
Abstract
In a location-based negative priming paradigm, the possibility of a disengagement option of the underlying inhibitory mechanism was tested. Whereas in previous studies disengagement was observed when providing utility information about the probe trial structure, in the present study the allocation of visual attention to the stimuli was manipulated. In the first step an automatic deployment of visual attention was implemented by presenting all stimuli as abrupt onsets (Experiment 1), which demonstrated commonly observed negative priming effects. In further conditions of non-automatic allocation of visual attention in which target and distractor were presented as no-onset stimuli, negative priming effects were eliminated (Experiments 2 and 3). The preferred interpretation is that in conditions of automatic control of attention, target and distractor compete for control of action. A non-automatic control of visual attention, on the other hand, leads to a top-down modulated selection, which results in prioritized target encoding and a loss of distractor impact on the selection process. Alternative accounts and the role of no-onset distractor processing were investigated in Experiment 4.
Similar content being viewed by others
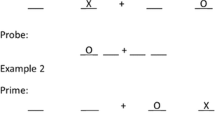

Avoid common mistakes on your manuscript.
Location-based negative priming
In our everyday interaction with the surrounding environment, we are constantly forced to select relevant information and to abandon irrelevant information in order to achieve an anticipated goal. The efficiency, the restrictions, and the flexibility of this selection process are main topics of research in cognitive psychology. Which processes are involved in realizing our intention to attend to relevant information and to neglect irrelevant information, e.g., to resist the strongest activated response at a certain moment (Tipper, 2001)? For the longest time the solution to this problem has been seen in the effect of inhibitory mechanisms. When one must respond to a certain target location in the presence of a distractor in a different location, both stimuli compete for control of action. Responding to the location of the target could involve the active inhibition of the distractor itself, its response and/or its location. The plausible and useful function of inhibition in such a selection task can be seen to prevent stimuli in irrelevant locations to gain control of action (Tipper, 1985; Tipper & Cranston, 1985).
However, our world is not static. This means that an irrelevant location in one situation (prime trial) might suddenly become relevant in the next situation (probe trial). When a target appears in a location that has previously been occupied by a distractor (so called ignored repetition), response times (RTs) are generally slowed down, compared with situations in which the target location has been empty in the preceding trial (control condition). In terms of an inhibition-based explanation, inhibitory processes bound to the distractor (location) of the prime trial are still in effect when the upcoming target appears in the same location and thus hampers performance.
The example just described is widely known as the negative priming procedure, which is one method of studying the nature of inhibitory mechanisms in selection tasks. The slowing of RTs in such an ignored repetition trial has been shown to generalize to a wide variety of materials and tasks (for overviews see Fox, 1995; Neill, Valdes, & Terry, 1995; Tipper, 2001). Most results refer to negative priming with location or other attributes serve as selection cues and identity (e.g., color and shape) as the dimension to be reported; so called identity-negative priming. Besides inhibition as a key mechanism for negative priming, several other theoretical frameworks have been proposed to account for identity-negative priming: Memory retrieval accounts (Neill & Valdes, 1992; Neill, Valdes, Terry, & Gorfein, 1992), temporal discrimination accounts (Milliken, Joordens, Merikle, & Seiffert, 1998), and integrated accounts (Tipper, 2001).
However, our example describes the location-based negative priming paradigm in which identity or other attributes function as selection cue and location represents the dimension to be reported. This distinction is quite important because there is an increasing amount of work that has demonstrated a dissociation of identity-based and location-based negative priming effects in similar task conditions. Kane, May, Hasher, Rahal, and Stoltzfus (1997), for example, showed that location-based negative priming is still intact in the elderly, whereas identity negative priming is impaired. It is now widely assumed that the mechanisms of identity and location negative priming are most likely not identical (Neill, Terry, & Valdes, 1994; Milliken, Tipper, Houghton, & Lupianez, 2000). Therefore, for the purpose of this study, when talking about negative priming we generally refer to location-based negative priming.
The impairment of behavior when the relevant probe target location is subject to the consequences of prime distractor inhibition clearly illustrates that this originally useful selection process can have detrimental effects in the current situation. In other words, the automatic inhibitory mechanism can interfere with goal-directed processing when its original situation has changed (Buckolz, Boulougouris, O‘Donnell, & Pratt, 2002a). Buckolz et al. conclude that it would be most useful for the efficiency of the cognitive system in complex behavior if selection processes were equipped with the option of disengagement, making the selection process in target localization tasks adaptive and flexible.
If this idea proves to be correct, then based on the adaptive nature of the inhibitory process, one should be able to demonstrate its dependence on task conditions. Indeed, Buckolz et al. (2002a) were able to disengage the location-based priming mechanism by providing participants with utility information about the upcoming target location. Only in conditions including visual cues with 100% cue-target validity (Experiment 2) predicting the location of the probe target, participants were able to translate this utility information into the disengagement of the negative priming mechanism. Thus, the consequences of prime distractor inhibition did not affect probe target processing and therefore, negative priming effects were significantly reduced.
Another study demonstrated that the inhibitory mechanism of location-based negative priming can be disengaged when participants know about the predictable absence of probe trial distractors. Buckolz, Boulougouris, and Khan (2002b) presented 100% reliable cues before or after the prime trial as to whether a distractor would or would not accompany the target in the probe trial. The certainty of the distractor absence in the probe trial led participants to adapt a most useful processing strategy in dropping the “selection state” (the expectation of target selection) and thus disengage the inhibitory mechanism that was at work in the prime trial. The conclusion that can be drawn from these studies is that the cognitive system possesses a sophisticated control mechanism that is able to make use of utility information and to disengage the automatic inhibitory mechanism in order to enhance future probe target processing.
The central question of our study is whether the disengagement of the automatic inhibitory mechanism in a selection task is bound to utility information as shown in the studies by Buckolz et al. (2002a, 2002b), or whether other processing components also disengage the inhibitory mechanism in a selection task. Our approach is to manipulate the deployment of visual attention toward the stimuli in a location-based negative priming paradigm. The allocation of visual attention is supposed to affect the encoding conditions of stimuli, biases the processing priority of a stimulus, and thus affects the selection process in general. We assume that the manipulation of the deployment of visual attention within a selection task such as negative priming could provide another possibility of demonstrating the flexible and adaptive nature of the underlying inhibitory mechanism.
Of course, there is already evidence that directing attention modulates priming effects. Recently, Stablum, Ricci, Pavgese, and Umiltá (2001) explored the role of attention in identity negative priming. The authors started from the evidence that directing attention to the local level in local/global processing of Navon letters requires a higher degree of selectivity than directing attention to the global level. Consequently, they found greater negative priming effects for the local than for the global attention situation. Yee, Santoro, Grey, and Woog (2000) investigated whether there is a connection between identity negative priming effects and processes that direct attention to target stimuli. They manipulated the level of processing (conceptual versus perceptual) at target selection. Negative priming was greater when conceptual processing was required at target selection. MacDonald, Joordens, and Seergobin (1999) also examined the influence of attention manipulation on negative priming. They used referent-size selection tasks in which participants were instructed to name the larger of two animals. The level of processing induced by this task enhanced the negative priming effects as well. Although these results show that there are connections between negative priming and processes that direct attention to target stimuli, the evidence refers to negative priming with location (or some other attributes) as selection cue and identity as the dimension to be reported (identity negative priming) and not to location-based negative priming. So far, referring to the inhibition account of identity negative priming one could argue that the inhibition system can adapt to the demands of the task (Richards, 1999).
The control of visual attention
Why would we expect that differences in the control of visual attention would affect the disengagement of the negative priming mechanism? Before we address this question, we will briefly introduce the concept of the control of visual attention.
The control of attention within a visual field can be viewed as a function of the goals of the observer and of image properties—a distinction that is widely known as goal-driven and stimulus-driven control of attention. One approach to studying the effects of control and deployment of visual attention in this context is the manipulation of the presentation mode of stimuli. If for example an object or a stimulus appears abruptly in the visual field, this is called abrupt onset stimulus (Yantis & Jonides, 1984). The sudden onset of stimuli captures processing resources and interferes with the processing of other stimuli. The uniqueness and the specific characteristics of abrupt onset stimuli have been demonstrated in many studies (Jonides & Yantis, 1988; Todd & Van Gelder, 1979; Yantis & Jonides, 1984, 1990).
Yantis and Jonides (1984; Jonides & Yantis, 1988), for instance, conducted a visual search task in which they compared an abrupt onset presentation with a no-onset presentation. In a no-onset procedure, an item does not appear abruptly in the visual field; instead it is covered by a mask from the beginning of its presentation on the screen and, only after a certain amount of time, the actual item is revealed by removing the mask. The authors’ results showed that in a singleton search task the identification of the target was always enhanced when the abrupt onset stimulus represented the target letter. On the other hand, when one of the no-onset stimuli revealed the target, then identification time depended heavily on display size, which was interpreted in terms of an attention demanding, serial search (Yantis & Jonides, 1990). The authors argued that participants generally identify abrupt onset stimuli first. The abrupt onset of the stimulus in a previously blank space automatically captures visual attention, regardless of any other no-onset stimuli in the display. A serial, self-terminating search for the no-onset letters takes place, only after the onset item is identified. It was also shown that the specifics of onset stimuli are not due to luminance changes on the screen (Yantis & Hillstrom, 1994), which means that the control of attention must occur at the level of objects rather than the level of luminance differences. Thus, for onset stimuli it can be assumed that stimulus controlled processing gains more weight in comparison to goal controlled processing.
That onset stimuli have higher priority can also be learned from the results of Experiment 2 by Fox (1994). Using a flanker paradigm, she primed the prime target location and found a greater negative priming effect. With control experiments it was then shown that the pre-cuing effect was produced by forward masking (cue presentation time of 150 ms, which fits the time scale of forward masking; DiLollo, 1980) of the target. The abrupt onset of the distractor in a previously empty spot may have captured attention to the distractor’s location. In this context, the stronger negative priming effects can be seen as an artefact of the particular pre-cuing manipulation. We interpret this result as showing that manipulation of priority setting (in this case by cues) influences the negative priming effect.
