
Abstract
Purpose
This study aimed to determine whether an increase in muscle shear modulus measured 30 min after eccentric exercise (30 min) reflects the magnitude of force deficit measured 48-h post-exercise (48 H).
Methods
A total of 53 healthy participants were distributed in five groups. Four groups performed either repeated eccentric elbow flexions or knee extensions at either a low or high load. A fifth group performed repeated concentric elbow flexions (control load).
Results
A significant decreased peak torque was found for elbow flexors and knee extensors 48 h after the eccentric exercises (all P values < 0.001). A significant increase in shear modulus was found at 30 min for the elbow flexors for low (+70.5 ± 44.3%, P < 0.001) and high load (+153.9 ± 192.4%, P < 0.001). Similarly, the shear modulus of knee extensors increased for low (+26.7 ± 19.1%, P < 0.001) and high load (+79.4 ± 67.1%, P < 0.001). The relative increase in shear modulus measured at 30 min was significantly correlated to the relative decrease in peak torque measured at 48 H for both elbow flexors (r = −0.80) and knee extensors (r = −0.82). A further analysis suggested that biceps brachii and rectus femoris were more affected by muscle damage than their synergists.
Conclusion
This study shows that an increase in muscle shear modulus measured 30 min after a damaging exercise reflects the decrease in peak torque measured at 48 H. Shear modulus may therefore, provide a useful tool for coaches and clinicians to non-invasively estimate the amount of muscle damage induced by a damaging exercise.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Muscle damage after eccentric muscle action is a common experience in sports or daily life activities. Accurate quantification of muscle damage requires histological examination of muscle biopsy samples by determining the number of myofibrillar disruptions (Raastad et al. 2010). As this is an invasive procedure, it is unlikely to be routinely used in sport settings and clinical practice. Although the amount of muscle damage can be accurately evaluated through the time course of the strength loss measured during concentric actions across a large range of motion including the angle at peak torque (Paulsen et al., 2012), the maximal isometric force deficit observed 48-h post-exercise (48 H) is also considered as a good non-invasive marker of muscle damage (Paulsen et al. 2012). However, these measurements do not provide an estimation of muscle damage immediately after exercise due to the confounding influence of muscle fatigue (Warren et al. 1999). In addition, force deficit is assessed through global joint torque that does not provide any information about the extent of damage in each individual muscle. A non-invasive method able to early detect muscle damage in individual muscles that would benefit both musculoskeletal rehabilitation and sport practice.
Eccentric contractions induce an immediate increase in muscle stiffness evidenced by an alteration of the passive force–length relationship (Hoang et al. 2007). Using shear wave elastography to quantify local changes in muscle shear modulus [i.e., an index of stiffness, (Eby et al. 2013)], our team showed that the muscle shear modulus of elbow flexor and plantar flexor muscles significantly increases in the early instants after exercise (Lacourpaille et al. 2014; Guilhem et al. 2016). We hypothesized that this finding could originate from the perturbation of calcium homeostasis (Whitehead et al. 2001). In that case, the increase in passive muscle shear modulus subsequent to a damaging exercise would be closely related to the number of myofibrillar disruptions and membrane damage resulting in calcium overload.
This study was designed to determine whether the early increase in muscle shear modulus (evaluated by shear wave elastography) after eccentric contractions reflect the magnitude of force deficit measured at 48 H, and therefore, the amount of muscle damage. We hypothesized that there would be a strong linear relationship between the relative increase in shear modulus measured 30 min after exercise (30 min) and the relative force deficit at 48 H. To induce a wide range of muscle damage, the exercise was performed on muscles more or less prone to damage (elbow flexors and knee extensors, respectively) (Jamurtas et al. 2005), and under low or high loading conditions. A secondary aim was to determine whether synergist muscles are similarly affected by the eccentric exercise. In this view we compared the index of increase in shear modulus measured at 30 min between the synergist muscles.
Materials and methods
Subjects
A total of 53 healthy participants (35 males and 18 females; age: 24.6 ± 3.4 years; height: 172.6 ± 8.7 cm; weight: 66.7 ± 10.4 kg) volunteered for this study. Four groups performed eccentric contractions with either their elbow flexor or knee extensor muscles and at either a low or high load (described in details below). To verify that the change in muscle shear modulus was induced by muscle damage rather than the exercise itself [e.g., increase in intramuscular pressure (Davis et al. 2003)], an additional group of participants performed concentric contractions (control load). This was only performed on elbow flexors because previous works demonstrate the ability of shear wave elastography to detect slight changes on this muscle group (Bouillard et al. 2012b; Lacourpaille et al. 2014; Eby et al. 2015, 2016; Agten et al. 2016). Participant characteristics are described in Table 1.
