
Abstract
The experiment conducted examined the effect of simultaneously presented onset and offset cues on the orienting of attention in the visual field. Subjects were presented with a display that consisted of four placeholder boxes around a central fixation point. An onset and an offset cue appeared simultaneously in two of the locations, and the other two locations provided a neutral baseline condition. Reaction times were measured in a simple target detection task with stimulus-onset asynchronies (SOAs) that ranged from 100 ms to 1,000 ms. As expected, the onset cue produced early facilitation and later occurring inhibition of return (IOR). The offset cue produced significant inhibition at all but the earliest SOA. These results suggest that simultaneously presented onset and offset cues both capture attention, but that attention is rapidly disengaged from the location of the offset cue, resulting in earlier occurring IOR. For the onset cues, attention is allocated for a longer period of time, producing the typical pattern of early facilitation and later occurring IOR. The differing time course of attention at each location may reflect separate facilitatory and inhibitory processes, and the priority given to the onset of a stimulus by the attentional system.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In the cluttered and dynamic visual environments that we interact with on a daily basis, there is a continuous flow of new objects appearing and old object disappearing in our visual field. The importance of reflexively orienting attention to abruptly appearing new objects seems obvious; information about the size, speed, and trajectory of the object must be obtained in order to determine whether some action must be performed in response to the object. The case for reflexively orienting to abruptly disappearing objects is not as clear-cut. After all, how often is there an advantage to orienting attention to a location where an object no longer exists? Perhaps because of this intuitive difference between these two types of event, researchers have almost exclusively used abrupt peripheral onsets to examine the reflexive (or exogenous) orienting system (e.g., Posner, 1980; Posner & Cohen, 1984; Yantis & Jonides, 1984; with a great many more examples possible). On the other hand, studies examining the attentional consequences of the sudden offset of an object can be counted on one hand (Gawryszewski, Thomaz, Machado-Pinheiro, & Sant'Anna, 1994; Riggio, Bello, & Umiltà, 1998; Pratt and McAuliffe, 2001; Samuel & Weiner, 2001). Furthermore, the vast majority of both onset and offset studies have examined situations in which a single peripheral event occurred (whether it be the abrupt appearance of a new object or the abrupt disappearance of an old object). While these studies suggest that onsets and offsets may have differential effects on the allocation of visual attention, it is also the case that objects in the real world do not always appear or disappear one at a time. To gain some understanding of how attention is oriented in the visual field when multiple different peripheral events occur, the present experiment examined simultaneous onset and offset events across a series of stimulus-onset asynchronies (SOAs).
Before examining how the visual attention system deals with simultaneous onset and offset events, some background regarding what is known with solitary onsets and offsets is necessary. As noted earlier, there is a substantial body of literature concerning the effect of a single onset event (typically called a "cue") on the allocation of attention. The study most germane to the present work was that conducted by Posner and Cohen (1984). The crux of this study was the examination of the effect of peripheral onset cue on the time to detect a subsequent peripheral target. Replicating earlier work (e.g., Posner, 1980), they found that targets at cued locations were detected faster than targets at uncued locations when the SOA was less than 300 ms. Unexpectedly, they also found that detection responses were slower for targets at the cued locations when the SOA was greater than 300 ms. The inhibitory effect has become known as inhibition of return (IOR) and has proven to be a very robust effect (e.g., Abrams & Dobkin, 1994; Maylor, 1985; Pratt, 1995; Tipper, Driver, & Weaver, 1991). Moreover, the biphasic pattern of early facilitation followed by late inhibition at the cued location has been replicated (e.g., Berger, Dori, & Henik, 1999; Rafal, Calabresi, Brennan, & Sciolto, 1989) and taken by some as the benchmark of attentional orienting (e.g., Taylor & Klein, 1998). It is worth noting that although the late IOR effect appears to be very reliable, the same is not true of early facilitation effects (e.g., Pratt & McAuliffe, 2002; Tassinari, Agliotti, Chelazzi, Peru, & Berlucchi, 1994; Tassinari & Berlucchi, 1993). Recent evidence suggests that the unreliable nature of early facilitation effects is likely due to the temporal and spatial parameters of the cues (e.g., Maruff, Yucel, Danckert, Stuart, & Currie, 1999; Pratt, Hillis, & Gold, 2001).