Pratt and McAuliffe (2001) compared single and simultaneous onset and offset cues on choice localization key press responses. They also found that onset cues have priority in orienting attention in localization tasks (see also Pratt & Hirshhorn, 2003). In a classical inhibition of return paradigm (IOR), Samuel and Weiner (2001) investigated the attentional consequences of object appearance and disappearance. Comparing onset stimuli and classical offset stimuli (detection of an object disappearance), they speculate that onset stimuli generate attentional consequences reflexively, whereas the attentional consequences of object disappearances may be more voluntary.
A plausible explanation of the differences between onset and no-onset items in their ability to capture visual attention was provided by Yantis (1998). He argued that the appearance of a new object (onset stimulus) requires the creation of an object representation, which can be seen as part of the encoding process of onset stimuli. The necessity of creating an object representation triggers an involuntary and automatic shift of attention to the new object (see also Yantis & Hillstrom, 1994). It is straightforward when he states: “Overall, the experiments we have carried out lead us to conclude that the appearance of a new perceptual object is an important perceptual event that has significant consequences for the deployment of attention...the visual system appears to be predisposed to attend to objects that require the creation of a new perceptual object representation” (Yantis, 1998, p. 251). The conclusion that can be drawn from these studies is that an abrupt onset presentation mode of stimuli results in an automatic capture of visual attention (bottom–up control), whereas the identification of no-onset stimuli represents a self-terminating serial search and therefore, voluntary deployment of visual attention (top–down control).
Location-based negative priming and the control of visual attention
In the present research, we used the distinction between onset versus no-onset presentation to implement different kinds of attentional control within a negative priming paradigm. We suspect that in a selection task such as negative priming the manipulation of the deployment of visual attention will have crucial consequences for the selection process itself.
There are two important questions: First, how does the manipulation of attentional control affect the selection process? Second, how does this result in a potential disengagement of the negative priming mechanism? We will now evaluate these questions in terms of an onset stimulus presentation and a no-onset stimulus presentation respectively.
Onset presentation of stimuli
In the context of the specific characteristics of a stimulus onset presentation, it is curious to note that almost all experiments dealing with negative priming have used an abrupt onset presentation of stimuli. Target as well as distractor always appeared together in an abrupt onset mode on the display and thus captured attention automatically (bottom–up control of visual attention). Since the necessity of creating a new object representation is an inherent characteristic of the onset presentation mode, involuntary attentional capture should apply for relevant and irrelevant onsets simultaneously. Such a view is in accordance with studies that demonstrate prioritized processing of multiple onset stimuli. The reported number of prioritized onsets varies from four (Yantis & Johnson, 1990; Yantis & Jones, 1991) to 14 items (Donk & Theeuwes, 2003). It is assumed that multiple abrupt onsets generate large bottom–up activation with the consequence that these stimuli receive involuntary attentional priority (Cave & Wolfe, 1990; Donk & Theeuwes, 2001, 2003).
In a negative priming paradigm, the generation of new object representations is required for both onset stimuli. Target and distractor capture attention simultaneously, which results in a competition for selection. Responding efficiently to the target location requires the inhibition of the distractor location, which in classical accounts is seen as the origin of negative priming effects observed in the subsequent trial (Houghton & Tipper, 1994; Tipper & Cranston, 1985). Even if one argues in favor of a random capture (Reder, Weber, Shang, & Nanuykov, 2003), e.g., half of the time the target location captures attention first and in the other 50% the location of the distractor receives attention first, the onset distractor would still reveal a strong impact on the selection process. Thus, it is not surprising that negative priming effects are commonly observed when presenting target and distractor simultaneously as abrupt onset stimuli.
No-onset presentation of stimuli
We argued that when presenting stimuli in a no-onset fashion these stimuli do not capture attention automatically. From offset studies we know that the identification of such stimuli reflects controlled and serial processes. In other words, it is not the stimuli themselves that lead to a capture of visual attention; but the expectation and the goal of the participant (attentional set) that allocate visual attention voluntarily to the target. We think that a no-onset presentation gives rise to a goal-directed object identification, which increases the efficiency of target selection. Such an assumption is in accordance with visual search studies that showed that providing a top–down setting participants can adopt a feature detection mode in which they are able to exclusively allocate attention to relevant target features (Bacon & Egeth, 1994). This is interpreted as a goal-directed selection of a specific known featural target identity in which irrelevant singletons are no longer supposed to interfere.
It is also known that participants have voluntary influence over which location will benefit from biased competition and thus which location will be selected (e.g., Jonides, 1981; Müller & Rabbitt, 1989). Support for our hypothesis that voluntary influences are not limited to the spatial distribution of attention, but can also bias the processing priority for certain stimuli, comes from a recent study by Awh, Matsukura, and Serences (2003). Manipulating the probability of distractor interference they demonstrated that top–down settings can lead to changes specific to distractor processing in terms of changes in biased competition during perceptual stages. They suggested: “...That changes in top–down settings can influence the degree to which competition is biased in favor of the attended stimuli” (p. 62). We think that in a selection task in which all stimuli are presented as no-onsets and attention is allocated voluntarily, the attentional set of the participant will potentially bias selection in terms of increased efficiency in target selection. In other words, a no-onset presentation of stimuli has the potential to implement a goal-directed selection in which target processing is facilitated.
If target selection is indeed facilitated by a voluntary allocation of visual attention to the stimuli, what enables participants to disengage the negative priming mechanism? One could assume that, when a top–down setting is provided that helps participants to efficiently select the target, the mere presence of a distractor becomes irrelevant within the selection process. If this is the case, one should not obtain any differences in target selection whether a distractor is present or absent. In an inhibition-based negative priming account, inhibition is seen as a mechanism that is needed to efficiently select the target by inhibiting the competing distractor. This distractor inhibition in the prime trial results in prolonged RTs when the probe target appears in the prime distractor location. Since we assume that when attention is voluntarily allocated to the stimuli, which results in a most efficient target selection and a strongly reduced distractor impact on this selection process, an inhibitory mechanism to reduce distractor impact might become unnecessary and can therefore be abandoned.
Taken one step further, prioritized target selection in prime and probe trial might even completely eliminate any demand for selection so that participants could drop the so-called “selection state” altogether. Such a top–down setting in which distractors do not automatically capture attention and may not reveal an influence on the selection process would provide a promising option for disengaging the inhibitory mechanism responsible for negative priming (see the General discussion section for an evaluation of whether such disengagement would happen intentionally or unintentionally).
The first experiment focused on the replication of negative priming effects in a classical location-based negative priming condition in which all stimuli were presented as abrupt onsets, representing an automatic or bottom–up control of visual attention. We expected significant negative priming effects. In Experiments 2 and 3, the automatic control of visual attention was altered to a non-automatic control, presenting stimuli in a no-onset mode for both prime and probe trials. We expected an efficient (goal-directed) target selection that eliminates the need for distractor inhibition. Participants may drop the “selection state” throughout the experiment, which provides a basis for the disengagement of the inhibitory mechanism of negative priming. A fourth experiment was conducted to investigate the processing or non-processing of no-onset distractors when visual attention is allocated voluntarily within the selection process (no-onset presentation). Presenting prime and probe trials in Experiments 4A and 4B in different presentation modes (onset vs. no-onset) will also be informative about the disengagement of the inhibitory mechanism when selection is required in at least one trial type (prime or probe trial).
General method (Experiments 1–3)
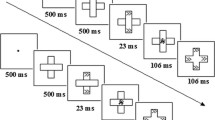
Participants completed three experiments in two sessions. They started in the first session with either Experiment 1 or 2. In the second session they completed Experiment 3. All experiments were based on a location negative priming paradigm similar to the one used by Tipper, Weaver, and Houghton (1994). The experimental design consisted of four black squares (side length 20 mm) in front of a grey background (see Fig. 1). In the squares, an “O” and an “X” or just a single “O” in black could appear in any position. Participants were seated approximately 900 mm from the monitor, which resulted in a visual angle of 2.1° (horizontally and vertically from the screen center) considering the size of the stimuli. Participants were instructed to respond as fast and as accurately as possible to the location of the “O” and to ignore the “X.” The keys “2,” “4,” “8,” and “6” corresponded to the location “bottom,” “left,” “top,” and “right” respectively. Responses had to be given with the right index finger, which rested at the location of the key “5” in the middle of the number block. RTs were measured from presentation of stimuli until pressing the actual location button. After response execution the index finger returned to the central key “5.” Resting the right index finger on the key “5” without pressing it ensured that the distance to each response key was kept constant. Therefore, measured RTs in this study consisted of movement times (from the central key to the required response key) and the actual button press time. After the given response in two consecutive trials (prime and probe trial), the word “next” appeared on the screen. Using the left hand for pressing the space bar, participants were able to control the beginning of the next trial combination on their own. A break split each experiment into two identical blocks. Each block contained four different conditions (see Fig. 2). In the ignored repetition (IR) condition the location of the prime-distractor becomes the location of the probe-target. There were no other repetitions of stimulus location between prime and probe trial. The C condition represents a control condition in which all stimuli change their location from prime to probe trial without repetition of any location. In both IR and control, a target and a distractor always appear in prime and in probe trials. Contrary to this, in the no-distractor (ND) condition a target appeared alone in prime as well as in probe trials. The probe-distractor (PD) condition, on the other hand, contained a distractor only in probe trials, but not in prime trials. Thus, these two conditions were the only ones in which a target could appear without a distractor. PD and ND were included for the purpose of interference control. With the exception of the IR condition, in which the probe target appeared in the prime distractor location, no other repetitions of location were possible.

Time characteristics of an ignored repetition (IR) trial combination in Experiment 1. The arrows represent the time and not a change of position on the display. All stimuli appear within the four consistent squares according to the time characteristics

Conditions included in all experiments. In the IR condition the location of the prime-distractor becomes the location of the probe-target. The C condition represents a control condition in which all stimuli change their location. The probe-distractor (PD) and no-distractor (ND) conditions were included for the purpose of interference control. With the exception of the IR condition there were no repetitions of positions.