All the participants were informed regarding the nature, aims and risks associated with the experimental procedures before their written consent was given. The local ethics committee approved the study (CPP IDF VIII, CPP-MIP-004P), and all the procedures conformed to the Declaration of Helsinki. Data from the group that performed the eccentric exercise with their elbow flexor muscles at a low load have been published in a separate communication that addressed a different research question (Lacourpaille et al. 2014).
Instrumentation
Ergometer
All experiments were performed on a Con-Trex MJ isokinetic dynamometer (CMV AG, Dubendorf, Switzerland). The axis of the dynamometer was aligned with the axis of rotation of the tested joint. All mechanical signals provided by the dynamometer were sampled at 512 Hz.
Elastography
An Aixplorer ultrasound scanner (version 7.0, Supersonic Imagine, Aix-en-Provence, France) coupled with a linear transducer array (4–15 MHz, SuperLinear 15-4; Vermon, Tours, France) was used in shear wave elastography mode (musculoskeletal preset) as previously described (Bercoff et al. 2004). This technique provides a two-dimensional map of shear modulus of a localized area in real-time at one sample/s, and it is linearly related to Young’s modulus measured using traditional material testing (Eby et al. 2013). Therefore, it provides a relevant characterization of passive muscle stiffness (Eby et al. 2013). Good reliability of this technique has been demonstrated (Lacourpaille et al. 2012). The inter-day coefficient of variation and standard error of measurement of shear modulus were 5.4% and 0.2 kPa, respectively, for the biceps brachii (Lacourpaille et al. 2012).
Experimental design
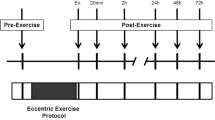
The experiments were conducted on two separate days. Participants first performed an initial test session (PRE) where peak torque during maximal isometric contraction and resting muscle shear modulus were measured. This test session was followed by an eccentric or concentric exercise depending on the respective group. Muscle shear modulus was then measured 30 min after the exercise. Peak voluntary isometric torque was assessed during the second session performed at 48 H.
Interventions
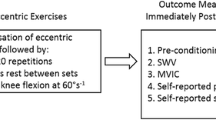
Eccentric knee extension
Participants performed 5 sets of 15 or 30 maximal isokinetic (60° s−1) eccentric contractions for the “low load” and “high load” groups, respectively. The exercise was executed with their dominant leg from 10° to 110° of knee angle (0° = full extension). The leg was passively repositioned at 10° after each eccentric contraction. Each set was followed by 1 min 30 s of a passive recovery period.
Eccentric elbow flexion
Participants completed 3 or 6 sets of 10 maximal isokinetic (120° s−1) eccentric contractions for the “low load” and “high load” groups, respectively. The eccentric contractions were performed with their dominant arm from a 60° to a 175° elbow angle (180° = full extension) with a shoulder abduction angle of 0°, i.e., arm along the body. Between each contraction, the arm was passively repositioned at 60°. A passive recovery period of 1 min 30 s was given between each set.
Concentric elbow flexion (control load)
A concentric exercise was performed to evaluate the effect of non-damaging contractions on post-exercise muscle shear modulus. Participants completed 3 sets of 10 maximal isokinetic (at 120° s−1) concentric contractions of the elbow flexor muscles of the dominant arm. The concentric contractions were performed from 175° to 60° of elbow joint angle with the same arm repositioning and recovery as during eccentric contractions of elbow flexors.
Test sessions
Peak torque
Considering that the force deficit at 48 H is a robust indicator of functional impairments due to muscle damage (Paulsen et al. 2012), peak voluntary torque was measured before and 48 h after the eccentric exercise during 5 s maximal voluntary isometric contractions (MVC). This evaluation was performed at a knee and elbow angle of 70° and 90°, respectively, after a warm-up of 20 isokinetic concentric extensions and flexions (120° s−1). A total of three contractions were performed, and the contraction with the highest isometric peak torque was considered for further analysis. Participants were vigorously encouraged to produce the greatest torque possible during each contraction.
