
Abstract
The aim of the present study was to investigate the acute effect of caffeine or quercetin ingestion on motor unit firing patterns and muscle contractile properties before and after resistance exercise. High-density surface electromyography (HDs-EMG) during submaximal contractions and electrically elicited torque in knee extensor muscles were measured before (PRE) and 60 min after (POST1) ingestion of caffeine, quercetin glycosides, or placebo, and after resistance exercise (POST2) in ten young males. The Convolution Kernel Compensation technique was used to identify individual motor units of the vastus lateralis muscle for the recorded HDs-EMG. Ingestion of caffeine or quercetin induced significantly greater decreases in recruitment thresholds (RTs) from PRE to POST1 compared with placebo (placebo: 94.8 ± 9.7%, caffeine: 84.5 ± 16.2%, quercetin: 91.9 ± 36.7%), and there were significant negative correlations between the change in RTs (POST1-PRE) and RT at PRE for caffeine (rs = − 0.448, p < 0.001) and quercetin (rs = − 0.415, p = 0.003), but not placebo (rs = − 0.109, p = 0.440). Significant positive correlations between the change in firing rates (POST2-POST1) and RT at PRE were noted with placebo (rs = 0.380, p = 0.005) and quercetin (rs = 0.382, p = 0.007), but not caffeine (rs = 0.069, p = 0.606). No significant differences were observed in electrically elicited torque among the three conditions. These results suggest that caffeine or quercetin ingestion alters motor unit firing patterns after resistance exercise in different threshold-dependent manners in males.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Caffeine is one of the most commonly consumed ergogenic aids in the world (Graham 2001). Because of the physiological effect of caffeine on exercise performance, its use is prevalent among athletes (Grgic et al. 2020). Quercetin is also gaining attention as an ergogenic aid with similar effects to caffeine. Caffeine and quercetin are structurally similar to adenosine; they block the binding of adenosine to A1 and A2a receptors and release of neurotransmitters such as acetylcholine and dopamine (Alexander 2006; Graham 2001; McLellan et al. 2016; Grgic et al. 2019). Moreover, both caffeine and quercetin increase calcium release from the sarcoplasmic reticulum within skeletal muscles (Graham 2001; Lee et al. 2002; McLellan et al. 2016; Grgic et al. 2019). Thus, quercetin may be applicable as a potential ergogenic aid to alter arousal, contraction velocity, and exerted tension of muscle fibers and improve physical performance in addition to caffeine.
Ingestions of caffeine and quercetin have been shown to change motor unit firing patterns and muscle contractile properties (Alexander 2006; Graham 2001; McLellan et al. 2016; Grgic et al. 2019; Lee et al. 2002; Cheuvront et al. 2009). Our previous study revealed that quercetin ingestion could enhance activations of motor units with relatively higher recruitment thresholds (RTs), evaluated using high-density surface electromyography (HDs-EMG) (Watanabe and Holobar 2021). Because of the structural similarities between caffeine and quercetin (Alexander 2006), caffeine ingestion may also change the motor unit firing pattern in the same threshold-dependent manner as quercetin ingestion. Changes in neuromuscular conditioning induced by caffeine or quercetin ingestion may alter motor unit firing during subsequent muscle exertion exercises (e.g., resistance exercise), and may alter the state of peripheral fatigue. Motor neurons with relatively higher and lower RT normally innervate muscle fibers that contribute to higher and lower forces, respectively (Henneman and Olson 1965; Henneman et al. 1965; Burke et al. 1973). Changes in muscle contractile properties could modify neuromuscular efficiency, and their different behaviors may be reasonable when considering different changes in peripheral components of the neuromuscular system. When caffeine or quercetin ingestion activates a higher motor unit firing pattern, the muscle fibers that contribute to the greater torques may show decreased contractile performance because of fatigue. However, it is unclear how caffeine or quercetin ingestion changes motor unit firing patterns and muscle contractile properties before and after resistance exercise.
This study aimed to investigate the acute effects of caffeine (200 mg) and quercetin glycoside (500 mg) ingestions on motor unit firing patterns and muscle contractile properties during isometric contractions before and after resistance exercise. We hypothesized that ingestions of both caffeine and quercetin would induce: (1) a decrease in RT of higher threshold motor units after ingestion, (2) an increase in the firing rate of higher threshold motor units after resistance exercise, and (3) a decrease in contraction torques during higher frequency electric stimulation.
Materials and methods
Participants
Prior to conducting data collection, we calculated the minimum required sample size using G*power (Dusseldorf, Germany) based on the analysis of motor units in our previous study (Watanabe and Holobar 2021). This calculation assumed an effect size of 0.40, alpha of 0.05, and a power of 0.80. This analysis indicated that a sample size of 54 motor units was necessary. In our previous study (Watanabe and Holobar 2021), we observed an average of 7 motor units per participant, leading us to recruit a total of 10 participants after considering the potential for dropouts. Ten healthy young males (mean ± SD: age: 24.1 ± 3.0 years, height: 172.7 ± 2.6 cm, body weight: 64.1 ± 5.8 kg) who did not habitually perform resistance exercises participated in this study. Those who had any exercise restrictions imposed by medical doctors were excluded from the study. The participants gave written informed consent for the study after receiving a detailed explanation of the purposes, potential benefits, and risks associated with participation. All procedures used in this study were approved by the Research Ethics Committee of Chukyo University (2019-003).
Experimental design
Participants came to the laboratory on 3 days separated by 72 h and were subjected to the three ingestion conditions (caffeine, quercetin, and placebo capsules) in a randomly determined order. Based on previous research indicating that plasma caffeine levels return to the baseline approximately 29 h after the ingestion of a 200-mg caffeine capsule (Kamimori et al. 2002) and that plasma quercetin levels return to the baseline approximately 24 h after the ingestion of 500 mg of quercetin (Davis et al. 2009), experimental days were separated by 72 h. They were asked to refrain from vigorous-intensity exercise, the consumption of foods or drinks containing quercetin or caffeine (e.g., coffee, tea, energy drinks), and the ingestion of other possible ergogenic aids sold as functional foods (e.g., supplement, government-approved health food) 24 h before testing.