A prime and a probe trial created individual trial combinations that were presented randomly and at the same frequency for each participant. Altogether, each experiment consisted of 192 prime-probe combinations. This breaks down to 48 prime-probe pairs for each condition (IR, C, PD, and ND respectively).
Negative priming effects were calculated by subtracting the probe C condition from the probe IR condition. Interference for probe trials was measured using probe PD and ND conditions (see Fig. 2). In order to measure interference effects for prime trials, the PD and ND conditions were taken together (both conditions did not contain a prime distractor) and were compared with the summarized IR and C conditions (both contained a prime distractor).
The design of our experiments does not include all possible conditions of prime-probe relations because repetition of location was only possible in the IR condition (probe target appears at prime distractor location). This means that not all stimuli appear equally often in all locations. This leads to an argument from Christie and Klein (2001) who state that, in such a design, prime trials contain information about probe target location. They argued that such an unbalanced design is insufficient to draw inferences from attended repetition effects (probe target appears in prime target location). In our Experiments (1–4), we do not have an attended repetition condition. Probe targets never appear in prime target locations. So what might then be the consequences for our research when prime trials do contain information about probe trials? In this study we included an equal amount of trials in the IR and C conditions. Thus, the probability of a probe target occurring in the prime distractor location is .5 (IR), which equals the probability of a probe target appearing in a previously empty location (C). The difference is that in an IR condition there is only one location, namely the prime distractor location, for the probe target to appear at. On the other hand, the C condition provides more than one control location for the probe target to appear. The advantage of specifying the location in IR conditions in contrast to C conditions could potentially improve the efficiency of code selection at the critical target location and thus facilitate responses to the target. In the worst case, this would mean a reduction in negative priming effects. That this is most likely not the case has been shown in a recent study by Reder et al. (2003, Experiment 1). Quite the opposite effect was observed. The authors found large negative priming effects when the probability of a target following a distractor at the same location was about 80%. Moreover, Buckolz et al. (2002a) have shown that only a cue validity of 100% indicating the upcoming target position is effective in significantly reducing negative priming. Therefore, because we do not have an attended repetition condition, probabilistic information conveyed by the prime trial about the probe trial is not a serious concern for our research.
Participants (Experiments 1–3)
Twenty-five undergraduate students (21 females) of the Department of Psychology at Humboldt University, Berlin, participated in all three experiments in exchange for course credits. Their ages ranged from 19 to 30 years (mean age 23.1 years). All students were right-handed and had normal or corrected-to-normal vision.
Apparatus
Stimuli were presented on a 15 inch color monitor that was connected to a 486 PC. Experiments were carried out using Experimental Run Time System (ERTS) software (Beringer, 2000).
Experiment 1: Negative priming and an automatic deployment of visual attention
The first experiment was conducted to replicate negative priming effects using a location-based negative priming paradigm, which includes a typical onset presentation mode of the stimuli representing an automatic deployment of visual attention. Target and distractor appeared abruptly on the display. The design of this experiment was based on an adaptation of the study by Tipper et al. (1994) as described above. In both studies, the relevant response dimension was the location of the target. Although the identity of the items was the basis of selection, it was irrelevant for the actual response. In terms of this experimental design and the described modalities we expected solid and comparable negative priming effects.
Procedure
Figure 1 shows a typical trial combination in Experiment 1. Each trial combination was initiated by pressing the space bar with the left hand. Four empty squares followed, which indicated potential locations of the prime stimuli. After 500 ms, a target alone or a target and a distractor appeared abruptly within the squares. Participants had to respond to the location of the prime target “O” with the corresponding key on the keyboard. Prime events disappeared with prime response execution and were replaced by four empty squares. After 300 ms the probe stimuli were displayed within the squares. Responding to the probe target location finished the trial combination and the word “next” was shown. Participants had to initiate the next trial combination on their own by pressing the space bar. This procedure allowed us to connect prime and probe trials as closely as possible and to distinguish them as a separate trial combination from the following ones. The interruption of the “trial flow” caused by the word “next” was thought to prevent effects of the previous trial combination (n−1) from influencing the processing of the current trial combination. The longer presentation time of the placeholders in the prime-trial was meant to soften potential effects caused by an orientation reaction at the beginning of a new trial combination.
Results
Prime trial
Correct RTs for each participant in each condition were subjected to an outlier elimination procedure to avoid the inclusion of suspiciously small or large RTs in further analyses. For this reason, we decided to set a definite minimum RT barrier of 150 ms (Fox, 1994). In addition, we excluded all RTs that exceeded the size of two standard deviations of the mean RT per participant and condition. This outlier analysis was done prior to all statistical analyses in all experiments for prime and probe RTs, and therefore, will not be mentioned again. In Table 1 the main results of the prime trial analysis are summarized. Considering the simplicity of the task, the extremely low rate of total errors of .5% was not surprising. Due to the low frequency of errors, no further error analysis was done. The outlier analysis and the error trials in the prime condition resulted in an elimination of 4.5% of the RTs. An interference effect of the summarized conditions (PD & ND and IR & C) of 26.05 ms was significant, t(24) = −9.78, p < .001.
Probe trial
Table 1 also shows results of the probe trial analyses. Similar to the error rate of the prime trial, the total error frequency in the probe trial was also very low (1.0%). A further error analysis was not conducted. Incorrect RTs of the probe trial, as well as incorrect RTs of the prime trial, were excluded from further analyses. Together with the outlier analysis, this resulted in a 5.7% elimination of RTs.
A repeated single factor ANOVA, which treated condition (IR, C, PD, and ND) as a lone variable, was conducted on the RT data. The analysis revealed a significant main effect of the within-participant factor, F(3,72) = 106.81, MSE = 226.18, p < .001. Probe trials that included a distractor (PD condition) resulted in longer RTs than probe trials without a distractor (ND condition). A repeated contrast analysis proved this interference effect of 24 ms as significant, F(1,24) = 34.41, MSE = 418.44, p < .001. When the probe target appeared in the same location as the prime distractor (IR), longer RTs were observed compared with conditions in which all positions of stimuli changed (C). This negative priming effect of 50 ms was also significant, F(1,24) = 105.61, MSE = 598.29, p < .001. Interference and negative priming effects for Experiment 1 are summarized in Table 2.
Discussion
The results of Experiment 1 are in accordance with findings of other location-based negative priming studies (Tipper et al. 1994). Significant interference effects of about 25 ms demonstrate the influence of irrelevant information while processing the relevant information. A strong negative priming effect of 50 ms, regarded as a replication of the results of Tipper, Weaver, Cameron, Brehaut, and Bastedo (1991) and Reder et al. (2003; negative priming effects of 30 and 40 ms respectively), legitimates our experimental design for further investigations.
We can now view the negative priming effects we found in Experiment 1 in the context of selection and attentional control. Both stimuli, target and distractor appeared abruptly on the screen (onset presentation), which requires the creation of new object representations and leads to an involuntary shift of visual attention to the stimuli. Thus, both onset stimuli capture visual attention automatically and compete for control of action.
There is evidence that onset effects in capturing visual attention can be modulated through top–down influences based on attentional control settings (Folk, Remington, & Johnston, 1992; Folk & Remington, 1999). That means that task goals can reveal strong impacts on the selection process and also reveal whether a distractor interferes with performance (Desimone & Duncan, 1995; Downing, 2000). However, strong interference effects in Experiment 1 indicate competition for selection and demonstrate that the instructional knowledge about the pre-defined, never changing target did (at least in our case) not lead to a major advantage of encoding this target. At the same time there may well have been benefits from foreknowledge about target identity that were not observable here since Experiment 1 did not include conditions without this knowledge. However, in the context of this study, the important aspect is that the foreknowledge about target identity did not result in benefits that eliminated interference by a present onset distractor. This is in accordance with visual search studies in which it was shown that despite specific featural knowledge about an upcoming target (Theeuwes, 1992) or incentives to prioritize a certain set of elements in which a target could appear (Donk & Theeuwes, 2003), participants were unable to prevent attentional capture by an irrelevant singleton. Our results indicate that a potential attentional control setting in terms of featural knowledge of an upcoming target seems not to bias the selection process to the degree of interference-free selection when both stimuli appear as abrupt onsets.
Experiment 2: Negative priming and a non-automatic deployment of visual attention
Since we were able to demonstrate stable negative priming effects in Experiment 1, the second experiment was designed to investigate whether the inhibitory mechanism that is responsible for negative priming effects can be disengaged when the allocation of visual attention to the stimuli is manipulated. That is why in Experiment 2 we included a no-onset stimulus presentation representing a non-automatic allocation of visual attention. As stated above, the encoding process of no-onset stimuli in visual search is often seen as a goal-directed, serial, and self-terminating search (Yantis, 1998). If one follows that argumentation, it seems plausible that visual attention is not automatically captured by either no-onset target or no-onset distractor. This means that, because there is no strong stimulus driven attentional capture involved in Experiment 2, the deployment of visual attention is highly influenced by top–down processes such as the strategies and expectations of the participants. Top–down settings, such as the instructional knowledge about target identity, might bias encoding and processing priorities between target and distractor (see Awh et al., 2003) and thus alter the selection process. We argue that in conditions of top–down allocation of visual attention the priority of stimulus processing is biased toward the goal-relevant information that results in a top–down modulated, biased selection demand within the negative priming task (top–down selection).
This means that presenting stimuli in a no-onset mode provides a top–down setting that might facilitate efficient target identification and selection. Because of this top–down support in target identification and because a no-onset distractor item is not supposed to automatically capture attention, its impact within the selection process should be strongly reduced, which should be reflected in reduced interference effects. More importantly, without the distractor attracting attention it seems plausible to assume that inhibitory processes, usually at work to reduce distractor influences, are no longer required in order to accomplish target selection. This should be relevant for prime and probe trials since the no-onset presentation was applied for the whole trial sequence. In this line of argumentation, an implemented top–down allocation of visual attention can provide a condition in which the inhibitory mechanism in the selection task can be disengaged because it is not needed throughout the experiment. If this proves to be true, then we should not observe reliable negative priming effects.