Elastography measurements
Muscle shear modulus was measured at rest before and 30 min after the exercise. Indeed, this study aimed to provide an early, non-invasive, and sensitive marker of muscle damage. Therefore, we would found a tradeoff between an immediate measurement after exercise and a significant increase in muscle shear modulus. To this aim, we did a pilot experiment to determine the time course of the increase in biceps brachii shear modulus during the first hour after exercise (one measurement every 10 min; unpublished data). We found that the shear modulus increased at 10 min after exercise and remained stable over the first hour. In addition, it has been shown that the measurement of the shear modulus is not altered during a submaximal isometric fatiguing task performed until task failure (Bouillard et al. 2012a). By convenience, we took a safety margin and chose to measure the shear modulus at 30 min after exercise.
For the knee extensors measurements, participants were seated on the dynamometer chair with their hip flexed at 85° (0° = full extension). Resting shear modulus measurements were performed on vastus lateralis (VL), vastus medialis (VM), and rectus femoris (RF) at 30°, 90°, and 110° of knee angle on the exercised leg (0° = full extension).
For elbow flexors, participants were seated on the dynamometer chair with their right arm positioned in the horizontal plane and the shoulder abducted to 90° (Lacourpaille et al. 2013, 2014). Resting shear modulus of biceps brachii (BB) and brachialis (BA) were measured at 70°, 110°, and 160° of elbow angle (0° = full extension).
As described elsewhere (Lacourpaille et al. 2014), the optimal transducer location was determined using a B-mode ultrasound image as a region with a muscle thickness of at least 1 cm, that avoided hypoechoic regions that relate to fascia and tendon tissue. The ultrasound transducer was positioned within the direction of the muscle shortening for each muscle and perpendicularly to the skin. This location was marked on the skin using a waterproof marker so that the transducer location remained constant between recordings. For each recording, muscle shear modulus was measured for 10 s (10 values were recorded at 1 sample per second), and participants were instructed to remain as relaxed as possible (Lacourpaille et al. 2012, 2014).
Data processing
Analysis of elastography data was performed using Aixplorer scanner software. Shear modulus values were averaged over a 15-mm-circle region of interest (ROI) (Q-Box function). Shear modulus values were averaged over the ten consecutive measurements to obtain a representative value.
The increase in muscle shear modulus after exercise-induced muscle damage is larger at longer muscle length (Lacourpaille et al. 2014). It is therefore impossible to compare the change in shear modulus between muscles if they are at a different length relative to their optimal length. Despite not being problematic for comparison of monoarticular synergist muscles, it becomes important to consider for comparison between monoarticular and biarticular synergist muscles. To overcome this limitation, we calculated an index of increase in shear modulus independent of muscle length. Using previously collected data from passive loading/unloading stretching cycles (Fig. 1a) (Lacourpaille et al. 2014), we found that the shape of the relationship between elbow joint angle (above the slack length ~95°) and the increase in muscle shear modulus 30 min after the exercise (kPa) is linear (R 2 = 0.83 and R 2 = 0.99, for individual and overall data, respectively, see Fig. 1b). In the current study, we estimated the slope of the relationship between the change in shear modulus (PRE—30 min) and the joint angle. We considered the two joint angles at which the muscles were most stretched (i.e., 90° and 110° for the knee extensors and 110° and 160° for the elbow flexors) to calculate the index of increase in shear modulus (expressed in %). This index allowed us to compare the increase in shear modulus between the synergist muscles regardless of their respective length.

a Relationship between biceps brachii muscle shear modulus and elbow joint angle (from 70° to 160°) obtained before (PRE) and 30 min after a damaging exercise (data from Lacourpaille et al. 2014). b Linear relationship was obtained between elbow joint angle (above the slack length; from 95° to 160°) and the absolute increase in muscle shear modulus at 30 min (in kPa). The two red arrows represent the two angles used in the present study to calculate the index of increase in muscle shear modulus. Typical examples of the shear modulus map (i.e., index of stiffness) superposed onto the B-mode image, with the color scale depicting graduation of shear elastic modulus (in kPa, blue being soft and red being stiff). To obtain a representative value, the shear modulus was averaged over the region of interest (white circle) (color figure online)
Statistical analysis
All analyses were performed using Statistica Version 7.0 (StatSoft, Tulsa, OK). Normality testing (Kolmogorov–Smirnov) and sphericity assumption (Mauchly’s test) were consistently passed and values are reported as mean ± SD. A one-way [within-subject factor: time (PRE and 48 H)] and a three-way ANOVA [within-subject factors: time (PRE and 30 min), muscle (BB and BA, or VM, VL and RF) and angle (70°, 110°, and 160° of elbow extension, or 30°, 90°, and 110° of knee flexion)] were performed to test the effect of the control load on torque and shear modulus, respectively. The effect of time and load on peak torque was assessed for each muscle group separately (elbow flexors and knee extensors) using a two-way ANOVA [within-subject factor: time (PRE and 48 H) and between-subject factor: load (low and high)]. A four-way ANOVA (within-subject factor: time, muscle, angle, between-subject factor: load) was performed to test the effect of the eccentric exercise on shear modulus. We ran a separate two-way ANOVA [within-subject factor: muscle (BB and BA) or (VM, VL and RF), between-subject factor: load (low and high)] for each muscle group to determine whether the index of increase in shear modulus was different between synergistic muscles. When appropriate, post hoc analyses were performed using the Newman-Keuls test. The effect size was calculated using Cohen’s d considering 0.2, 0.5, and 0.8 as a small, medium and large effect, respectively (Cohen 1988). The significance level was set to P < 0.05.