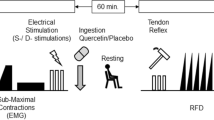
The protocols are shown in Fig. 1. After sitting on a dynamometer, participants’ hip and knee joint angles were fixed at 90°, and the distal part of the right shank was fixed to the dynamometer with a force transducer (LU-100KSE; Kyowa Electronic Instruments, Tokyo, Japan) to measure the force exerted by knee extensor muscles. Knee extension torque was calculated by multiplying the force and arm length. The arm length was measured between the knee joint axis and force transducer.

Schematic overview of the experimental protocol. MVC, maximal voluntary contraction; EMG, electromyography; EET, electrically elicited torque, PRE, before ingestion; POST1, 60 min after ingestion; POST2, after resistance exercise
After standardized warm-up protocols (submaximal isometric contraction of 2 × 50%, 2 × 80%), maximum voluntary contraction (MVC) of isometric knee extension was measured. Participants performed 2–3 MVCs and rest for more than 30 s between efforts. The peak during 5 s MVC was recorded, and the greatest value between the efforts was used for further analysis. Participants also performed submaximal ramp contractions for recording HDs-EMG to identify individual motor unit activation in the vastus lateral (VL) muscle. Electrically elicited knee extensor torque was employed to quantify the contractile properties of muscle (See Electrically elicited knee extensor torque).
After the measurements (PRE), the participants took six capsules containing 200 mg of caffeine with 2.0–2.5 g of dextrin, 500 mg of quercetin glycosides with 2.0–2.5 g of dextrin, or only 2.0–2.5 g of dextrin (placebo) with 500 mL of water. Quercetin glycosides are more water-soluble and bioavailable than quercetin aglycone, which does not exist in a glycosidic or conjugated form (Makino et al. 2009). When absorbed, quercetin glycorandomsides are enzymatically converted into the aglycone form and exhibit beneficial effects similar to those of the corresponding quercetin aglycone. Quercetin glycosides were enzymatically manufactured at San-Ei Gen F.F.I., Inc. (Osaka, Japan) from isoquercitrin prepared from quercetin-3-O-rutinoside. In the Generally Recognized As Safe statement from the Food and Drug Administration, up to 200 mg/serving of caffeine and 500 mg/serving of quercetin are acceptable. The capsules for caffeine, quercetin glycosides, and placebo had the same shape, color, and weight. A randomized, double-blind, placebo-controlled treatment was administered in this study. After ingestion of the capsules, the participants remained seated for 60 min to allow for sufficient elevation of plasma caffeine and quercetin concentrations. Post-ingestion measurements (POST1) were then conducted (Graham 2001; Burak et al. 2017). After POST1 measurements, participants performed isometric knee extension exercises that simulated resistance exercises. After POST1 measurement and 2 min rest, participants completed three sets of 10 repetitions of 70%MVC isometric knee extension with 2 min rest between each set. Each set is constructed by repeating 5 s contraction and 5 s rest. Participants were instructed to perform knee extension in response to a visual target displayed on a monitor in front of them. Following the completion of the exercise, a post-exercise measurement (POST2) was performed immediately and was completed with 8-m. We used the same electrodes for recording surface EMG and applying electrical stimulation between PRE, POST1, and POST2; thus, we did not replace them for measurements taken on the same day.
In order to minimize variations in preparation and assessment time among participants, the experiment was executed in accordance with a pre-established time-line, derived from a preliminary experiment (e.g., 5 min elapsed from participant's seating on the dynamometer to the initiation of the experiment; 8 min interval between measurements of MVC and electrically elicited torque, etc.).
Submaximal contractions and recording of high-density surface EMG
Figure 2 shows experimental set up for recording HDs-EMG and methods of calculating motor unit firing patterns. To record HDs-EMG signals, participants performed ramp contractions from 0 to 50%MVC during isometric knee extension. The device was used for MVC measurements and joint angles were also 90° during contractions. Ramp contractions consisted of a 17 s increasing phase from the baseline to 50%MVC with an approximately 3%MVC/sec rate of force increase and 10 s sustained phase at 50%MVC. Many previous studies have been used ramp contractions to measure motor unit activity condition (Watanabe and Holobar 2021; Kunugi et al. 2021; Del Vecchio et al. 2019a; Adam and De Luca 2005). During these submaximal contractions, the performed and target torques were shown on the monitor of a personal computer as visual feedback. The target torque in each measurement period was based on the MVC at PRE, because participants could exert the same absolute torque during submaximal contractions at PRE, POST1, and POST2 on the same day. At only PRE, participants performed ramp contractions from 0 to 30%MVC during isometric knee extension to facilitate the identification of motor unit filters that could increase identification accuracy of individual motor units in decomposition analysis.

Methods of high-density surface electromyography overview. a Experimental set up. b Recording of high-density surface electromyography and motor unit decomposition. c Analysis of individual motor units and their tracking processes. EMG, electromyography; MVC, maximal voluntary contraction
HDs-EMG signals were recorded from the VL muscle with a semi-disposable adhesive grid of 64 electrodes and a 1-mm diameter and 8-mm inter-electrode distance (GR08MM1305, OT Bioelettronica, Torino, Italy). The electrodes were organized in 13 rows and 5 columns with 1 missing electrode in the distal left corner. The midpoint of the line between the greater trochanter and superior lateral edge of the patella was used as the center of the electrode grid, and the line was also used to determine the direction of electrode grids, whereby columns of electrodes were aligned along the line. A reference electrode (WS2, OT Bioelettronica, Torino, Italy) was fixed to the proximal end of the shank. Monopolar surface EMG signals were recorded with a bandpass filter (10–500 Hz) and amplified by a factor of 256, sampled at 2000 Hz, and converted to digital form by a 16-bit analog-to-digital converter (Sessantaquattro, OT Bioelettronica, Torino, Italy). The signal from the force transducer was synchronized with the analog-to-digital converter.
Recorded monopolar surface EMG signals were transferred to analysis software (MATLAB R2019a, MathWorks GK, Tokyo, Japan), and individual motor units were identified by the Convolution Kernel Compensation technique using DEMUSE software (Holobar and Zazula 2004, 2008; Merletti et al. 2008; Holobar et al. 2009). We followed the decomposition procedure previously and extensively validated based on signals from various skeletal muscles (Holobar et al. 2009; Farina et al. 2010; Gallego et al. 2015a, 2015b; Yavuz et al. 2015; Watanabe et al. 2016, 2018; Watanabe and Holobar 2021). The pulse-to-noise ratio, introduced by Holobar et al. (2014), was used as an indicator of the motor unit identification accuracy, and motor units with pulse-to-noise ratio > 30 dB (corresponding to an accuracy of motor unit firing identification > 90%) were considered for further analysis (Holobar et al. 2014). After decomposition, firing timings of individual motor units were independently examined by an experienced investigator. Firing times for individual motor units were used to calculate instantaneous motor unit firing rates. To eliminate firing intervals uncharacteristically high (exceeding 30 Hz) or low (less than 4 Hz) for the VL muscle, we excluded firings with intervals shorter than 33.3 ms or longer than 250 ms. These procedures were the same as those used in our previous studies (Watanabe et al. 2016, 2020; Watanabe and Holobar 2021).