Procedure
The structure of Experiment 2 is similar to Experiment 1 (see also Fig. 3). The placeholders appear at the beginning of a trial containing a mask in each square. The mask is created by placing target and distractor on top of each other. The realization of the no-onset stimuli is adapted from the original version (Jonides & Yantis, 1988) in which a digital 8 contained the letter that was identifiable only after two line segments were removed. In our experiment all stimuli will be removed from two squares, whereas in each of the other two squares only one item disappears so that a target and a distractor will remain. The same experimental conditions as in Experiment 1 were applied.

Experimental design and time characteristics of an ignored repetition (IR) trial combination in Experiment 2.
Results
Prime trial
The statistical analyses of Experiment 2 were carried out in a similar manner to Experiment 1. Total error rate was also very low (.6%), thus we did not conduct any further error analysis. The outlier analysis and the error trials of the prime trial resulted in a 4.6% elimination of RTs. In Table 3 the results of Experiment 2 are listed. RTs of prime trial conditions including a distractor (IR and C summarized) did not differ from conditions without a distractor (PD and ND summarized). An interference effect of 3.46 ms was not significant, t(24) = −.94, p = .356.
Probe trial
Total error rate of the probe trials in Experiment 2 was .8%. Error trials of prime and probe trials plus the outlier analysis resulted in a total elimination of 6.0% of RTs. A repeated ANOVA with the single factor condition (IR, C, PD, and ND) revealed no significance, F(3,72) = 2.06, MSE = 191.54, p = .113. A repeated contrast analysis demonstrated that RTs in the IR condition (probe target follows prime distractor location) did not differ from RTs in the control condition (all positions changed). The obtained negative priming effect of 5.81 ms was not significant, F(1,24) = 1.38, MSE = 610.34, p = .252. Although the interference effect (conditions with a distractor compared with conditions without one) of 6.79 ms was not much higher, the repeated contrast analysis showed significance, F(1,24) = 5.71, MSE = 201.50, p = .025. Interference and negative priming effects for Experiment 2 are listed in Table 2.
Discussion
Experiment 2 provides us with quite interesting results. Conducting a negative priming experiment using a no-onset stimulus presentation leads to a complete elimination of commonly observed negative priming effects. Moreover, interference effects of the distractor stimuli were not obtained in the prime trial and were substantially reduced by 70% in the probe trial, F(1,24) = 13.18, MSE = 562.32, p = .001 (see the Results section of Experiment 3 for a cross experiment analysis).
This result is in line with our argumentation. The no-onset presentation of stimuli represents a completely different control of visual attention than has so far been used in classical negative priming paradigms. We argued that only in conditions in which the target and the distractor stimulus capture visual attention automatically strong competition for selection is given and the resulting distractor inhibition will cause negative priming in subsequent IR trials. No-onset stimuli such as those in Experiment 2 do not share the specific characteristics of capturing visual attention automatically. As stated above, the deployment of visual attention to those items is much more influenced, if not regulated, by external factors such as expectation and/or task goals. Therefore, one can assume that not the perceptual characteristics of no-onset stimuli per se direct attention in a stimulus-driven manner, but the explicit knowledge about the features of the target.
In other words, when stimuli do not capture attention automatically the influence of an attentional control setting might come into play and guide the selection process. One could imagine that participants might translate the instructional goals into attentional control settings that specify which features of the stimuli are relevant. Preparing for searching a circle may result in tuning a passive input filter in such a way that the stimulus possessing this feature can pass and receive prioritized processing (Folk et al., 1992; Pratt & Hommel, 2003). One could go even further and assume that in such top–down settings a stimulus that matches the goal representation will be automatically processed in terms of a “cognitive reflex” (Hommel, 2000).
Concerning the target, this would lead to a top-down modulated prioritized processing in terms of a more direct translation of visual information into a response Footnote 1 . External factors allocate visual attention more directly to the relevant information so that irrelevant information might even be less attended to if not neglected. In such a feature detection mode, irrelevant items no longer interfere (Bacon & Egeth, 1994). This is exactly what we observed concerning the no-onset distractor item. Responding to the target was either not influenced at all by a present distractor (prime trial) or was barely affected by it (probe trial). The attentional control setting assumed to play an increased role when stimuli are presented as no-onsets may enhance target detection so that the inhibition of a no-onset distractor that does not capture attention might become unnecessary. In classical terms of negative priming the loss of distractor inhibition in the prime trial would consequently result in unobserved negative priming effects. However, at this point we do not know whether a hypothetical attentional filter is capable of eliminating the processing of no-onset distractor items. In terms of an early selection theory, one could assume that in the applied top–down setting an irrelevant item cannot pass the attentional filter (Broadbent, 1958). On the other hand, such a filter could also work on enhancing and facilitating feature processing that matches the goal representation rather than blocking other features from processing. In any case, our results clearly show that no-onset distractors did not reveal any influence on prime target selection regardless of any processing level.
Although our findings are in line with our predictions that top–down settings such as the goal-directed deployment of attention can alter the selection process and provide an option for disengaging the negative priming mechanism, it is necessary to explore alternative explanations.
-
1.
The elimination of negative priming effects in a no-onset stimulus presentation cannot be explained by forward masking. According to Fox (1994), forward masking reduces interference, but not negative priming. In our Experiment 2, both are reduced.
On the other hand, one could also view the mask placeholders in Experiment 2 as an intervening event that could affect processing in the negative priming procedure by decoupling prime and probe trial. According to Tipper et al. (1991), this also seems unlikely since they could show that predictable intervening events do not interrupt inhibitory processes in location negative priming. The possibility that mask placeholders as intervening events would block prime trial information from affecting probe trial processing can be investigated by using the no-onset presentation for probe trial stimuli only (see Experiment 4B)
-
2.
In an IOR paradigm, Samuel and Weiner (2001) argue that the disappearance of a stimulus itself produces early inhibitory consequences. Our study differs in that such attentional consequences revealed by disappearing stimuli (our mask placeholders) are not only confined to the relevant information, which was the case in their study. Instead, viewing the mask placeholders in all locations as a single stimulus binding attention to the locations (Cepeda, Cave, Bichot, & Kim, 1998), target and distractor are equally exposed to the consequences of the disappearing mask in the placeholders. Thus, this argument also seems insufficient to explain the elimination of negative priming effects in our design
-
3.
Since the same participants performed Experiments 1 and 2, it is interesting to note that RTs of IR trials in Experiment 2 are virtually the same as in Experiment 1. Because there are negative priming effects in Experiment 1 but none in Experiment 2, one could attribute the removal of negative priming to increased RTs in the control condition (C) of Experiment 2 and less to a reduction in IR trial RTs to control level, as would be the natural way. On the other hand, compared with Experiment 1, RTs of Experiment 2 were generally increased in the C, PD, and ND conditions respectively. Therefore, from this point one would also have to expect increased RTs in the IR condition, which was in fact not the case. That is why we argue that the removal of negative priming is indeed due to a reduction in RTs in the critical IR trials because this is the only condition in Experiment 2 in which RTs did not increase.
Also, we further assessed possible practice effects as a potential cause of the elimination of negative priming effects in Experiment 2. For this reason we contrasted negative priming effects of participants who performed Experiment 2 first with those who completed it second. However, there was no indication that practice would lead to a reduction in negative priming, t(23) = 2.06, p > .05). On the contrary, the analysis showed a slight but non-significant tendency of increased negative priming for those participants who performed Experiment 2 after Experiment 1
-
4.
Another objection concerns the possibility of a pre-processing of the target item during the presentation of the mask placeholders. We have used special no-onset stimuli by placing target and distractor on top of each other. Removing one stimulus from this overlapping situation reveals the actual stimulus for the current display. Now, imagine that participants are able to reduce the mask placeholders in each location by mentally removing the distractor stimulus from the mask leaving only the target stimuli in all locations. This option would also predict a biased selection, which is contrary to our assumption based on the characteristics of the mask placeholders in Experiment 2 and would, therefore, provide a competing account of our findings. This motivated our third experiment in which we used a different no-onset mask
Experiment 3: Pre-processing of target information?
As already stressed, our participants had clear knowledge of the identity of the upcoming relevant (O) and irrelevant (X) stimuli. Only the task-relevant location of the stimuli was not predictable. In Experiment 2, the pre-defined stimuli in their role as target and distractor appeared together in the form of mask placeholders several hundred milliseconds before the actual selection took place (see also Fig. 3). From task switching, it is known that a stimulus has to be available in its physical form before a system is able to adjust to the situational demand (e.g., a certain response) revealed by this stimulus (Rogers & Monsell, 1995; Meiran, 1996; Sohn & Carlson, 2000). This situation might be transferable to our study. Because the mask placeholders (target and distractor together in each location) contain goal information, one could argue that, based on the instruction, the information processing system has the advantage of perceptually adjusting to the required selection of the target (O) before the actual selection takes place. The question is whether it could be possible that the target receives favored processing over the distractor based on its physical presence in the mask. Thus, the instruction itself may instigate the creation of a target object representation when the mask placeholders are presented, which then serves as a basis for an adequate response to the location of the target. One could imagine that this results in a 100% top–down effect, disregarding the distractor representation. This would probably have the same consequences of removing negative priming, but based on different origins as suggested so far in this study. In their model of selective attention Houghton and Tipper (1994) assumed that different object representations compete for control of action. Applied to our Experiment 2, it seems possible that the target representation is advantaged in this competition due to the knowledge about the target and its physical presence in the mask. In general, this bias could potentially reduce the demand for selection when target and distractor are revealed by the mask placeholders, which could account for the removal of negative priming.
In order to disprove this hypothesis, we conducted a third experiment in which we implemented mask placeholders without any goal information. If this variation were to lead to the same results as in Experiment 2, the argument that target pre-processing was responsible for the missing negative priming effects would then become rather unlikely.