For both the elbow flexors and knee extensors, Pearson’s correlation coefficient was calculated between the relative changes in shear modulus averaged over all the synergist muscles (in % of PRE) at 30 min and the relative changes in force loss at 48 H, by pooling both low and high loads. For this test we considered the joint angle at which the highest increase in shear modulus was observed (i.e., 160° of elbow extension and 110° of knee flexion).
Results
Peak torque
There was no significant effect of time (PRE vs. 48H) on maximal elbow flexion torque for control load [P = 0.45 (d = 0.10)] (Fig. 2). In contrast, there was a significant effect of time for maximal elbow flexion torque showing a global decrease in peak torque at 48 H (−26.2 ± 19.9%; P < 0.001) regardless of the load level (no significant time × load interaction; P = 0.117) (Fig. 2). A significant time × load interaction (P = 0.044) was found for the knee extension peak torque. More precisely, peak knee extensor torque was significantly lower at 48 H compared to PRE for low [P = 0.012 (d = 0.75)] and high load [P = 0.008 (d = 1.20)]. In addition, the peak torque decrease at 48 H was larger for high (−33.7 ± 19.6%) than low load (−16.8 ± 15.1%; P < 0.001).

Maximal voluntary knee extensor (a) and elbow flexor (b) torque (N.m) obtained before (PRE, gray histograms) and 48 h (48 H, black histograms) after an eccentric (low or high load) or concentric (control) exercise. * P < 0.05 significant difference when compared with PRE value. † P < 0.05 significant difference when compared with Low load
Muscle shear modulus
The control load revealed no significant changes in shear modulus at any time point or joint angle (all P values > 0.704). In contrast, we observed a significant time × load × angle interaction on elbow flexors (P < 0.001). When compared to PRE, the shear modulus of the elbow flexors measured at 30 min was significantly higher at 160° for both low [+70.5 ± 44.3%; P < 0.001 (d = 1.91)] and high load [+153.9 ± 192.4%; P < 0.001 (d = 1.18)]; with a significant effect of load at this elbow angle (P < 0.001) (Fig. 3). However, the shear modulus measured at 30 min did not change when measured at 70° (all P values > 0.989) and 110° of elbow flexion (all P values > 0.349). For knee extensors, there was a significant time × load × angle interaction on shear modulus (P < 0.01). Post hoc test showed that shear modulus increased at 30 min in the most stretched position (110° of knee flexion) for low [+26.7 ± 19.1%; P < 0.001 (d = 0.89)] and high load [+79.4 ± 67.1%; P < 0.001 (d = 1.28)]. A significant increase in shear modulus at 30 min was also found at 90°, but only for high load [+64.7 ± 56.1%; P < 0.001 (d = 1.22)] (cf. Fig. 3). Additionally, the shear modulus at 30 min was higher for a high load than a low load at 110° of knee flexion (P < 0.001) (cf. Fig. 3). No significant increase in shear modulus was found at 30° for both loads (all P values > 0.987).

a Shear modulus of knee extensors (rectus femoris, vastus lateralis, and vastus medialis are pooled) obtained at three relaxed knee joint angles (30°, 90°, and 110°) for low and high loads, before (gray histograms, PRE) and 30 min (black histograms, 30 min) after eccentric exercise. b Shear modulus of elbow flexors (biceps brachii and brachialis values are pooled) obtained at three relaxed elbow joint angles (70°, 110°, and 160°) for control (bottom panel), low and high loads (upper panel), before and 30 min after eccentric exercise. * P < 0.05 significant difference when compared with PRE value. † P < 0.05 significant difference when compared with Low load
Relations between shear modulus changes and force loss
By pooling both low and high load, there was a strong correlation between the relative changes in elbow flexors and knee extensors shear modulus (mean of all synergist muscles) at 30 min and the relative decrease in peak isometric torque at 48 H (r = −0.80 and r = −0.82, for elbow flexors and knee extensors, respectively) (Fig. 4).