Also, as noted earlier, there has been considerably less work examining the effect of offset cues on the allocation of attention in the visual field. Using a modification of the Posner and Cohen (1984) procedure, and offset cues and SOAs of 100, 200, 300, 500, and 800 ms, Gawryszewski, Thomaz, Machado-Pinheiro, and Sant'Anna (1994) found inhibitory effects at both the short and the long SOAs. A similar pattern of results, but with use of much more complex displays, was found by Samuel and Weiner (2001). Across three experiments, Riggio, Bello, and Umiltà (1998) found varying early facilitation effects but strong late inhibitory effects for both solitary onset and solitary offset cues. These findings led to a range of conclusions. On the one hand, Gawryszewski et al. and Riggio et al. suggested that their findings provide evidence that facilitatory and inhibitory effects are independent of each other, although both presumably rely on attention orienting. On the other hand, Samuel and Wiener suggested that there may be separate inhibitory mechanisms for onsets and offsets, with offsets generating more voluntary attention responses and onsets generating more reflexive responses.
Although it is clear that both onsets and offsets, in isolation, can produce shifts of attention, it is not clear how attention is allocated when the two types of event occur simultaneously. The only study to address this issue (that we are aware of) was done by Pratt and McAuliffe (2001). Using a Posner and Cohen (1984) type of procedure of display with two peripheral cue/target locations, they presented either a solitary peripheral onset cue, or a solitary peripheral offset cue, or simultaneous peripheral onset and offset cues at short (100 ms) and long (900 ms) SOAs. Both types of solitary event produced early facilitation and late inhibition effects. In trials in which the two events occurred simultaneously (the onset in one peripheral location, the offset in the other), faster responses were found to targets at the onset location at the 100-ms SOA, but no differences between cue locations was found at the 900-ms SOA. Pratt and McAuliffe concluded that although both types of cue produce approximately equal IOR effects when presented simultaneously, onset cues get priority in attentional orienting at short SOAs when the task involves localizing targets in space.
There are some reasons, however, to suggest that the method used to examine simultaneous onsets and offsets by Pratt and McAuliffe (2001) may not have provided the most complete picture possible. First, and most critical, is that there were only two possible cue/target locations, requiring both of the locations to be cued in the simultaneous onset-offset trials. Thus, there was no baseline (i.e., uncued) condition against which the facilitatory and inhibitory effects could be compared. In finding no differences in responses to onset cued and offset cued trials at the long SOA, Pratt and McAuliffe suggested that both cues produced a roughly equal amount of IOR. But the determination of IOR involves the comparison of reaction times (RTs) from cued location to uncued locations, and there were no uncued locations in the onset-offset condition of Pratt and McAuliffe. Rather, because both cue types in isolation produced IOR, Pratt and McAuliffe assumed that equivalent RTs in the simultaneous cue condition were due to production of similar IOR effects by each of the simultaneous cues. Second, performance may be different when all possible target locations are cued than when an uncued location remains in the visual field (for an example of this argument, see Tipper, Weaver, & Watson, 1996). Third, Pratt and McAuliffe only used two SOAs, one very short (100 ms) and one very long (900 ms), which provided relatively little information about the time course of the effects at the two simultaneously cued locations.
The present study was designed to provide a more definitive examination of the effect of simultaneous onset and offset cues on the allocation of attention, with the aim of reaching conclusions lesstentative than those allowed by the Pratt and McAuliffe (2001) study. To provide the critical uncued baseline condition against which the effects of onset and offset cues could be compared, four possible cue/target locations were used. Thus, on every trial, four placeholders appeared, with a simultaneous onset and offset occurring in two of them. This also means that not every possible target location was cued, eliminating that as a potential confound. Moreover, to allow for a thorough understanding of the time course of the cue effects, five SOAs (100, 250, 500, 750, or 1,000 ms) were used. As in the previous studies that investigated offset events, simple detection responses were measured.