Procedure
Experiments 3 and 2 differed only in the design of the no-onset mask placeholders. Whereas in Experiment 2 both stimuli (O and X) served to occupy each square, in Experiment 3 the symbol “&” was used as a substitution to create the placeholder. In this variation, the mask placeholders did not contain any goal information.
Note that using ampersands as masks leads to a slightly different no-onset presentation than in Experiment 2. Whereas in Experiment 2 one stimulus disappeared in the mask to reveal the target or distractor item, now the ampersands were replaced by the target and the distractor. Taking this strictly means that in the no-onset presentation of Experiment 3 target as well as distractor item can also be viewed as onset stimuli, appearing in formerly occupied locations (ampersand mask). However, in our study we think this is of lesser concern. Important for the manipulation of attention control is the contrast between onset and no-onset events. The essential point is that in onset presentations all locations are empty before stimulus appearance. In the context of this study, the crucial characteristic of the no-onset presentation is that all locations are occupied by a perceptual event (mask) before stimulus appearance. Thus, stimuli appear in formerly occupied locations. Given this primary characteristic of a no-onset presentation as applied in our study, Experiments 2 and 3 are functionally the same.
Results
Prime trial
Error rate was .5%. No further error analyses were carried out. Error trials and outlier analysis revealed an elimination rate of 4.3% of RTs. Investigating the influence of a distractor on processing the target led to an interference effect of 3.88 ms, which reached significance, t(24) = −2.29, p = .031. The results of the prime trial analysis are presented in Table 4.
Probe trial
A total error rate of .8% is in accordance with our previous experiments. No further error analysis was conducted. Error trials of prime and probe condition and outlier analyses produced a total elimination rate of 5.3% of RTs. The IR and C conditions revealed a very small negative priming effect of 2.42 ms. The conditions of probe distractor (PD) and no distractor (ND) show an only slightly higher interference effect (8.45 ms) compared with Experiment 2 (6.79 ms). A repeated ANOVA of RTs with the single factor condition (IR, C, PD, ND) showed a significance of F(3,72) = 8.18, MSE = 91.17, p < .001. A repeated contrast analysis obtained no significance of the negative priming effect (F < 1), but a significant interference effect, F(1,24) = 14.01, MSE = 127.60, p = .001. Interference and negative priming effects for Experiment 3 are summarized in Table 2.
Because all participants performed all three experiments, an ANOVA was conducted to compare negative priming and interference effects between the experiments when varying the presentation mode (onset vs. no-onset). A repeated contrast analysis revealed that both negative priming and interference effects were significantly reduced when changing an onset presentation (Experiment 1) to a no-onset presentation of stimuli (Experiment 2), F(1,24) = 57.51, MSE = 859.71, p < .001 for negative priming effects and F(1,24) = 13.18, MSE = 562.32, p = .001 for interference effects respectively. At the same time, both no-onset experiments (2 and 3) did not differ in the magnitude of negative priming (F < 1) and interference effects (F < 1) respectively.
Discussion
Experiment 3 was carried out to disprove the argument of a pre-processing of the target as a cause of missing negative priming effects in Experiment 2. The implementation of mask placeholders without any goal information in Experiment 3 offered no possibility of an advantaged processing of the target. The results are straightforward; no negative priming effects could be observed either. Although significant interference effects were found, they were also strongly reduced and did not differ in their magnitude from those in Experiment 2. Also, the observed removal of negative priming cannot be attributed to increased RTs in control trials. RTs for IR and control trials are virtually the same, if not even shorter, than RTs in the control trials of Experiment 1. This means that the elimination of negative priming effects is achieved through a reduction of IR trial RTs. Even though a different no-onset presentation was used for Experiment 3, the main results did not differ in the two no-onset experiments. This is further support for our assumption that the current design of the mask placeholders might provide the same attentional control setting in which the possibility of disengaging the inhibitory mechanisms of negative priming is given.
Experiment 4: Onset and no-onset presentation of stimuli
Even though in Experiments 2 and 3 we demonstrated that a removal of negative priming can be found when changing the presentation mode of stimuli for prime and probe trials, there are at least two remaining questions that need further consideration. A first question concerns the selection process in conditions of no-onset stimulus presentation. We argued that a no-onset presentation of stimuli results in a feature detection mode in which a goal-directed top-down selection favors target encoding based on its known features. However, by arguing in favor of an increased efficiency of target selection, it is still not clear what exactly happens to the distractor item. In no-onset conditions, the prime distractor in particular did not seem to reveal any impact on target selection. Therefore, the question is, does an eliminated impact of no-onset prime distractors automatically imply that these distractors are generally not processed?
This question was addressed in Experiment 4A. We have shown that a no-onset distractor does not (or only minimally) compete for selection. This might be due to an attentional filter that biases stimulus selection in terms of prioritized encoding of task-relevant stimulus features that the distractor item does not possess. However, it is known that interference can tell us about competition for action when selecting a relevant item amongst irrelevant items. On the other hand, interference effects are an unreliable source when it comes to conclusions about the level of processing of an irrelevant item (e.g., Stablum et al., 2001). In other words, even though we demonstrated no impact of no-onset distractors in the selection process, we do not know whether distractors are generally not processed when presented as no-onset stimuli. For this purpose, in Experiment 4A, prime trials were always presented as no-onset stimuli (as in Experiment 3) whereas probe trials represented onset stimuli (as in Experiment 1). The classical negative priming view holds that the prime trial selection, and more specifically the prime distractor inhibition, determines the occurrence of negative priming in the subsequent probe trial. Thus, if a no-onset presentation of prime stimuli results in a complete non-processing of the no-onset distractor, negative priming effects should not be observed in Experiment 4A.
The second question that needs to be investigated in more detail concerns the removal of negative priming in no-onset conditions and the hypothetical mechanism responsible for this removal. In the Discussion of Experiment 2, we briefly introduced a masking account as an alternative explanation of diminished negative priming effects. One could argue that whenever there is a mask placeholder between prime and probe trials both trial types are segregated in a way that any inhibitory information of the prime trial becomes neutralized or eliminated by the perceptually strong masks. Therefore, each trial would then be processed completely independently and isolated from each other so that influences of prime stimuli on probe trial processing and vice versa (e.g., retrieval of prime information when processing the probe trial) would be blocked and could not take place. In such a scenario, the elimination of negative priming effects would be an inevitable consequence of the no-onset procedure in Experiments 2 and 3. The aim of Experiment 4B was therefore to disprove a masking explanation of our results. For this purpose, prime trials of Experiment 4B contained abrupt onset stimuli (as in Experiment 1) and probe trials included no-onset stimuli (as in Experiment 3). In such a design a mask placeholder which revealed the subsequent no-onset stimuli is positioned between prime and probe trial. If a perceptually strong mask would block any prime trial information from influencing probe trial processing, we should not observe negative priming effects. On the other hand, negative priming effects despite a no-onset presentation in the probe trial would most likely discount a masking explanation.
The common aspect of both experiments is that they include onset as well as no-onset stimulus presentation for either the prime or the probe trial respectively. This goes beyond the manipulations in the previous experiments (1–3), in which all attentional manipulations were equally applied to prime and probe trials. Therefore, the results of Experiments 4A and 4B will also be informative about the disengagement of the inhibitory mechanism when competition for selection is required in at least one trial of the sequence (prime or probe).
Participants
Fifteen students (10 females) of the Department of Psychology at Humboldt University, Berlin took part in exchange for course credits. Participants had not performed Experiments 1–3. Their ages ranged from 20 to 27 years (mean age 24.7 years). All students were right-handed and had normal or corrected-to-normal visual acuity.
Procedure
Prime trials in Experiment 4A characterize a no-onset presentation mode whereas probe trials are presented in an onset presentation mode. Experiment 4B (onset/no-onset) also included both presentation modes, but they were exactly reversed between prime and probe trial. Participants started the experiment with a block in which they practiced the simple response mode. Positions were highlighted and participants had to press the corresponding key on the keyboard. Sixty-four practice trials were used. After completing the practice part, seven participants started with Experiment 4A whereas the other eight participants started with Experiment 4B. Within one session, all participants completed both parts of Experiment 4. Except for the reported changes, the experimental conditions and apparatus were the same as in the first three experiments.
Results
Experiment 4A (no-onset/onset) prime trial
A low error rate of 1.1% was found for the prime trials, which revealed together with the conducted outlier analysis an elimination rate of 4.8%. Table 5 gives an overview of the main results of the prime trial analysis. The interference effect (4.5 ms) of prime trial distractors on target processing did not differ numerically from the prime trial interference effects in Experiments 2 and 3 and also demonstrated a marginal significance of t(14) = −2.20, p = .045.
Experiment 4A probe trial
A low error rate of 1.4% and the outlier analysis resulted in a total elimination of RTs of 6.0%. Main results are listed in Table 5. Experiment 4A, in which the no-onset presentation of the stimuli was implemented in the prime trial, yielded an interference effect of 29.3 ms between the conditions “no distractor” (ND) and “probe distractor” (PD), which seems comparable to Experiment 1 (24 ms). In the ignored repetition condition, reliable negative priming of 35.2 ms was found. A repeated single factor ANOVA revealed a significant main effect of the within-participant factor (conditions), F(3,42) = 68.44, MSE = 147.76, p < .001. A repeated contrast analysis demonstrated strong significance of both the interference effect, F(1,14) = 84.05, MSE = 153.67, p < .001, as well as the negative priming effect, F(1,14) = 58.87, MSE = 315.14, p < .001.
Experiment 4B (onset/no-onset) prime trial
An error rate of 1.1% was found for the prime trials, which revealed together with the conducted outlier analysis a total elimination rate of 7.6%. Table 6 gives an overview of the main results of the prime trial analysis. In Experiment 4B, stimuli of the prime trial were presented in an onset mode exactly as in Experiment 1. Therefore, it is not surprising that RTs for prime trial conditions including a distractor (IR and C summarized) and conditions without a distractor (PD and ND summarized) reveal an interference effect (t(14) = −7.74, p < .001) identical to the one in Experiment 1 (26.12 vs. 26.05 ms).