Significant correlations (P < 0.05) were found between the relative increase in muscle shear modulus (in % of pre-exercise value, %PRE) at 30-min post-exercise (30 min) and the relative decrease in peak voluntary torque 48 h after exercise (48 H) for elbow flexors (Elbow—blue circles) and knee extensors (Knee—red circles) (low and high loads are pooled) (color figure online)
Between-muscle differences
For both muscle groups, the index of increase in shear modulus was higher for the high than the low load (Fig. 5). There was a significant muscle × load interaction for elbow flexors (P = 0.04) showing a larger index of shear modulus increase in BB than BA for high load (P = 0.03), while no significant difference was found between these muscles for low load (P = 0.78; Fig. 5). For knee extensors, a main effect of muscle was observed (P < 0.017). The index of increase in shear modulus was higher for RF than both VL (P < 0.023) and VM (P < 0.017), while no significant difference was found between VL and VM (P = 0.786) (Fig. 5). These differences were observed regardless of the load (load effect: P = 0.604).

Index of increase (expressed as a percentage) in biceps brachii (BB, gray histograms), brachialis (BA, black histograms), vastus lateralis (VL, white histograms), vastus medialis (VM, white hatched histograms), and rectus femoris (RF, white histograms whit black dots) 30 min after an eccentric exercise (low and high loads). * P < 0.05 significant difference between synergist muscles. † P < 0.05 significant difference when compared with the Low load
Discussion
The present study has two main observations: (1) there was a high correlation between the decrease in force-generating capacity at 48-h post-eccentric exercise and the increase in muscle shear modulus measured 30-min post-exercise; (2) synergist muscles are not similarly affected by a damaging exercise with the biarticular muscles (BB and RF) more affected than their monoarticular synergists.
Overall, our results show that low and high eccentric loads induced a force deficit of about −18% (ranged from +2.0% to −45.4%) and −32% (ranging from −3.4% to −66.8%), respectively. These results allowed us to determine the relevance of the increase in shear modulus as a potential marker of the amount of muscle damage over a wide range of muscle force-generating capacity deficit.
The present study showed that an intense and non-damaging exercise (concentric contractions) does not change the shear modulus. This demonstrates that this measure is not influenced by metabolic factors originating from fatiguing contractions. Conversely, we showed a rise in muscle shear modulus 30 min after the damaging exercises [from +27% (knee extensors-low load) to +147% (elbow flexors-high load)]. This finding is in accordance with previous observations from our group and others (Green et al. 2012; Lacourpaille et al. 2014; Guilhem et al. 2016). Note that the mechanisms involved in this early rise in shear modulus remains unknown. It has been demonstrated that fluid accumulation and inflammation are not the determinant factors of this phenomenon (Lacourpaille et al. 2014). The initial damage of muscle fiber membranes and cytoskeleton triggers a significant loss of the cytoskeletal protein desmin 15 min after eccentric contractions (Lieber et al. 1996) and increase in resting calcium in muscle fibers (Balnave and Allen 1995, 1996). By connecting adjacent myofibrils to the Z-line, desmin plays a role in the transmission of tension from the contractile apparatus to the sarcolemma (Capetanaki et al. 2007). The desmin loss induced by muscle damage may thus affect tension transfer to the fiber membrane and should mainly contribute to decrease muscle stiffness. The increase in shear modulus is likely associated with the rapid calcium homeostasis perturbation subsequent to the myofibrillar disruptions induced by eccentric contractions (Whitehead et al. 2001; Chen et al. 2007). This interpretation is supported by the following observations. First, calcium homeostasis perturbation has been reported to be sufficient to trigger an augmentation of stable attached cross-bridges number (Whitehead et al. 2001) and/or the binding between titin and actin proteins (Herzog 2014). Second, we observed a large increase in shear modulus at long muscle lengths (+54 and +112%, for knee extensors and elbow flexors, respectively) while no changes were observed when the muscle was below the slack length. This result may reflect the increased sensitivity of muscle fibers to Ca2+ increases as muscle is elongated (Stephenson and Wendt 1984; Balnave and Allen 1996). Note that our approach provides only an indirect estimation of muscle damage. The direct relationship between changes in shear modulus and muscle damage remains to be established through an invasive approach that would quantify the proportion of disrupted fibers (Raastad et al. 2010) and assess sarcoplasmic Ca2+ regulation (Chen et al. 2007).