Method
Participants
Eighteen undergraduate students from the University of Toronto participated in the experiment in return for course credits. All were naïve to the purposes of the experiment.
Apparatus and procedure
The experiment took place in a dimly lit, sound-attenuated room. Participants were seated 44 cm in front of a computer monitor. A head/chin rest was used to keep the viewing distance constant. The computer keyboard was used as the response device and was kept within comfortable reach of the participant.
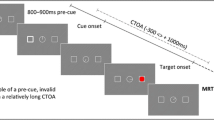
The trial sequence for the experiment is shown in Fig. 1. The initial display of the trial sequence was presented for 1,000 ms. In each condition the initial display consisted of four placeholder boxes, each centered 4.5° from a center fixation point. Each box was 1° wide and tall, and appeared in white (31.5 cd/m2) on a black background (0.5 cm/m2). Two of the boxes in the initial display were filled with white circles (0.8° in diameter). Following the initial display an onset cue was presented in one of the empty placeholders (a white circle identical to the two that appeared in the initial display). At the same time, an offset cue was presented by removing one of the initial two white circles. The target appeared following SOAs of 100, 250, 500, 750, or 1,000 ms. The target was the white outline of a larger box (1.2°) around one of the placeholder boxes. Participants pressed the spacebar as quickly as possible after target detection and the target stayed on until the response was made. If participants responded faster than 100 ms or slower than 1,000 ms, the response was considered an error.

The basic trial sequence used in the experiment. In any given trial, the target was equally likely to appear in the onset, the offset, or either of the two uncued locations
Design
Each participant completed 480 trials, 20% of which were catch trials in which no target appeared after the simultaneous onset and offset cues. Onset and offset cues were equally likely to occur in either of the two filled boxes (offsets) and the two empty boxes (onsets). The target was equally likely to occur at all four locations.
Results
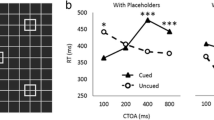
The mean RTs from the correct trials are presented in Fig. 2. The mean RTs were analyzed with a 3 (trial type: onset, offset, or uncued) × 5 (SOA: 100, 250, 500, 750, or 1,000) analysis of variance (ANOVA). There was a main effect for SOA [F(4, 68)=5.9, MSE=1,272, p<.0007]. Participants were the slowest in responding to the 100-ms SOA (448 ms), fastest at the 250-ms SOA (416 ms), and then increasingly slower as the SOA was increased up to 1,000 (427 ms, 426 ms, 435 ms). The main effect for trial type was not significant [F(2, 34)<2.7, p>.14]. Most importantly, there was a significant two-way interaction between SOA and trial type [F(8, 136)=2.56, MSE=404, p<.015], indicating that the three types of cue had different effects at the various SOAs. Planned comparisons were done to better understand the interaction between SOA and trial type. At the 100-ms SOA the onset cue produced significant facilitation (p<.002), whereas the inhibition produced by the offset cue was not significant (p>.70). At the 250-ms SOA the onset cue produced significant facilitation (p<.03) whereas the offset cue produced significant inhibition (p<.05). The offset cue continued to produce significant inhibition at the 500-ms SOA (p<.04). RTs were also slower at the onset location than the uncued location at the 500-ms SOA, but the inhibition was not significant (p>0.15). Both the onset cue and the offset cue produced significant inhibition at both the 750-ms and the 1,000-ms SOAs (ps<0.05).

Mean RTs from the correct trials
The error rates are shown in Table 1 and were analyzed with a 3 (trial type: onset, offset, or uncued) × 5 (SOA: 100, 250, 500, 750, or 1,000) ANOVA. None of the main effects, nor any interaction effects, reached significance (ps>.15).