Probe trial
A very low error rate of 0.7% and the conducted outlier analysis resulted in a total elimination of 8.1% of trials. Results of the probe trial analysis are shown in Table 6. In Experiment 4B only the stimuli of the probe trial were presented as no-onset stimuli. This experimental manipulation resulted in strongly reduced interference effects of 8.3 ms and negative priming effects of 14.9 ms respectively. A repeated ANOVA of RTs with the single factor condition (IR, C, PD, and ND) showed significance, F(3,42) = 11.85, MSE = 82.41, p < .001. A repeated contrast analysis demonstrated significance for both the interference effect, F(1,14) = 6.71, MSE = 152.55, p = .021, as well as the negative priming effect, F(1,14) = 26.54, MSE = 125.66, p < .001.
Comparison of Experiments 4A and 4B
Since both experimental parts were performed by the same participants, effects of interference in prime trials and effects of interference and negative priming in probe trials were compared with respect to the order of presentation mode of stimuli (Part 4A vs. Part 4B). Concerning the prime trials, interference effects were much larger in Experiment 4B containing onset prime stimuli, F(1,14) = 26.94, MSE = 64.96, p < .001) compared with Experiment 4A in which no-onset stimuli were used. Larger interference effects in probe trials were also obtained when stimuli were presented as onset stimuli (Experiment 4A) compared with a no-onset presentation, F(1,14) = 17.22, MSE = 387.15, p = .001. Stronger negative priming effects were found when no-onset stimuli were used in the prime trial and onset stimuli in the probe trial (Experiment 4A) compared with the reversed order in Experiment 4B, F(1,14) = 16.11, MSE = 382.17, p = .001.
When participants performed Experiments 4A and 4B, the order of 4A and 4B was balanced. Eight participants performed Experiment 4B (onset/no-onset) first and experiment 4A (no-onset/onset) afterwards and seven had the reversed order. A potential influence of order of performance was investigated conducting a repeated ANOVA including a within-participant factor (condition, IR, C, PD, and ND) and between a within-participant factor (order, A first, B first). The within-participant factor revealed no interaction between condition and order (F < 1). The between-participant factor did not show any significance in terms of order either, F(2,13) = 1.12, MSE = 4560.20, p = .309.
Discussion
Experiment 4 consisted of two parts. Experiment 4A was designed to shed further light on the selection process in conditions of no-onset presentations. In particular, we wanted to investigate whether an efficient target selection automatically goes along with an insufficient or even a non-processing of distractor items in a no-onset presentation. For this purpose, only prime trial stimuli were presented as no-onsets whereas probe trial stimuli appeared in an onset presentation mode. If a no-onset presentation of stimuli leads to an exclusion of distractor processing, we should not have expected negative priming effects. As before, in conditions of a non-automatic deployment of visual attention, the no-onset distractor failed to reveal a considerable impact on prime target selection. The no-onset target was once again efficiently selected and interference due to a no-onset distractor in the prime trial was barely seen (4.5 ms). Nevertheless, the results are somewhat surprising if one assumes a non-processing of distractor items in a no-onset presentation. Strong negative priming effects were observed, which rule out no-onset distractors being completely filtered out and not processed. This result provides further evidence that missing interference effects do not allow inferences about the processing level of irrelevant information (Fox, 1995; Richards, 1999; Stablum et al., 2001). Thus, even in a no-onset presentation of stimuli in which a distractor does not compete with target selection, irrelevant distractor items can nevertheless be processed and can lead to negative priming effects in the subsequent trial.
This result has some implications for the understanding of how a no-onset presentation and the assumed voluntary deployment of visual attention to the stimuli affect the selection process. Since both relevant and irrelevant no-onset prime stimuli are processed, the assumption of an attentional filter blocking stimuli that do not possess goal-relevant features from general processing seems insufficient to explain the results of Experiment 4A. A voluntary deployment of visual attention in conditions of a no-onset presentation may indeed prioritize target encoding and processing, but nevertheless, seems not to be exclusively directed at the relevant target. In more speculative terms, considering that both no-onset stimuli were displayed until response execution, there would be no reason why an irrelevant item should not be attended (even inhibited) after the identification of the target. Attending the irrelevant no-onset item in the prime trial of Experiment 4A could be motivated by the onset probe trial, in which a distractor affects processing. In other words, it could be a voluntary or involuntary consequence of the probe trial bottom–up selection in which both onset stimuli capture attention automatically and distractor inhibition is necessary for target selection (see General discussion for further elaboration).
The aim of Experiment 4A was to investigate whether a no-onset presentation would automatically exclude irrelevant information from being processed. This was clearly not the case. We demonstrated that a no-onset prime distractor that does not influence target selection can still be subject to inhibition when distractor inhibition is required in the probe trial. These results provide further evidence for the claim that the mechanism responsible for negative priming is quite adaptable to task requirements.
Experiment 4B was designed to further explore the removal of negative priming in Experiments 2 and 3. An alternative view to the disengagement of the negative priming mechanism due to the attentional control setting was described in a masking account. Here it was argued that perceptually strong mask placeholders between prime and probe trials could possibly eliminate or block prime trial influences on probe trial processing. The results of Experiment 4B speak rather against this alternative. Presenting only probe stimuli as no-onsets had the effect that the mask revealing the probe stimuli followed the prime and preceded the probe trial. The demonstration of negative priming effects despite such a “perceptually intervening event” does not fit well an explanation in which a mask placeholder would completely isolate prime trial and probe trial processing. Thus, masking alone seems unlikely to completely account for the removal of negative priming in Experiment 2 or 3. Nevertheless, negative priming effects in Experiment 4B were indeed smaller than in Experiment 4A, for which there could be several reasons. First, one could argue that some form of masking could at least partially contribute to a reduction in negative priming when no-onset stimuli were used in the probe trial. A second possibility to account for different negative priming effects in Experiments 4A and 4B could be seen in the changes of presentation modes within each experiment. Changing the presentation mode from prime to probe could lead to strong changes in attentional control settings (see below). An onset presentation, going along with strong interference between distractor and target in any trial (prime or probe), might cause negative priming. This effect might be increased for probe trials because they are the last event in the sequence and are more likely to be remembered when participants begin the subsequent trial. Footnote 2
The observation of negative priming in Experiment 4A as well as in 4B provides important insights concerning the flexibility of the inhibitory mechanism responsible for negative priming. Whereas in conditions of top–down allocation of attention negative priming effects were not found (Experiments 2 and 3), the inhibitory mechanism seems to be “turned back on” as soon as prime or probe trial contain any instance of onset distractors, revealing an impact on target selection. Therefore, the implementation of any onset event, in which distractors influence processing, seems to lead to global changes in top–down settings probably affecting processing throughout the whole trial sequence (for further debate see General discussion).
General discussion
The aim of the present study was to demonstrate the flexible and adaptive nature of the underlying mechanism in a localization task such as negative priming. In previous studies, it was assumed that such inhibitory mechanisms can be voluntarily disengaged by participants when utility information via cuing is provided (Buckolz et al. 2002a, 2002b). In our study, we demonstrated a possibility of disengagement when the allocation of visual attention to the stimuli was manipulated. Differences in the deployment of visual attention were achieved by varying the presentation mode of stimuli. Presenting target and distractor as abruptly appearing onset stimuli was seen as an automatic or bottom–up control of visual attention because both onset stimuli capture attention automatically. In such an onset presentation mode, strong negative priming effects were found (Experiment 1). A non-automatic or top–down control of visual attention was implemented by presenting target and distractor as no-onset stimuli, which are not supposed to capture attention automatically. Here it is assumed that attention can be voluntarily deployed to the stimuli based on task goals and attentional control settings. In Experiment 2, in which all stimuli were no-onsets, negative priming effects were eliminated. This result was replicated in Experiment 3 in which a different kind of no-onset presentation was used. Thus, we demonstrated that changes in the presentation mode lead to changes in negative priming. In order to understand why a no-onset presentation of stimuli can lead to a disengagement option for the negative priming mechanism, we have to go back to two questions raised at the beginning of this paper: First, how does a change of presentation mode of stimuli affect the selection process? Second, what enables participants to disengage the negative priming mechanism?
The presentation mode of stimuli affects target selection
In a typical onset presentation of stimuli, both target and distractor capture attention automatically and compete for action (Yantis & Johnson, 1990; Yantis & Jones, 1991). Target selection is accomplished by distractor inhibition. That an onset distractor competes for selection is seen in strong interference effects in Experiment 1. Even though the target item was pre-defined and never changed, an attentional control setting (Folk et al., 1992) concerning specific featural knowledge about the upcoming target was not able to bias the selection process in terms of interference-free target processing when all stimuli were presented as abrupt onsets.
A no-onset presentation of stimuli works quite differently. Target and distractor do not automatically capture visual attention so that the allocation of visual attention is more strongly influenced by top–down processes (e.g., Yantis, 1998). In other words, the absence of strong stimulus-driven attentional capture enables attentional control settings to guide and bias selection. The knowledge of the specific target features might lead to a direct allocation of attention to the item possessing these features resulting in prioritized processing (Awh et al., 2003; Bacon & Egeth, 1994; Folk et al., 1992; Pratt & Hommel, 2003). Such a top–down-supported efficient target selection should scarcely be influenced by the presence of irrelevant items. This is what we observed in Experiments 2 and 3. Eliminated or strongly reduced interference effects indicate that the presence of a no-onset distractor barely revealed any impact on selecting the no-onset target. Referring to our results, we conclude that in conditions without strong attentional capture by target and distractor an attentional control setting can come into play and alter the selection process in terms of efficient and prioritized target encoding and processing.