Our results showed that the relative increase in shear modulus measured at long muscle lengths 30 min after an eccentric exercise was strongly correlated to the force loss at 48 H (r ≥ −0.80; Fig. 4). It is noteworthy that this relationship does not seem linear over the entire range (cf. Fig. 4) but apparently level off at around 50% of decrease in peak voluntary torque. It is possible that the slope of the relationship depends of the amount of muscle damage. In case of severe muscle damage (i.e., >50% force reduction; Paulsen et al. 2012), an accumulation of leucocytes in the exercised muscles is systematically reported (Beaton et al. 2002; Paulsen et al. 2010). This accumulation might worsen the sarcolemmal damage (Proske and Morgan 2001). Alternatively, the changes in the angle of peak torque observed in the presence of muscle damage (Butterfield and Herzog 2005) might lead to an over- or underestimation of the isometric strength loss. As such, it would affect the slope of the relationship between the increase in muscle shear modulus and the strength loss at 48 H. Regardless the origin of this non-linear relationship, we can reasonably conclude that the greater the increase in shear modulus, the bigger the functional impairment. Additionally, considering that the decrease in maximal force-generating capacity is related to the number of disrupted fibers (Raastad et al. 2010), it seems reasonable to assume that our results indicate that the increase in muscle shear modulus reflects the amount of muscle damage. Interestingly this finding was observed subsequently for low and high eccentric loads in muscles with different susceptibility to damage, underlying its potential for sport settings and clinical practice.
The increase in muscle shear modulus demonstrated a muscle-length dependency with an absence of increase at a short muscle length (Fig. 3). As all the synergist muscles were not placed at the same relative length, the increase in shear modulus could not be used to compare the amount of damage between muscles. We therefore calculated an index of increase in muscle shear modulus that was independent of muscle length (Fig. 1b). This index was higher for high than low load exercises, for all the muscles (Fig. 5). In addition, the between synergistic muscles comparison revealed that the index of increase in shear modulus was higher for the biarticular rectus femoris and biceps brachii muscles than for the monoarticular vastii and brachialis muscles. This finding is in accordance with the larger edema (assessed using T2-MRI measurements) observed in rectus femoris than vastus lateralis and vastus medialis after repeated eccentric contractions (Prior et al. 2001; Paulsen et al. 2010). Similar results were reported for the biceps brachii that exhibited a higher T2 signal intensity change after a fast eccentric exercise than the brachialis (Kulig et al. 2001). This finding is likely explained by the predominance of fast twitch fibers in these muscles (rectus femoris and biceps brachii) (Johnson et al. 1973). Indeed, several studies in human (e.g., Chapman et al. 2013; Kulig et al. 2001; Paddon-Jones et al. 2005) suggest that type II fibers are more prone to damage, although none of them has demonstrated this relationship directly. In addition, other works showed that the rectus femoris experienced greater damage due to the longer muscle lengthening distance during seated eccentric knee extension compared to daily life multi-joints tasks (Takahashi et al. 1994; Prior et al. 2001). Additionally, the greater moment arm (Kawakami et al. 1994) combined with the longer fibers of the biceps brachii may result in larger biceps brachii fiber strain compared to brachialis during eccentric contractions.
In the present study, we considered the decrease in peak torque measured at 48 H during maximal voluntary isometric contractions as a surrogate of exercise-induced myofibrillar disruptions (Raastad et al. 2010). This is questionable as muscle damage is also associated with delayed soreness that might peak at 48 H. As experimental pain alone (i.e., without associated tissue damage) may be associated with reduced maximal voluntary contraction torque to an extent that can vary between individuals (Salomoni et al. 2016), soreness may have acted as a cofounding factor in our experiment. In addition, shear modulus was not measured in all the synergist muscles, with only ~80% (knee extension) and ~87% (elbow flexion) of the physiological cross sectional area considered (Kawakami et al. 1994; Farahmand et al. 1998). Together, this may explain why the correlation between force deficit at 48 H and increased shear modulus at 30 min is not perfect (Hopkins 2000). Despite these limitations, the relationship is considered as very high (Hopkins 2000) meaning that the shear modulus of the main agonist muscles is sufficient to provide a strong estimation of the amount of muscle damage.