General discussion
This study was designed to expand on the work done by Pratt and McAuliffe (2001) in examining the effect of simultaneously presented onset and offset cues on the allocation of visual attention. As expected, the onset cues produced early facilitation at the 100-ms and 250-ms SOAs, followed by inhibition at the 750-ms and 1,000-ms SOAs. Of more interest, the offset cues did not have a significantly different effect from the uncued locations at the shortest SOA and then produced inhibition at all of the remaining SOAs.
The biphasic pattern of RTs observed at the location of the onset cue is exactly what one would expect based on previous work (e.g., Posner & Cohen, 1984). Moreover, the present findings indicate that an onset cue is capable of capturing attention even when an offset cue is presented simultaneously at a different location. This is consistent with the long-held notion that onset cues initially capture and hold attention briefly (producing the short-duration facilitation effect) and then IOR is generated at the cued location when attention is disengaged and moved to another location (producing the long-duration inhibitory effect). The fact that this happens even when there is a simultaneous offset event in the visual field supports the notion that the appearance of new objects tends to reflexively cause the orienting of attention to that location (e.g., Yantis & Hillstrom, 1994).
The pattern of RTs produced by the offset cue does not match the pattern of the onset cues. Rather, after producing no facilitatory effect, the offset cues produced inhibitory effects at all but the shortest SOAs. Why would the offset cues, which consisted in the same change of visual information as the onset cues, produce such a dramatically different time course of RTs? It is unlikely that the lack of spatial overlap between cues and targets with the offset cues was the cause as various researchers have shown early facilitation without such spatial overlap (e.g., Berger, Dori, & Henik, 1999; Pratt, Hillis, & Gold, 2001). Our suggestion is that this reflects the fact that facilitation and IOR are two separate phenomena. Evidence in support of this idea was found in a previous study by Collie, Maruff, Yucel, Dancket, and Currie (2000) that manipulated the spatial and temporal relationships between cue and target in a simple RT task. Collie et al. (2000) suggested that facilitation occurs as a result of an attentional shift to the peripheral cue, whereas IOR may arise as a motor bias against responding to stimuli in the cued hemifield. Interestingly, the study also reported that IOR may require longer than 150 ms to take effect, which explains why IOR was not observed at the shortest SOA in the offset condition in this experiment.
Also supporting the separate process notion is work by Danzinger and Kingstone (1999) that showed that attention directed to a cued location can mask the presence of early occurring IOR at the same location. When an onset and an offset cue occur simultaneously, attention is directed to the location of the onset cue, unmasking the presence of IOR at the location of the offset cue. This suggests that there may be an advantage in directing attention towards the location of an onset cue, which may reflect the priority given to the appearance of new objects by the visual attention system. Attending to a new object allows us to evaluate its properties and then act accordingly. It is equally important for the disappearance of an object to capture our attention; however, the lack of new information to evaluate may lessen the priority of offsets in the visual attention system. Thus, treating facilitation and IOR as two separable processes provides a good account for the different time course of RTs observed in response to onset and offset cues.
References
Abrams, R. A. & Dobkin, R. S. (1994). The gap effect and inhibition of return: interactive effect on eye movements latencies. Experimental Brain Research, 98, 483–487.
Berger, A. Dori, H., & Henik, A. (1999). Peripheral non-informative cues do induce early facilitation of target detection. European Journal of Cognitive Psychology, 11, 119–137.
Collie, A., Maruff, P., Yucel, M., Dancket, J., & Currie, J. (2000). Spatiotemporal distribution of facilitation and inhibition of return arising from the reflexive orienting of covert attention. Journal of Experimental Psychology: Human Perception and Performance, 26, 1733–1745.
Danziger, S., & Kingstone, A. (1999). Unmasking the inhibition of return phenomenon. Perception & Psychophysics, 61, 1024–1037.
Di Lollo, V., Enns, J. T., Yantis, S., & Decheif, L. G. (2000). Response latencies to the onset and offset of visual stimuli. Perception & Psychophysics, 62, 218–225.
Gawryszewski, L. G., Thomaz, T. G., Machado-Pinheiro, W., & Sant'Anna, A. N. (1994). Onset and offset of a visual cue have different effects on manual reaction time to a visual target. Brazilian Journal of Medical Biology, 27, 67–73.