Assuming prioritized no-onset target processing, one might wonder about the nature of no-onset distractor processing. Since a no-onset distractor revealed only minimal if any impact at all on target selection, we investigated whether a no-onset presentation may automatically go along with non-processing of the no-onset distractor (Experiment 4A). Presenting prime stimuli as no-onsets and probe stimuli as onsets surprisingly resulted in strong negative priming effects. The observation of negative priming effects in such a condition means that a no-onset prime distractor was inhibited and thus disregards any non-processing hypothesis of no-onset distractors as an automatic consequence of the presentation mode. Even though no-onset prime distractors do not compete for selection they can nevertheless be processed and are even subject to inhibition when probe trials require distractor inhibition.
One way to think about how selection could be accomplished in such a no-onset condition is to assume that a no-onset distractor is processed simply after the no-onset target is identified and selected. According to this assumption, missing interference effects would be a plausible consequence of a no-onset presentation. No-onset distractors do not interfere because they are attended to after target selection. There are many studies which investigated the time course of attentional shifts between objects in a display (Theeuwes, Godijn, & Pratt, 2004; Woodman & Luck, 1999). Considering that in our design all no-onset stimuli were displayed until response execution, there would be sufficient time to attend the distractor after target selection. Such an assumption is of course hypothetical and cannot be proven on the basis of our study. An important question is, whether no-onset presentations would generally lead to this kind of processing order or whether the no-onset distractor in the prime trial of Experiment 4A is only processed as a consequence of the implemented onset presentation in the probe trial in which the onset distractor needs to be inhibited for target selection.
Another way to think about target selection in no-onset conditions is to assume that no-onset distractor processing is attentionless. Footnote 3 Here, distractor processing would be carried out automatically and does not cause interference. According to this line, target and distractor processing would be done in parallel, rather than sequentially as suggested above.
However, at this point, we can only speculate about distractor processing when a no-onset presentation is given in prime and probe trials (Experiments 2 and 3). At any rate, Experiment 4A clearly showed that presenting stimuli as no-onsets does not necessarily exclude a no-onset distractor from being processed. Even though no-onset distractors in the prime trial seem not to influence target selection, they are also subject to inhibition when distractor inhibition is required in the onset probe trial. This is an important result and provides further evidence for the flexible nature of the inhibitory mechanism in adapting to task demands in a negative priming paradigm.
The disengagement of the inhibitory mechanism
In this study, we showed that presenting stimuli either as onsets or as no-onsets influences the occurrence of negative priming effects. We argued that the presentation mode of stimuli alters the process of target selection, which in turn results in a disengagement of the inhibitory mechanism responsible for negative priming. The results are straightforward. Presenting all stimuli as no-onsets leads to an elimination of negative priming effects (Experiments 2 and 3), which means that the inhibitory mechanism was disengaged. In contrast, negative priming is re-established when one trial, either prime or probe, requires onset distractor inhibition (Experiments 4A and 4B). Our results demonstrate that the inhibitory mechanism responsible for negative priming is quite adaptable and can be engaged or disengaged according to specific task demands (Buckolz et al. 2002a, 2002b). The observed flexibility of engaging or disengaging the inhibitory mechanism depending on situational demands raises the question whether the disengagement in no-onset presentations is brought about intentionally by participants. Buckolz et al. (2002b), for example, provided their participants with advance knowledge about the selection requirements in the negative priming task. An informative cue indicated whether participants had to handle a distractor event or not. When participants knew that selection in the probe trial was not required due to distractor absence, negative priming effects diminished. Here, the removal of the negative priming effect is intentional. The disengagement due to the manipulation of the presentation mode of stimuli can be seen in a similar light. Buckolz et al. (2002b) argue that the negative priming effect is dependent on the selection requirements of the task. In our study, participants might realize that distractors in a no-onset presentation (Experiments 2 and 3) have no impact on target selection. Therefore, participants may intentionally set aside the negative priming process because they notice that nothing happened in the no-onset prime trial that would potentially produce future slowing at the distractor location in the no-onset probe trial. Such an intentional view of disengagement becomes even more pronounced when looking at Experiments 4A and 4B. When both presentation modes are included in one experiment (Experiment 4A or 4B), the inhibitory mechanism is somehow held active, which resulted in negative priming effects. The two experiments differed in that no-onsets were used for prime trials and onsets were used for probe trials (Experiment 4A) or onsets for prime trials and no-onsets for probe trials (Experiment 4B). We argued that an automatic capture of attention (onsets) is a much stronger process than a non-automatic control in a no-onset presentation (Pratt & McAuliffe, 2001; Pratt & Hirshhorn, 2003; Samuel & Weiner, 2001). Thus, implementing both presentation modes in a single experiment might result in broad changes in the attentional control setting. When participants encountered a single trial with an onset presentation in which distractors interfere with target selection, the negative priming mechanism was kept on. Furthermore, we found stronger negative priming effects in Experiment 4A (onsets for probe trials) than in Experiment 4B (onsets for prime trials), which is indeed not easy to explain. One guess could be that when an onset trial is the last event in a sequence (Experiment 4A), participants might transfer the last event of processing to the next trial. This means that negative priming effects might be increased for onset probe trial conditions because this event is more likely to be remembered when participants start the following sequence.
Even though it is quite tempting to view our results in the light of an intentionally guided disengagement of the inhibitory mechanism, one has to be careful with such inferences. At this point, we cannot exclude the possibility that the disengagement of the inhibitory mechanism in Experiments 2 and 3 is purely unintended. Rather than an intended process, the disengagement can also be seen as an unintended by-product of the presentation mode of stimuli (e.g., reactive inhibition, masking). The top–down method of processing in a no-onset presentation reduces the distractor impact, which removes the potential for a negative priming effect. In other words, the top–down guidance of selection is indeed intentional. The modulation of negative priming, on the other hand, may be caused by an unintended effect of this top–down guidance on distractor processing in a no-onset presentation. Footnote 4
Thus, our study has provided evidence for the possibility of disengagement of the inhibitory mechanism when stimuli were presented as no-onsets. We assumed that when stimuli do not automatically capture attention the selection process is guided by top–down influences (attentional set) in which the distractor impact is reduced. However, at this point we cannot really differentiate whether the resulting disengagement is brought about intentionally by participants or whether it is an unintended by-product of the top–down processing.
Is the removal of negative priming in no-onset conditions a result of masking?
In order to provide a possible answer to this question it is essential to consider in which respect some form of masking could potentially be responsible for the elimination of negative priming. At least two ways are conceivable
First, the mask placeholders could function as intervening events and thus interrupt processing. According to this line, a masking account raises the possibility that mask placeholders would prevent prime trial information from influencing probe trial processing. To disprove this argument, in Experiment 4B prime stimuli were presented as onsets and probe stimuli as no-onsets. Negative priming effects should not have been observed if mask-placeholders of no-onset probe trials had segregated probe from prime trial processing. This was clearly not the case. Prime distractor inhibition was carried on to probe trial processing, which resulted in negative priming effects. Even though this priming effect was smaller than the effect in Experiment 4A, the demonstration of negative priming in such a condition makes a pure masking explanation for the removal of negative priming effects in Experiments 2 and 3 rather unlikely. This is also in line with Tipper et al. (1991, Experiment 4) who showed that in a target localization task predictable events intervening between prime and probe trials do not disrupt the maintenance of inhibition. The authors implemented a Go/No-Go task as an intervening event in which participants were required to respond, or to refrain from responding, depending on the identity of the intervening event stimulus. Thus, not only were visual stimuli presented between prime and probe trials (No-Go trials), participants even had to manually respond to the stimuli in Go trials. As reported by Tipper et al., negative priming effects were not impaired by the intervening events.
A second way of how masking could potentially be responsible for the elimination of negative priming is to assume that mask placeholders result in some kind of forward masking. Using a singleton search task, Gibson (1996), for example, argued that the placeholders in a no-onset presentation visibly persist beyond their physical offsets and that a visual integration of a mask with the subsequent no-onset stimulus delays the perception of that stimulus. Therefore, the disadvantage of attentional capture of no-onset stimuli compared with onset stimuli lies in pure visual masking due to the mask placeholders. In our context, one may assume that such forward masking could disrupt prime and probe processing as mentioned above and impair perceptual identification of subsequent no-onset stimuli, which in turn could be responsible for eliminated negative priming effects. The empirical evidence that Gibson takes into account consists of generally slowed RTs in no-onset displays compared with onset displays as observed in his study. We found similar results when comparing RTs of Experiment 1 (onset presentation) and Experiment 2 (no-onset presentation). However, using no-onset masks without any goal information in Experiment 3 (ampersands), negative priming and interference effects were diminished to the same degree as in Experiment 2 whereas RTs were at least as fast as in Experiment 1. Furthermore, DiLollo (1980) found that the degree of visual integration of the mask with the following no-onset stimulus (or in other words its visible persistence) was greatest for very short stimulus durations and was nearly lost when durations exceeded 200 ms. The duration of the mask placeholders in our experiments was 600 ms. Thus, the argumentation of visual forward masking should not pressure our preferred interpretation of the results.
Taken together, two conceivable forms of masking were introduced that could potentially account for the elimination of negative priming when presenting stimuli in a no-onset fashion. Even though we listed arguments that make an alternative interpretation of our results solely based on segregation or visual masking rather unlikely, these arguments provide only indirect support for excluding a masking account of our results. The current data from this study cannot directly disregard masking with all confidence. Thus, subsequent research will be needed to clarify this issue.
Conclusion
The current study provided further evidence for the flexibility and adaptability of the negative priming mechanism. By manipulating the deployment of visual attention to the stimuli, we demonstrated that this inhibitory negative priming mechanism can be either held active or disengaged according to specific task demands. Presenting prime and probe trial stimuli as no-onsets (non-automatic control of visual attention) resulted in a disengagement of the inhibitory mechanism. By implementing an onset presentation (automatic control) for either prime or probe trial, the inhibitory mechanism was held active across all trials. Further research is needed in order to specify in more detail how disengagement due to a no-onset presentation of stimuli is brought about.