Detected motor units were tracked between PRE, POST1, and POST2 on the same day using the Convolution Kernel Compensation technique to calculate the motor unit identification filters from HDs-EMG signals at PRE and apply them to HDs-EMG signals at POST1 and POST2(Francic and Holobar 2021). Motor units were not tracked between condition for caffeine, quercetin and placebo. Therefore, our analysis of motor units was limited to those active in the PRE session only, but facilitated the repeated measures statistical comparison. Also, tracked motor units were assessed for their similarity to the action potential waveform shape by cross-correlation analysis. Previous studies demonstrated that the method employed enables highly efficient tracking of individual motor unit firing patterns across various conditions (Del Vecchio et al. 2019b; Kunugi et al. 2021). The previously introduced criterion of pulse-to-noise ratio > 30 dB was also applied to motor unit tracking, ensuring an accuracy of motor unit firing identification > 90% at POST1 and POST2. Motor unit firing rates with > 30% coefficient of variation were excluded from further analysis (Fuglevand et al. 1993). Motor units detected during a condition for caffeine, quercetin, or placebo were merged across all participants and their mean values were used for further analysis. RTs were determined by the torque value (%MVC) corresponding to the second motor unit firing after recruitment and computed from the series of motor unit firings identified by decomposition. The firing rates for that motor unit were determined using the median value of interspike intervals between motor unit spikes at 44 and 50%MVC. POST1:PRE, POST2:PRE, and POST2:POST1 were calculated as measures of the rate of change in motor unit activities after capsule ingestion and exercise.
Electrically elicited knee extensor torque
The effects of caffeine and quercetin ingestions on contraction velocity and tension exerted by muscle fibers were analyzed through the measurement of electrically induced torque at various stimulation frequencies. This approach was used in several previous studies to investigate the effects of supplementation on skeletal muscle contractile properties (Haider and Folland 2014; Coombes et al. 2001, 2002). Elicited torques of knee extensor muscles during electric stimulation were measured to estimate muscle contractile properties (Tomita et al. 2020). The quadriceps femoris muscle was stimulated with two self-adhesive electrodes (6 cm) using a stimulator device (DS7AH, Digitimer, Hertfordshire, UK). Proximal and distal quadriceps muscles were carefully palpated by an experienced investigator and electrodes were attached to the superficial quadriceps femoris (rectus femoris, VL, and vastus medialis). The electrode length could appropriately cover the quadriceps femoris and be attached to the electric insulation sheet to prevent stimulation to knee flexor. Prior to measurement, electrical stimulation was administered incrementally by increasing the current by 50 mA while observing the resultant knee extension torque. The torque exerted at the current level at which the torque plateaued was determined as the electrically elicited torque via twitch stimulation. Electrical stimulation was then performed at frequencies of 10, 20, 40, and 80 Hz at a current level of 30% of singlet stimulation (Tomita et al. 2020). The pulse duration was 200 μs during stimulation. Signals from the dynamometer were input to a computer running LabChart software at an analog–digital conversion rate of 2000 Hz (PowerLab; ADInstrument). The stimulation duration was < 10 s and the final 3 s of the evoked torque were selected for each frequency and averaged for further analysis. The corresponding electrically elicited torque was subsequently calculated using the previously described method.
Statistics
The results are reported as the mean ± SD. Before conducting statistical analysis, the normal distribution of the data was verified using the Shapiro–Wilk test. Since our results included non-normal distributed data and were based on small samples, we used non-parametric statistical tests. The Friedman test was used to compare all variables among PRE, POST1, and POST2. For the motor unit firing rate and RT, the Kruskal–Wallis test was used, and for other variables, the Friedman test was used to compare caffeine, quercetin, and placebo. When tests revealed significances, we performed Bonferroni post hoc tests for pair-wise comparisons. Associations between RT and change in RT (ΔRT) from PRE to POST1 and the change in firing rate (Δfiring rate) from POST1 to POST2 were analyzed by Spearman correlation coefficients (rs). The level of significance of the three-condition comparison was set at 0.05. Statistical analysis was performed using SPSS (version 21.0, SPSS, Tokyo, Japan).
Results
Number of identified motor units
During ramp contractions, 59, 49, and 52 motor units were tracked with caffeine, quercetin, and placebo during the given tasks, respectively, and they were used for analysis (Table 1). Figure 3a depicts representative data of motor unit firing patterns in one participant for three different conditions. In this participant, 5, 6 or 8 motor units were tracked among PRE, POST1, and POST2 in placebo, caffeine or quercetin conditions, respectively.

Representative data of motor unit firing patterns in one participant for three different conditions at before (PRE) and after (POST1) ingestions and after resistance exercise (POST2). Each motor unit is indicated by a different color. a Performed torque and motor unit firing behavior. b Motor unit recruitment threshold. c Motor unit firing rate. PLA, placebo condition; CAF, caffeine condition; QUE, quercetin condition; MVC, maximal voluntary contraction; POST1:PRE, POST2:PRE, and POST2:POST1, rate of change in motor unit activities from PRE to POST1, PRE to POST2 and POST1 to POST2
Recruitment threshold
Figure 3b displays the representative data of motor unit RTs and their rate of change for one participant in each condition. In this example, motor unit RTs were decreased in all conditions from PRE to POST2. Furthermore, decrease in motor unit RTs was observed from PRE to POST1 in the caffeine and quercetin conditions.
Under all conditions, RTs of motor units were significantly decreased from PRE to POST1 (placebo: p = 0.007, caffeine: p < 0.001, quercetin: p = 0.004) and POST2 (placebo: p < 0.001, caffeine: p < 0.001, quercetin: p < 0.001). From POST1 to POST2, RTs of motor units were significantly decreased with placebo and caffeine (placebo: p < 0.001, caffeine: p = 0.005) but not with quercetin ingestion (p = 0.060) (Fig. 4a). POST1:PRE was significantly lower with caffeine (p < 0.001) and quercetin (p = 0.045) than placebo. POST2:PRE was significantly lower with caffeine than placebo (p = 0.029) (Fig. 4b).