Martin-Emerson, R. & Kramer, A. F. (1997). Offset transients modulate attentional capture by sudden onsets. Perception & Psychophysics, 59, 739–751.
Maruff, P., Yucel, M., Danckert, J., Stuart, G., & Currie, J. (1999). Facilitation and inhibition arising from the exogenous orienting of covert attention depends on the temporal properties of spatial cues and targets. Neuropsychologia, 37, 731–744.
Maylor, E. (1985). Facilitatory and inhibitory components of orienting in visual space. In M. I. Posner & B. B. Marin (Eds.), Attention and Performance XI (pp 189–304). Hillsdale, N.J.: Erlbaum.
Miller, J. (1989). The control of attention by abrupt visual onsets and offsets. Perception & Psychophysics, 45, 567–571.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D.G. Bouwhuis (Eds.), Attention and Performance X (pp 531–556). Hillsdale, N.J.: Erlbaum.
Pratt, J. (1995). Inhibition of return in a discrimination task. Psychonomic Bulletin & Review, 2, 117–120.
Pratt, J., & McAuliffe, J. (2001). The effects of onsets and offsets on visual attention. Psychological Research, 65, 185–191.
Pratt, J. & McAuliffe, J. (2002). Determining if attentional control settings are inclusive or exclusive. Perception & Psychophysics 64, 1361–1370.
Pratt, J., Hillis, J., & Gold, J. M. (2001). The effect of the physical characteristics of cues and targets on facilitation and inhibition. Psychonomic Bulletin & Review, 8, 489–495.
Rafal, R. D., Calabresi, P. A., Brennan, C. W., & Sciolto, T. K. (1989). Saccade preparation inhibits reorienting to recently attended locations. Journal of Experimental Psychology: Human Perception and Performance, 15, 673–685.
Riggio, L., Bello, A., & Umiltà, C. (1998). Inhibitory and facilitatory effects of cue onset and offset. Psychological Research, 61, 104–118.
Samuel, A. G. & Weiner, S. K. (2001). Attentional consequences of object appearance and disappearance. Journal of Experimental Psychology: Human Perception and Performance, 27, 1433–1451.
Tassinari, G., & Berlucchi, G. (1993). Sensory and attentional components of slowing of manual reaction time to non-fixated visual targets by ipsilateral primes. Vision Research, 33, 1525–1534.
Tassinari, G., Agliotti, S., Chelazzi, L., Peru, A., & Berlucchi, G. (1994). Do peripheral non-informative cues induce early facilitation of target detection. Vision Research, 34, 179–189.
Taylor, T. L. & Klein, R. M. (1998). Inhibition and return to color: a replication and nonextenison of Law, Pratt, and Abrams (1995). Perception & Psychophysics, 60, 1452–1456.
Tipper, S. P, Driver, J., & Weaver, B. (1991). Object-centered inhibition of return of visual attention. Quarterly Journal of Experimental Psychology: Human Experimental Psychology, 43A, 289–298.
Tipper, S. P, Weaver, B., & Watson, F. (1996). Inhibition of return to successively cued spatial locations: a commentary on Pratt and Abrams (1995). Journal of Experimental Psychology: Human Perception and Performance, 22, 1289–1293.
Yantis, S., & Hillstrom, A. P. (1994). Stimulus-driven attentional capture: evidence from equiluminant visual objects. Journal of Experimental Psychology: Human Perception and Performance, 20, 95–107.
Yantis, S. & Jonides, J. (1984). Abrupt visual onsets and selective attention: evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10, 601–621.
Acknowledgements
This research was supported by a grant from the Natural Sciences and Engineering Council of Canada to Jay Pratt.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pratt, J., Hirshhorn, M. Examining the time course of facilitation and inhibition with simultaneous onset and offset cues. Psychological Research 67, 261–265 (2003). https://doi.org/10.1007/s00426-003-0130-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-003-0130-0