Notes
Note, even though no-onset targets are assumed to be processed efficiently in a no-onset condition with distractor presence, such a claim refers to the selection in a no-onset condition only. Hereby, the assumption of efficient no-onset target processing is not at odds with the observation that general RTs in no-onset trials are slowed compared with onset trials as seen in Experiments 1 and 2. A no-onset presentation can have several implications, which may account for overall slowed RTs. For example, no-onset trials can generally be experienced as more difficult. Also, the mask itself could be responsible for prolonged RTs in Experiment 2 (e.g., longer stimulus identification times)
We thank an anonymous reviewer for suggesting this possibility.
We thank an anonymous reviewer for offering this suggestion.
We thank an anonymous reviewer for bringing up this point.
References
Awh, E., Matsukura, M., & Serences, J. T. (2003). Top-down control over biased competition during covert spatial orientation. Journal of Experimental Psychology: Human Perception and Performance, 29, 52–63.
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485–496.
Behringer, J. (2000). Experimental run system. Frankfurt am Main, Germany: BeriSoft.
Broadbent, D. E. (1958). Perception and communication. New York: Pergamon.
Buckolz, E., Boulougouris, A., O’Donnell, C., & Pratt, J. (2002a). Disengaging the negative priming mechanism in locations tasks. European Journal of Cognitive Psychology, 14,207–225.
Buckolz, E., Boulougouris, A., & Khan, M. (2002b). The influence of probe-trial election requirements on the location negative priming effect. Canadian Journal of Experimental Psychology, 56, 273–282.
Cave, K. R., & Wolfe, J. M. (1990). Modelling the role of parallel processing in visual search. Cognitive Psychology, 22, 225–271.
Cepeda, N. J., Cave, K. R., Bichot, N. P., & Kim, M. S. (1998). Spatial selection via feature-driven inhibition of distractor location. Perception & Psychophysics, 60, 727–746.
Christie, J., & Klein, R. M. (2001). Negative priming for spatial location? Canadian Journal of Experimental Psychology, 55, 24–38.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of selective visual attention. Annual Review of Neuroscience, 18, 193–222.
Di Lollo, V. (1980). Temporal integration in vision. Journal of Experimental Psychology: General, 109, 75–97.
Donk, M., & Theeuwes, J. (2001). Visual marking beside the mark: Prioritizing selection by abrupt onsets. Perception and Psychophysics, 63, 891–900.
Donk, M., & Theeuwes, J. (2003). Prioritizing selection of new elements: Bottom-up versus top-down control. Perception and Psychophysics, 65, 1231–1242.
Downing, P. E. (2000). Interactions between visual working memory and selective attention. Psychological Science, 11, 467–473.
Folk, C. L., & Remington, R. W. (1999). Can new objects override attentional control settings? Perception and Psychophysics, 61, 727–739.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044.
Fox, E. (1994). Interference and negative priming from ignored distractors: The role of selection difficulty. Perception and Psychophysics, 56, 565–574.
Fox, E. (1995). Negative priming from ignored distractors in visual selection: A review. Psychonomic Bulletin and Review, 2, 145–173.
Gibson, B. S. (1996). Visual quality and attentional capture: A challenge to the special role of abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 22, 1496–1504.
Hommel, B. (2000). Intentional control of automatic stimulus-response translation. In Y. Rossetti & A. Revonsuo (Eds.), Beyond dissociation: Interaction between dissociated implicit and explicit processing (pp. 221–242). Amsterdam: Benjamins.
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. Carr (Eds.), Inhibitory mechanisms in attention, memory and language (pp. 53–113). Orlando, FL: Academic.
Jonides, J. (1981). Voluntary vs. automatic control over the mind’s eye’s movement. In J. B. Long & A. D. Baddeley (Eds.), Attention and performance IX (pp. 187–203). Hillsdale, NJ: Erlbaum.
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception and Psychophysics, 43, 346–354.
Kane, M. J., May, C. P., Hasher, L., Rahal, T., & Stolzfus, E. R. (1997). Dual mechanisms of negative priming. Journal of Experimental Psychology: Human Perception and Performance, 23, 637–650.
MacDonald, P. A., Joordens, S., Seergobin, K. N. (1999). Negative priming effects that are bigger than a breadbox: Attention to distractor does not eliminate negative priming, it enhances it. Memory and Cognition, 27, 197–207.
Meiran, N. (1996). Reconfiguration of processing mode prior to task performance. Journal of Experimental Psychology: Learning, Memory and Cognition, 22, 1423–1442.
Milliken, B., Joordens, S., Merikle, P. M., & Seiffert, A. E. (1998). Selective attention: A reevaluation of the implications of negative priming. Psychological Review, 105, 203–229.
Milliken, B., Tipper, S., Houghton, G., & Lupianez, J. (2000). Attending, ignoring and repetition: On the relation between negative priming and inhibition of return. Perception and Psychophysics, 62, 1280–1296.
Müller, H., & Rabbitt, P. M. A. (1989). Reflexive and voluntary orienting of visual attention: Time course of activation and resistance to interruption.Journal of Experimental Psychology: Human Perception and Performance, 15, 315–330.
Neill, W. T., & Valdes, L. A. (1992). Persistence of negative priming: Steady state or decay? Journal of Experimental Psychology: Learning, Memory, and Cognition, 18, 565–576.
Neill, W. T., Valdes, L. A., Terry, K. M., & Gorfein, D. S. (1992). Persistence of negative priming. II. Evidence for episodic trace retrieval. Journal of Experimental Psychology: Learning, Memory and Cognition, 18, 993–1000.
Neill, W. T., Terry, K. M., & Valdes, L. A. (1994). Negative priming without probe selection. Psychonomic Bulletin and Review, 1, 119–121.
Neill, W. T., Valdes, L. A., & Terry, K. M. (1995). Selective attention and the inhibitory control of cognition. In F. N. Dempster, & C. J. Brainerd (Eds.), Interference and inhibition in cognition (pp. 207–261). New York: Academic.
Pratt, J., & Hirshhorn, M. (2003). Examining the time course of facilitation and inhibition with simultaneous onset and offset cues. Psychological Research, 67, 261–265.
Pratt, J., & Hommel, B. (2003). Symbolic control of visual attention: The role of working memory and attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 29, 835–845.
Pratt J., & McAuliffe J. (2001) The effects of onsets and offsets on visual attention. Psychological Research,65, 185–191.
Reder, L. M., Weber, K., Shang, J., & Vanyukov, P. M. (2003). The adaptive character of the attentional system: Statistical sensitivity in a target localization task. Journal of Experimental Psychology: Human Perception and Performance, 29, 631–649.
Richards, A. (1999). The effects of cuing target location and response mode on interference and negative priming using a visual selection paradigm. Quarterly Journal of Experimental Psychology, 52A, 449–463.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictable switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124, 207–231.
Samuel, A. G., & Weiner, S. K. (2001). Attentional consequences of object appearance and disappearance. Journal of Experimental Psychology: Human Perception and Performance, 27, 1433–1451.
Sohn, M.-H., & Carlson, R. A. (2000). Effects of repetition and foreknowledge in task-set reconfiguration. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 1445–1460.
Stablum, F., Ricci, R., Pavgese, A., & Umiltá, C. (2001). Global/local processing and negative priming: The influence of selection difficulty and stimulus exposure. Psychological Research, 65, 81–97.
Theeuwes, J. (1992). Perceptual selectivity for color and shape. Perception and Psychophysics, 51, 599–606.
Theeuwes, J., Godijn, R., & Pratt, J. (2004). A new estimation of the duration of attentional dwell time. Psychonomic Bulletin and Review, 11, 60–64.
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology: Human Experimental Psychology, 37A, 571–590.
Tipper, S. P. (2001). Does negative priming reflect inhibitory mechanisms? A review and integration of conflicting views. Quarterly Journal of Experimental Psychology, 54A, 321–343.
Tipper, S. P., & Cranston, M. (1985). Selective attention and priming: Inhibitory and facilitatory effects of ignored primes. Quarterly Journal of Experimental Psychology, 37A, 591–611.
Tipper, S. P., Weaver, B., Cameron, S., Brehaut, J. C., & Bastedo J. (1991). Inhibitory mechanisms of attention in identification and localization tasks: Time course and disruption. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17, 681–692.
Tipper, S. P., Weaver, B., & Houghton, G. (1994). Behavioral goals determine inhibitory mechanisms of selective attention. Quarterly Journal of Experimental Psychology, 47A, 809–840.
Todd, J. T., & Van Gelder, P. (1979). Implications of a transient-sustained dichotomy for the measurement of human performance. Journal of Experimental Psychology: Human Perception and Performance, 5, 625–636.
Woodman, G. F., & Luck, S. J. (1999). Electrophysiological measurement of rapid shifts of attention during visual search. Nature, 400, 867–869.
Yantis, S. (1998). Control of visual attention. In H. Pashler (Ed.), Attention (pp. 223–256). Hove, UK: Psychology.
Yantis, S., & Hillstrom, A. P. (1994). Stimulus-driven attentional capture: Evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception and Performance, 20, 95–107.
Yantis, S., & Johnson, D. N. (1990). Mechanisms of attentional priority. Journal of Experimental Psychology: Human Perception and Performance, 16, 812–825.
Yantis, S., & Jones, E. (1991). Mechanisms of attentional selection: Temporally modulated priority tags. Perception and Psychophysics, 50, 166–178.
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10, 601–621.
Yantis, S., & Jonides, J. (1990). Abrupt visual onsets and selective attention: Voluntary versus automatic allocation. Journal of Experimental Psychology: Human Perception and Performance, 16, 121–134.
Yee, P. L., Santoro, K. E., Grey, A. L., & Woog, V. (2000). Deeper processing at target selection increases the magnitude of negative priming. Memory and Cognition, 28, 1429–1436.
Acknowledgements
The authors would like to thank two anonymous reviewers for their helpful comments, and Peter Frensch, Sven Heusner, Claudia Preuschhof, and Paul McCormick for valuable discussions and for improving the English.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Fischer, R., Hagendorf, H. The control of visual attention and its influence on prioritized processing in a location negative priming paradigm. Psychological Research 70, 317–335 (2006). https://doi.org/10.1007/s00426-005-0220-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-005-0220-2