Effects of caffeine, quercetin, and placebo ingestions on motor unit recruitment thresholds. a Motor unit recruitment thresholds during ramp contractions before (PRE) and after (POST1) ingestions and after resistance exercise (POST2). Friedman test was used for analysis. * p < 0.05. b Rate of change in recruitment thresholds from PRE to POST1 (POST1:PRE), from PRE to POST2 (POST2:PRE), and from POST1 to POST2 (POST2:POST1). Kruskal–Wallis test was used for analysis. The lines within boxes, boxes, and bars indicate the median values, 25 and 75% of quartiles, and range of data, respectively. * p < 0.05
Firing rate
Figure 3c shows the representative data of motor unit FRs and their rate of change for one participant in each condition. From PRE to POST1 and PRE to POST2, motor unit FRs were increased in all conditions. The increase in motor unit FR from PRE to POST1 in caffeine or quercetin condition appears to exhibit slightly greater than in placebo condition.
Under all conditions, firing rates of motor units were significantly increased from PRE to POST1 (placebo: p = 0.019, caffeine: p < 0.001, quercetin: p < 0.001) and POST2 (placebo: p < 0.001, caffeine: p < 0.001, quercetin: p = 0.006), and from POST1 to POST2 (placebo: p < 0.001, caffeine: p < 0.001, quercetin: p < 0.001) (Fig. 5a). POST1:PRE was significantly higher with caffeine and quercetin than placebo (caffeine: p < 0.001, quercetin: p < 0.001). POST2:POST1 was significantly lower with quercetin than placebo (p = 0.018) (Fig. 5b).

Effects of caffeine, quercetin, and placebo ingestions on motor unit firing rates. a Motor unit firing rates during ramp contractions before (PRE) and after (POST1) ingestions and after resistance exercise (POST2). Friedman test was used for analysis. * p < 0.05. b Rate of change in firing rates from PRE to POST1 (POST1:PRE), from PRE to POST2 (POST2:PRE), and from POST1 to POST2 (POST2:POST1). Kruskal–Wallis test was used for analysis. The lines within boxes, boxes, and bars indicate the median values, 25 and 75% of quartiles, and range of data, respectively. * p < 0.05
Correlation between recruitment threshold and Δrecruitment threshold, recruitment threshold, and Δfiring rate
Caffeine and quercetin showed significant correlations between RT and ΔRT from PRE to POST1 (caffeine: rs = 0.448, p < 0.001, slope = − 0.12, quercetin: rs = 0.415, p = 0.003, slope = − 0.25), but the placebo did not (rs = 0.109, p = 0.440) (Fig. 6). With quercetin and placebo, significant correlations were observed between RT and Δfiring rate from POST1 to POST2 (Fig. 7) (placebo: rs = 0.382, p = 0.007, slope = 0.16, caffeine: rs = 0.069, p = 0.606, quercetin: rs = 0.380, p = 0.005, slope = 0.12).

Effects of caffeine, quercetin, and placebo ingestions on correlations (Spearman) between the motor unit recruitment threshold before ingestion (PRE) and recruitment threshold difference from PRE to POST1. Each circle depicts one tracked motor unit. Dotted black, solid red, and solid blue lines show linear regression lines for placebo, caffeine, and quercetin, respectively

Effects of caffeine, quercetin, and placebo ingestions on correlations (Spearman) between the motor unit recruitment threshold before ingestion (PRE) and firing rate difference from POST1 to POST2. Each circle depicts one tracked motor unit. Solid black, dotted red, and solid blue lines show linear regression lines for placebo, caffeine, and quercetin, respectively
Electrically elicited torque and MVC
Under all conditions, no significant changes were observed from PRE to POST1 in electrically elicited torque by twitch (all conditions: p > 0.05), 10 Hz (all conditions: p > 0.05), 20 Hz (all conditions: p > 0.05), 40 Hz (all conditions: p > 0.05), 80 Hz (all conditions: p > 0.05), and MVC (all conditions: p > 0.05) (Table 2, 3). A significant decrease in electrically elicited torque by twitch (all conditions: p < 0.05), 10 Hz (caffeine: p = 0.042, quercetin: p = 0.011, placebo: p = 0.042), 20 Hz (caffeine: p = 0.042, quercetin: p = 0.011, placebo: p = 0.022), 40 Hz (caffeine: p = 0.022, quercetin: p = 0.042, placebo: p = 0.011), 80 Hz (caffeine: p = 0.042, quercetin: p = 0.022, placebo: p = 0.022), and MVC (caffeine: p = 0.030, quercetin: p = 0.011, placebo: p = 0.022)from POST1 to POST2 was noted under all conditions (p < 0.05) (Table 2). None of POST1:PRE (all: p > 0.05) and POST2:POST1 (all: p > 0.05) for electrically elicited torque or MVC showed significant differences among conditions (Table 2, 3).
Discussion
The present study investigated the acute effect of caffeine or quercetin ingestion on the motor unit firing pattern and muscle contractile properties before and after resistance exercise. In this study, caffeine or quercetin ingestion: (1) lowered RT after ingestion, (2) had different threshold-dependent effects on motor unit activity after resistance exercise, and (3) had no effect on electrically elicited muscle contraction torques and MVC. These findings support our hypothesis that caffeine or quercetin ingestion would induce a threshold-dependent decrease in motor unit recruitment after ingestion and a threshold-dependent increase in the firing rate after resistance exercise. The hypothesis that caffeine or quercetin ingestion induces changes in muscle contractile properties after resistance exercise was rejected.
To our knowledge, this is the first study to investigate the acute effect of caffeine or quercetin ingestion on the motor unit firing pattern before and after resistance exercise. The merits of this study include the following: we identified individual motor units and tracked the same motor units in PRE, POST1, and POST2. The method in this study provides evidence of changes in RTs and firing rates of specific motor units (Figs.2, 3), allowing us to determine the threshold-dependent effects of supplementation and resistance exercise on each motor unit (Figs.4, 5).
Effects of ingestion.
