. doi:10.1016/j.ejca.2010.02.008
Shaulian E (2010) AP-1–the Jun proteins: Oncogenes or tumor suppressors in disguise? Cell Signal 22:894–899. doi:10.1016/j.cellsig.2009.12.008
Shaulian E, Karin M (2002) AP-1 as a regulator of cell life and death. Nat Cell Biol 4:E131–136. doi:10.1038/ncb0502-e131
Smith LM et al (1999) cJun overexpression in MCF-7 breast cancer cells produces a tumorigenic, invasive and hormone resistant phenotype. Oncogene 18:6063–6070. doi:10.1038/sj.onc.1202989
Song Y et al (2006) An association of a simultaneous nuclear and cytoplasmic localization of Fra-1 with breast malignancy. BMC Cancer 6:298. doi:10.1186/1471-2407-6-298
Stinson S et al (2011) miR-221/222 targeting of trichorhinophalangeal 1 (TRPS1) promotes epithelial-to-mesenchymal transition in breast cancer. Sci Signal 4:pt5. doi:10.1126/scisignal.2002258
Strell C, Entschladen F (2008) Extravasation of leukocytes in comparison to tumor cells. Cell Commun Signal 6:10
Tremblay PL, Huot J, Auger FA (2008) Mechanisms by which E-selectin regulates diapedesis of colon cancer cells under flow conditions. Cancer Res 68:5167–5176. doi:10.1158/0008-5472.CAN-08-1229
Usui A et al (2012) The molecular role of Fra-1 and its prognostic significance in human esophageal squamous cell carcinoma. Cancer 118:3387–3396. doi:10.1002/cncr.26652
Wagner EF, Eferl R (2005) Fos/AP-1 proteins in bone and the immune system. Immunol Rev 208:126–140. doi:10.1111/j.0105-2896.2005.00332.x
Young MR, Colburn NH (2006) Fra-1 a target for cancer prevention or intervention. Gene 379:1–11. doi:10.1016/j.gene.2006.05.001
Young MR et al (2002) Transactivation of Fra-1 and consequent activation of AP-1 occur extracellular signal-regulated kinase dependently. Mol Cell Biol 22:587–598
Zajchowski DA et al (2001) Identification of gene expression profiles that predict the aggressive behavior of breast cancer cells. Cancer Res 61:5168–5178
Zen K et al (2008) CD44v4 is a major E-selectin ligand that mediates breast cancer cell transendothelial migration. PLoS One 3:e1826. doi:10.1371/journal.pone.0001826
Zhao C et al (2014) Genome-wide profiling of AP-1-regulated transcription provides insights into the invasiveness of triple-negative breast cancer. Cancer Res 74:3983–3994. doi:10.1158/0008-5472.CAN-13-3396
Acknowledgments
We are very grateful to the excellent technical assistance of Maila Rossberg and Kathrin Eylmann.
Conflict of interest
All authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
L. Oliveira-Ferrer and M. Kürschner have contributed equally to this work.
Rights and permissions
About this article

Cite this article
Oliveira-Ferrer, L., Kürschner, M., Labitzky, V. et al. Prognostic impact of transcription factor Fra-1 in ER-positive breast cancer: contribution to a metastatic phenotype through modulation of tumor cell adhesive properties. J Cancer Res Clin Oncol 141, 1715–1726 (2015). https://doi.org/10.1007/s00432-015-1925-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00432-015-1925-2