The results of the present study may have important applications in musculoskeletal rehabilitation and sport practice. Indeed, muscle force decrease subsequent to eccentric exercise governs an individual’s ability to resume or sustain physical activity. As such, early, simple, and non-invasive detection of the force deficit may allow clinicians and coaches to adapt the timing and the intensity of the rehabilitation or training session following a damaging exercise. In addition, the accumulation of muscle damage throughout a sport season is thought be a key factor for muscle strain injury etiology in elite-level athletes (Opar et al. 2012). Therefore, further studies are needed to determine how the evaluation of muscle shear modulus may help coaches to adapt the training volume in the next days after damaging exercises and to determine if this approach could contribute to reduce the occurrence of muscle strain injury.
Conclusion
Using two different eccentric exercise loads, we observed a close relationship between a decrease in force-generating capacity 48-h post-eccentric exercise and early increase in muscle shear modulus in muscle groups more (elbow flexors) or less (knee extensors) prone to muscle damage. Therefore, the current study provides evidence that an increase in muscle shear modulus 30 min post-exercise can be considered as an early and non-invasive marker of the amount of muscle damage. In addition, calculation of the index of increase revealed that synergist muscles are not similarly affected by a damaging exercise, as evidenced by biarticular muscles (BB and RF) that are more affected than their monoarticular synergists. Overall, these results may have important applications in musculoskeletal rehabilitation and elite sport practice.
Abbreviations
- ANOVA:
-
Analysis of variance
- BA:
-
Brachialis
- BB:
-
Biceps brachii
- MVC:
-
Maximal voluntary contraction
- RF:
-
Rectus femoris
- SSI:
-
Supersonic shear imaging
- VL:
-
Vastus lateralis
- VM:
-
Vastus medialis
References
Agten CA, Buck FM, Dyer L et al (2016) Delayed-onset muscle soreness: temporal assessment with quantitative MRI and shear-wave ultrasound elastography. AJR Am J Roentgenol. doi:10.2214/AJR.16.16617
Balnave CD, Allen DG (1995) Intracellular calcium and force in single mouse muscle fibres following repeated contractions with stretch. J Physiol 488:25
Balnave CD, Allen DG (1996) The effect of muscle length on intracellular calcium and force in single fibres from mouse skeletal muscle. J Physiol 492:705–713
Beaton LJ, Tarnopolsky MA, Phillips SM (2002) Variability in estimating eccentric contraction-induced muscle damage and inflammation in humans. Can J Appl Physiol 27:516–526
Bercoff J, Tanter M, Fink M (2004) Supersonic shear imaging: a new technique for soft tissue elasticity mapping. Ultrason Ferroelectr Freq Control IEEE Trans 51:396–409
Bouillard K, Hug F, Guével A, Nordez A (2012a) Shear elastic modulus can be used to estimate an index of individual muscle force during a submaximal isometric fatiguing contraction. J Appl Physiol (1985) 113:1353–1361. doi:10.1152/japplphysiol.00858.2012
Bouillard K, Nordez A, Hodges PW et al (2012b) Evidence of changes in load sharing during isometric elbow flexion with ramped torque. J Biomech 45:1424–1429. doi:10.1016/j.jbiomech.2012.02.020
Butterfield TA, Herzog W (2005) Quantification of muscle fiber strain during in vivo repetitive stretch-shortening cycles. J Appl Physiol 99:593–602. doi:10.1152/japplphysiol.01128.2004
Capetanaki Y, Bloch RJ, Kouloumenta A et al (2007) Muscle intermediate filaments and their links to membranes and membranous organelles. Exp Cell Res 313:2063–2076. doi:10.1016/j.yexcr.2007.03.033
Chapman DW, Simpson JA, Iscoe S, Robins T, Nosaka K (2013) Changes in serum fast and slow skeletal troponin I concentration following maximal eccentric contractions. J Sci Med Sport 16:82–85
Chen W, Ruell PA, Ghoddusi M et al (2007) Ultrastructural changes and sarcoplasmic reticulum Ca2+ regulation in red vastus muscle following eccentric exercise in the rat: effects of eccentric exercise on SR Ca2+ regulation. Exp Physiol 92:437–447. doi:10.1113/expphysiol.2006.036442
Cohen JH (1988) Statistical power for the behavioral sciences. REMEDICA Publishing, London