In the present study, RTs were significantly reduced after capsule ingestion under all three conditions: caffeine, quercetin, and placebo (Fig. 4a), but the rate of change was significantly greater with caffeine and quercetin than placebo from PRE to POST1 (Fig. 4b). The rate of change of variables were calculated as measures of change in motor unit activities after capsule ingestion and exercise. Both caffeine and quercetin conditions showed a negative correlation between ΔRT (PRE–POST1) and RT at PRE (caffeine: rs = 0.448, p < 0.001; quercetin: rs = 0.415, p = 0.003), but no correlation was observed under the placebo condition (rs = 0.109, p = 0.440) (Fig. 6). These results suggest that caffeine and quercetin ingestions decrease RT of motor units with relatively higher RT. This finding for quercetin ingestion supports the results of our previous studies (Watanabe and Holobar 2021). We showed that quercetin ingestion decreased RT of motor units with higher RT. There are various factors that can influence the recruitment threshold of motor units, including modulation of synaptic inputs (ter Haar Romeny et al. 1982). Caffeine and quercetin have been demonstrated to function as antagonists of A1 adenosine receptors (Alexander 2006), and are considered to alter arousal and induce excitation in the central nervous system (Graham 2001; McLellan et al. 2016; Grgic et al. 2019). While the data from this study do not allow for a detailed description of the physiological pathways, it is hypothesized that caffeine and quercetin may alter synaptic input, leading to changes in neural drive and motor unit recruitment thresholds (ter Haar Romeny et al. 1982). The results of this study, as in previous studies (Watanabe and Holobar 2021; Bazzucchi et al. 2011; Patrizio et al. 2018), suggest that ingestions of 200 mg of caffeine and 500 mg of quercetin lower RTs of motor units with relatively higher RTs. Previous research showed that lower recruitment thresholds for motor units can occur after four weeks of training (Del Vecchio et al. 2019a), suggesting that these lower recruitment thresholds may be a neuromuscular strategy that promotes muscle performance, such as increasing the number of recruited motor units. Multiple studies reported that caffeine and quercetin consumptions can improve endurance exercise performance, and it has been proposed that the adenosine antagonist properties of these compounds may comprise a contributing mechanism (Kalmar and Cafarelli 1999; Graham 2001; Grgic 2021; Pickering and Grgic 2019). The lower recruitment thresholds of motor units in this study may reflect the physiological changes that lead to improved endurance performance with caffeine and quercetin ingestions, as reported in many previous studies.
Electrically elicited torque and MVC were not significantly different among the conditions of caffeine, quercetin, and placebo ingestions from PRE to POST1, suggesting that caffeine or quercetin ingestion have no effect on muscle contractile properties or maximal muscle strength. Several studies reported that caffeine and quercetin ingestions affect muscle contractile properties (Lopes et al. 1983; Bazzucchi et al. 2011; Watanabe and Holobar 2021), being inconsistent with the results of the present study. One possible reason for this may be that the doses used in this study were smaller compared with previous studies. Neyroud et al. (2019) suggested that for a direct action on skeletal muscle and force to potentiate basal/resting conditions, toxic doses of caffeine for humans would be required (Neyroud et al. 2019). Therefore, it is reasonable to consider that the effect of the dose utilized in this study on peripheral factors was not discernible, whereas its effect on the central nervous system was observable. Our results suggested that 200 mg of caffeine and 500 mg of quercetin have no effect on muscle contractile properties or maximal muscle strength.
Effects of ingestion after resistance exercise.
In the present study, the firing rate was significantly increased after resistance exercise under all three conditions: caffeine, quercetin, and placebo (Fig. 5), but the rate of change was significantly lower under the quercetin than placebo condition. A significant correlation was noted between Δfiring rate (PRE–POST2) and RT at PRE, noted with placebo (rs = 0.380, p = 0.005) and quercetin (rs = 0.382, p = 0.007), but not caffeine (rs = 0.069, p = 0.606) (Fig. 7). Although same motor units were not tracked among the conditions for caffeine, quercetin and placebo, results of correlation analyses showed increases and decreases in Δfiring rate (PRE–POST2) in motor units with relatively lower RT following the ingestion of caffeine or quercetin, respectively. Lynge and Hellsten (2001) showed that the number of all three of A1, A2A, and A2B adenosine receptors in vascular cells of skeletal muscle tissue and cytosolic staining of the adenosine A2A receptor was greater in type I muscle fibers, whereas the A2B receptor was almost absent in type I fibers (Lynge and Hellsten 2000). This finding is relevant to our data, showing that caffeine ingestion increases firing rates of motor units with relatively lower RT. On the other hand, our previous study showed that quercetin ingestion decreases firing rates of motor units with relatively lower RT (Watanabe and Holobar 2021). To elaborate, caffeine and quercetin exhibit distinct effects on motor units that are mobilized at low recruitment thresholds, suggesting that both quercetin and caffeine stimulate motor units with distinct recruitment thresholds. Also, Patrizio et al. (2018) and Bazzucchi et al. (2019) showed an increase in the median frequency of surface EMG (Patrizio et al. 2018) and a relative increase in muscle fiber conduction velocity following quercetin ingestion (Bazzucchi et al. 2019). Thus, these findings suggest that quercetin ingestion enhances the recruitment of motor units with relatively higher RT (Patrizio et al. 2018; Bazzucchi et al. 2019), which could compensate for the decrease in firing rates of motor units with relatively lower RT (Watanabe and Holobar 2021). These differences in firing rates of motor units with relatively lower RT would explain differences in recruitment threshold-dependent alternations in the motor unit firing pattern between caffeine and quercetin. Previous research showed that interventions that depend on the recruitment threshold can have different effects. For example, Ross et al. (2010) demonstrated that whole-body vibration increased the recruitment thresholds of motor units recruited at low thresholds and decreased those of motor units recruited at high thresholds (Pollock et al. 2012). Motor units recruited at low recruitment thresholds are activated via mono-synaptic pathways, while those recruited at high thresholds are activated through both mono- and polysynaptic pathways (Romaiguere et al. 1991). These findings suggest that caffeine and quercetin may potentially act on distinct synaptic pathways.
Electrically elicited torque and MVC decreased from POST1 to POST2 under all three conditions, but there was no significant difference in electrically elicited torque, MVC, or each rate of change among caffeine, quercetin, and placebo. Our results revealed that caffeine or quercetin ingestion has no effect on muscle contractile properties (peripheral fatigue) or muscle strength with resistance exercise, suggesting that caffeine or quercetin ingestion at these doses affects the neuromuscular function, but not peripheral fatigue. Therefore, the limiting factor for decreasing MVC in this study may have been central rather than peripheral.
Limitations
The participants of this study were young males. Thus, the present findings may not be generalizable to female or older individuals. Females were excluded from this study for several reasons. The first involves the sensitivity of motor units detected between the sexes, with prior studies demonstrating lower detectability of motor units in females compared with males (Lulic-Kuryllo and Inglis 2022). The second reason is that a more complex cohort would be necessary if including females, taking into account factors such as the effects of the menstrual cycle (Pickering and Grgic 2019). Most previous studies examining the effects of caffeine on exercise only recruited males (Grgic et al. 2019). However, the findings of this study may be generalizable to the female population. For example, Skinner et al. (2019) reported that the magnitude of the ergogenic effect was approximately the same between males and females after consuming 3 mg/kg of caffeine 90 min prior to a cycling test (Skinner et al. 2019), suggesting that caffeine and quercetin may be ergogenic for females as well as males. In addition to the young males included in this study, the effects of caffeine and quercetin on females and the elderly require further investigation.
Additionally, this study utilized small, absolute doses of caffeine (200 mg, 2.8–3.7 mg/kg) and quercetin (500 mg, 6.9–9.3 mg/kg). In many previous studies, dosages standardized by body weight were administered; in this study, the weight-normalized dosages varied among individuals. Previous research did not report differences in efficacy across the range of inter-individual differences in weight-normalized dosages used in this study (Desbrow et al. 2012). Furthermore, this study employed smaller dosages than in previous studies; Lawrence et al. reported that low doses of caffeine (< 3 mg/kg, approximately 200 mg) prior to exercise can alter central nervous system activity and produce an ergogenic effect (Spriet 2014). Our previous study also reported that the ingestion of quercetin at 500 mg, the same dosage as used in the present study, affected motor unit activity (Watanabe and Holobar 2021). In previous studies, 1000 mg of quercetin aglycones were administered orally per day (Patrizio et al. 2018; Bazzucchi et al. 2019), but in this study, quercetin glycosides were used; quercetin glycosides have a tenfold higher bioavailability compared with quercetin aglycones (Murota et al. 2010). Therefore, it cannot be stated that the dosage used in this study was small in comparison with previous studies. This study examined the effects of single, small, absolute doses of caffeine and quercetin on the neuromuscular function. Further research is required to determine the effects of various dosages of caffeine and quercetin on motor unit activity and muscle contractile properties.
Conclusion
The present study investigated the effect of caffeine or quercetin ingestion on motor unit firing and muscle contractile properties before and after resistance exercise. Caffeine or quercetin ingestion decreased RT in a threshold-dependent manner after ingestion, and caffeine ingestion increased the firing rate of motor units with relatively lower RT, while quercetin ingestion decreased the firing rate of motor units with relatively lower RT after resistance exercise. Our data also suggest that, for the limited and investigated dosages, caffeine and quercetin ingestions do not affect muscle contractile properties. The present study concluded that caffeine or quercetin ingestion alters motor unit firing patterns after resistance exercise in different RT-dependent manners. Further studies are necessary to clarify the effects of caffeine and quercetin on the neuromuscular properties of populations not represented in this study (e.g., females and older adults).
Data availability
Data that support the findings of the present study are presented in the text, figures and table, and are available from the corresponding author upon reasonable request.
Abbreviations
- HDs-EMG:
-
High-density surface electromyography
- MVC:
-
Maximum voluntary contraction
- RT:
-
Recruitment threshold
- VL:
-
Vastus lateral
References
Adam A (2005) Firing rates of motor units in human vastus lateralis muscle during fatiguing isometric contractions. J Appl Physiol 99(1):268–280. https://doi.org/10.1152/japplphysiol.01344.2004
Alexander SP (2006) Flavonoids as antagonists at A1 adenosine receptors. Phytother Res 20(11):1009–1012. https://doi.org/10.1002/ptr.1975
Bazzucchi I, Felici F, Montini M, Figura F, Sacchetti M (2011) Caffeine improves neuromuscular function during maximal dynamic exercise. Muscle Nerve 43(6):839–844. https://doi.org/10.1002/mus.21995
Bazzucchi I, Patrizio F, Ceci R, Duranti G, Sgro P, Sabatini S, Di Luigi L, Sacchetti M, Felici F (2019) The effects of quercetin supplementation on eccentric exercise-induced muscle damage. Nutrients. https://doi.org/10.3390/nu11010205
Burak C, Brull V, Langguth P, Zimmermann BF, Stoffel-Wagner B, Sausen U, Stehle P, Wolffram S, Egert S (2017) Higher plasma quercetin levels following oral administration of an onion skin extract compared with pure quercetin dihydrate in humans. Eur J Nutr 56(1):343–353. https://doi.org/10.1007/s00394-015-1084-x
Burke RE, Levine DN, Tsairis P, Zajac FE 3rd (1973) Physiological types and histochemical profiles in motor units of the cat gastrocnemius. J Physiol 234(3):723–748. https://doi.org/10.1113/jphysiol.1973.sp010369
Cheuvront SN, Ely BR, Kenefick RW, Michniak-Kohn BB, Rood JC, Sawka MN (2009) No effect of nutritional adenosine receptor antagonists on exercise performance in the heat. Am J Physiol Regul Integr Comp Physiol 296(2):R394-401. https://doi.org/10.1152/ajpregu.90812.2008
Coombes JS, Powers SK, Rowell B, Hamilton KL, Dodd SL, Shanely RA, Sen CK (2001) Effects of vitamin E and alpha-lipoic acid on skeletal muscle contractile properties. J Appl Physiol 90(4):1424–1430. https://doi.org/10.1152/jappl.2001.90.4.1424
Coombes JS, Rowell B, Dodd SL, Demirel HA, Naito H, Shanely RA, Powers SK (2002) Effects of vitamin E deficiency on fatigue and muscle contractile properties. Eur J Appl Physiol 87(3):272–277. https://doi.org/10.1007/s00421-002-0631-3
Davis JM, Murphy EA, Carmichael MD (2009) Effects of the dietary flavonoid quercetin upon performance and health. Curr Sports Med Rep 8(4):206–213. https://doi.org/10.1249/JSR.0b013e3181ae8959
Del Vecchio A, Casolo A, Negro F, Scorcelletti M, Bazzucchi I, Enoka R, Felici F, Farina D (2019a) The increase in muscle force after 4 weeks of strength training is mediated by adaptations in motor unit recruitment and rate coding. J Physiol 597(7):1873–1887. https://doi.org/10.1113/JP277250
Del Vecchio A, Negro F, Holobar A, Casolo A, Folland JP, Felici F, Farina D (2019b) You are as fast as your motor neurons: speed of recruitment and maximal discharge of motor neurons determine the maximal rate of force development in humans. J Physiol 597(9):2445–2456. https://doi.org/10.1113/JP277396
Desbrow B, Biddulph C, Devlin B, Grant GD, Anoopkumar-Dukie S, Leveritt MD (2012) The effects of different doses of caffeine on endurance cycling time trial performance. J Sports Sci 30(2):115–120. https://doi.org/10.1080/02640414.2011.632431
Farina D, Holobar A, Merletti R, Enoka RM (2010) Decoding the neural drive to muscles from the surface electromyogram. Clin Neurophysiol 121(10):1616–1623. https://doi.org/10.1016/j.clinph.2009.10.040
Francic A, Holobar A (2021) On the reuse of motor unit filters in high density surface electromyograms recorded at different contraction levels. Ieee Access 9:115227–115236. https://doi.org/10.1109/Access.2021.3104762
Fuglevand AJ, Winter DA, Patla AE (1993) Models of recruitment and rate coding organization in motor-unit pools. J Neurophysiol 70(6):2470–2488. https://doi.org/10.1152/jn.1993.70.6.2470
Gallego JA, Dideriksen JL, Holobar A, Ibanez J, Glaser V, Romero JP, Benito-Leon J, Pons JL, Rocon E, Farina D (2015a) The phase difference between neural drives to antagonist muscles in essential tremor is associated with the relative strength of supraspinal and afferent input. J Neurosci 35(23):8925–8937. https://doi.org/10.1523/JNEUROSCI.0106-15.2015
Gallego JA, Dideriksen JL, Holobar A, Ibanez J, Pons JL, Louis ED, Rocon E, Farina D (2015b) Influence of common synaptic input to motor neurons on the neural drive to muscle in essential tremor. J Neurophysiol 113(1):182–191. https://doi.org/10.1152/jn.00531.2014
Graham TE (2001) Caffeine and exercise: metabolism, endurance and performance. Sports Med 31(11):785–807. https://doi.org/10.2165/00007256-200131110-00002
Grgic J (2021) Effects of caffeine on resistance exercise: a review of recent research. Sports Med 51(11):2281–2298. https://doi.org/10.1007/s40279-021-01521-x
Grgic J, Mikulic P, Schoenfeld BJ, Bishop DJ, Pedisic Z (2019) The influence of caffeine supplementation on resistance exercise: a review. Sports Med 49(1):17–30. https://doi.org/10.1007/s40279-018-0997-y
Grgic J, Grgic I, Pickering C, Schoenfeld BJ, Bishop DJ, Pedisic Z (2020) Wake up and smell the coffee: caffeine supplementation and exercise performance-an umbrella review of 21 published meta-analyses. Br J Sports Med 54(11):681–688. https://doi.org/10.1136/bjsports-2018-100278
Haider G, Folland JP (2014) Nitrate supplementation enhances the contractile properties of human skeletal muscle. Med Sci Sports Exerc 46(12):2234–2243. https://doi.org/10.1249/MSS.0000000000000351
Henneman E, Olson CB (1965) Relations between Structure and Function in the Design of Skeletal Muscles. J Neurophysiol 28:581–598. https://doi.org/10.1152/jn.1965.28.3.581
Henneman E, Somjen G, Carpenter DO (1965) Functional Significance of Cell Size in Spinal Motoneurons. J Neurophysiol 28:560–580. https://doi.org/10.1152/jn.1965.28.3.560
Holobar A, Zazula D (2004) Correlation-based decomposition of surface electromyograms at low contraction forces. Med Biol Eng Comput 42(4):487–495. https://doi.org/10.1007/BF02350989
Holobar A, Zazula D (2008) On the selection of the cost function for gradient-based decomposition of surface electromyograms. Annu Int Conf IEEE Eng Med Biol Soc 2008:4668–4671. https://doi.org/10.1109/IEMBS.2008.4650254
Holobar A, Farina D, Gazzoni M, Merletti R, Zazula D (2009) Estimating motor unit discharge patterns from high-density surface electromyogram. Clin Neurophysiol 120(3):551–562. https://doi.org/10.1016/j.clinph.2008.10.160
Holobar A, Minetto MA, Farina D (2014) Accurate identification of motor unit discharge patterns from high-density surface EMG and validation with a novel signal-based performance metric. J Neural Eng 11(1):016008. https://doi.org/10.1088/1741-2560/11/1/016008
Kalmar JM (1985) Cafarelli E (1999) Effects of caffeine on neuromuscular function. J Appl Physiol 87(2):801–808. https://doi.org/10.1152/jappl.1999.87.2.801
Kamimori GH, Karyekar CS, Otterstetter R, Cox DS, Balkin TJ, Belenky GL, Eddington ND (2002) The rate of absorption and relative bioavailability of caffeine administered in chewing gum versus capsules to normal healthy volunteers. Int J Pharm 234(1–2):159–167. https://doi.org/10.1016/s0378-5173(01)00958-9
Kunugi S, Holobar A, Kodera T, Toyoda H, Watanabe K (2021) Motor unit firing patterns on increasing force during force and position tasks. J Neurophysiol 126(5):1653–1659. https://doi.org/10.1152/jn.00299.2021
Lee EH, Meissner G, Kim DH (2002) Effects of quercetin on single Ca(2+) release channel behavior of skeletal muscle. Biophys J 82(3):1266–1277. https://doi.org/10.1016/S0006-3495(02)75483-0
Lopes JM, Aubier M, Jardim J, Aranda JV, Macklem PT (1983) Effect of caffeine on skeletal muscle function before and after fatigue. J Appl Physiol Respir Environ Exerc Physiol 54(5):1303–1305. https://doi.org/10.1152/jappl.1983.54.5.1303
Lulic-Kuryllo T, Inglis JG (2022) Sex differences in motor unit behaviour: A review. J Electromyogr Kinesiol 66:102689. https://doi.org/10.1016/j.jelekin.2022.102689
Lynge J, Hellsten Y (2000) Distribution of adenosine A1, A2A and A2B receptors in human skeletal muscle. Acta Physiol Scand 169(4):283–290. https://doi.org/10.1046/j.1365-201x.2000.00742.x
Makino T, Shimizu R, Kanemaru M, Suzuki Y, Moriwaki M, Mizukami H (2009) Enzymatically modified isoquercitrin, alpha-oligoglucosyl quercetin 3-O-glucoside, is absorbed more easily than other quercetin glycosides or aglycone after oral administration in rats. Biol Pharm Bull 32(12):2034–2040. https://doi.org/10.1248/bpb.32.2034
McLellan TM, Caldwell JA, Lieberman HR (2016) A review of caffeine’s effects on cognitive, physical and occupational performance. Neurosci Biobehav Rev 71:294–312. https://doi.org/10.1016/j.neubiorev.2016.09.001
Merletti R, Holobar A, Farina D (2008) Analysis of motor units with high-density surface electromyography. J Electromyogr Kinesiol 18(6):879–890. https://doi.org/10.1016/j.jelekin.2008.09.002
Murota K, Matsuda N, Kashino Y, Fujikura Y, Nakamura T, Kato Y, Shimizu R, Okuyama S, Tanaka H, Koda T, Sekido K, Terao J (2010) alpha-Oligoglucosylation of a sugar moiety enhances the bioavailability of quercetin glucosides in humans. Arch Biochem Biophys 501(1):91–97. https://doi.org/10.1016/j.abb.2010.06.036
Neyroud D, Cheng AJ, Donnelly C, Bourdillon N, Gassner AL, Geiser L, Rudaz S, Kayser B, Westerblad H, Place N (2019) Toxic doses of caffeine are needed to increase skeletal muscle contractility. Am J Physiol Cell Physiol 316(2):C246–C251. https://doi.org/10.1152/ajpcell.00269.2018
Patrizio F, Ditroilo M, Felici F, Duranti G, De Vito G, Sabatini S, Sacchetti M, Bazzucchi I (2018) The acute effect of Quercetin on muscle performance following a single resistance training session. Eur J Appl Physiol 118(5):1021–1031. https://doi.org/10.1007/s00421-018-3834-y
Pickering C, Grgic J (2019) Caffeine and Exercise: What Next? Sports Med 49(7):1007–1030. https://doi.org/10.1007/s40279-019-01101-0
Pollock RD, Woledge RC, Martin FC (2012) Effects of whole body vibration on motor unit recruitment and threshold. J Appl Physiol 112(3):388–395. https://doi.org/10.1152/japplphysiol.01223.2010
Romaiguere P, Vedel JP, Azulay JP, Pagni S (1991) Differential activation of motor units in the wrist extensor muscles during the tonic vibration reflex in man. J Physiol 444:645–667. https://doi.org/10.1113/jphysiol.1991.sp018899
Skinner TL, Desbrow B, Arapova J, Schaumberg MA, Osborne J, Grant GD, Anoopkumar-Dukie S, Leveritt MD (2019) Women experience the same ergogenic response to caffeine as men. Med Sci Sports Exerc 51(6):1195–1202. https://doi.org/10.1249/MSS.0000000000001885
Spriet LL (2014) Exercise and sport performance with low doses of caffeine. Sports Med 44(2):S175-184. https://doi.org/10.1007/s40279-014-0257-8
ter Haar Romeny BM, Denier van der Gon JJ, Gielen CC (1982) Changes in recruitment order of motor units in the human biceps muscle. Exp Neurol 78(2):360–368. https://doi.org/10.1016/0014-4886(82)90054-1
Tomita A, Kawade S, Moritani T, Watanabe K (2020) Novel perspective on contractile properties and intensity-dependent verification of force-frequency relationship during neuromuscular electrical stimulation. Physiol Rep 8(22):e14598. https://doi.org/10.14814/phy2.14598
Watanabe K, Holobar A (2021) Quercetin ingestion modifies human motor unit firing patterns and muscle contractile properties. Exp Brain Res 239(5):1567–1579. https://doi.org/10.1007/s00221-021-06085-w
Watanabe K, Holobar A, Kouzaki M, Ogawa M, Akima H, Moritani T (2016) Age-related changes in motor unit firing pattern of vastus lateralis muscle during low-moderate contraction. Age (dordr) 38(3):48. https://doi.org/10.1007/s11357-016-9915-0
Watanabe K, Holobar A, Mita Y, Kouzaki M, Ogawa M, Akima H, Moritani T (2018) Effect of resistance training and fish protein intake on motor unit firing pattern and motor function of elderly. Front Physiol 9:1733. https://doi.org/10.3389/fphys.2018.01733
Watanabe K, Holobar A, Mita Y, Tomita A, Yoshiko A, Kouzaki M, Uchida K, Moritani T (2020) Modulation of neural and muscular adaptation processes during resistance training by fish protein ingestions in older adults. J Gerontol A Biol Sci Med Sci 75(5):867–874. https://doi.org/10.1093/gerona/glz215
Yavuz US, Negro F, Sebik O, Holobar A, Frommel C, Turker KS, Farina D (2015) Estimating reflex responses in large populations of motor units by decomposition of the high-density surface electromyogram. J Physiol 593(19):4305–4318. https://doi.org/10.1113/JP270635
Acknowledgements
This study was supported by the Bilateral Program of JSPS (JPJSBP-82626) to KW and AH and by the Slovenian Research Agency (project J2-1731, L7-9421 and Program funding P2-0041) to AH.
Author information
Authors and Affiliations
Contributions
TN contributed to analysis and interpretation of data; and drafting the article and revising it critically for important intellectual content. TN, TH, AH, SK, MO and TO contributed to conception and design of the experiments; collection, analysis, and interpretation of data; and revising the article for important intellectual content. K.W. obtained funding support; contributed to collection, analysis and interpretation of data; and also contributed to drafting the article and revising it critically for important intellectual content. All authors approved the final version of the manuscript. The authors have no disclosures.
Corresponding author
Ethics declarations
Conflict of interest
There are no competing interests.
Additional information
Communicated by Westerterp/Westerblad.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Nishikawa, T., Hirono, T., Holobar, A. et al. Acute effects of caffeine or quercetin ingestion on motor unit firing pattern before and after resistance exercise. Eur J Appl Physiol 124, 1645–1658 (2024). https://doi.org/10.1007/s00421-023-05376-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-023-05376-0