Davis J, Kaufman KR, Lieber RL (2003) Correlation between active and passive isometric force and intramuscular pressure in the isolated rabbit tibialis anterior muscle. J Biomech 36:505–512. doi:10.1016/S0021-9290(02)00430-X
Eby SF, Song P, Chen S et al (2013) Validation of shear wave elastography in skeletal muscle. J Biomech 46:2381–2387. doi:10.1016/j.jbiomech.2013.07.033
Eby SF, Cloud BA, Brandenburg JE et al (2015) Shear wave elastography of passive skeletal muscle stiffness: influences of sex and age throughout adulthood. Clin Biomech 30:22–27. doi:10.1016/j.clinbiomech.2014.11.011
Eby S, Zhao H, Song P et al (2016) Quantitative evaluation of passive muscle stiffness in chronic stroke. Am J Phys Med Rehabil 95:899–910. doi:10.1097/PHM.0000000000000516
Farahmand F, Sejiavongse W, Amis AA (1998) Quantitative study of the quadriceps muscles and trochlear groove geometry related to instability of the patellofemoral joint. J Orthop Res 16:136–143
Green MA, Sinkus R, Gandevia SC et al (2012) Measuring changes in muscle stiffness after eccentric exercise using elastography: measuring muscle stiffness changes after eccentric exercise using mre. NMR Biomed 25:852–858. doi:10.1002/nbm.1801
Guilhem G, Doguet V, Hauraix H et al (2016) Muscle force loss and soreness subsequent to maximal eccentric contractions depend on the amount of fascicle strain in vivo. Acta Physiol 217:152–163. doi:10.1111/apha.12654
Herzog W (2014) Mechanisms of enhanced force production in lengthening (eccentric) muscle contractions. J Appl Physiol 116:1407–1417. doi:10.1152/japplphysiol.00069.2013
Hoang PD, Herbert RD, Gandevia SC (2007) Effects of eccentric exercise on passive mechanical properties of human gastrocnemius in vivo. Med Sci Sports Exerc 39:849–857. doi:10.1249/MSS.0b013e318033499b
Hopkins WG (2000) Measures of reliability in sports medicine and science. Sports Med 30:1–15
Jamurtas AZ, Theocharis V, Tofas T et al (2005) Comparison between leg and arm eccentric exercises of the same relative intensity on indices of muscle damage. Eur J Appl Physiol 95:179–185. doi:10.1007/s00421-005-1345-0
Johnson MA, Sideri G, Weightman D, Appleton D (1973) A comparison of fibre size, fibre type constitution and spatial fibre type distribution in normal human muscle and in muscle from cases of spinal muscular atrophy and from other neuromuscular disorders. J Neurol Sci 20:345–361
Kawakami Y, Nakazawa K, Fujimoto T et al (1994) Specific tension of elbow flexor and extensor muscles based on magnetic resonance imaging. Eur J Appl Physiol 68:139–147
Kulig K, Powers CM, Shellock FG, Terk M (2001) The effects of eccentric velocity on activation of elbow flexors: evaluation by magnetic resonance imaging. Med Sci Sports Exerc 33:196–200
Lacourpaille L, Hug F, Bouillard K et al (2012) Supersonic shear imaging provides a reliable measurement of resting muscle shear elastic modulus. Physiol Meas 33:N19–28. doi:10.1088/0967-3334/33/3/N19
Lacourpaille L, Nordez A, Hug F (2013) Influence of stimulus intensity on electromechanical delay and its mechanisms. J Electromyogr Kinesiol Off J Int Soc Electrophysiol Kinesiol 23:51–55. doi:10.1016/j.jelekin.2012.06.010
Lacourpaille L, Nordez A, Hug F et al (2014) Time-course effect of exercise-induced muscle damage on localized muscle mechanical properties assessed using elastography. Acta Physiol 211:135–146. doi:10.1111/apha.12272
Lieber R, Thornell E, Friden J (1996) Muscle cytoskeletal disruption occurs within the first 15 min of cyclic eccentric contraction. J Appl Physiol 80:278–284
Opar MDA, Williams MD, Shield AJ (2012) Hamstring strain injuries. Sports Med 42:209–226
Paddon-Jones D, Keech A, Lonergan A, Abernethy P (2005) Differential expression of muscle damage in humans following acute fast and slow velocity eccentric exercise. J Sci Med Sport 8:255–263
Paulsen G, Crameri R, Benestad HB, Fjeld JG, Morkrid L, Hallen J, Raastad T (2010) Time course of leukocyte accumulation in human muscle after eccentric exercise. Med Sci Sports Exerc 42:75–85
Paulsen G, Mikkelsen UR, Raastad T, Peake JM (2012) Leucocytes, cytokines and satellite cells: what role do they play in muscle damage and regeneration following eccentric exercise. Exerc Immunol Rev 18:42–97
Prior BM, Jayaraman RC, Reid RW et al (2001) Biarticular and monoarticular muscle activation and injury in human quadriceps muscle. Eur J Appl Physiol 85:185–190. doi:10.1007/s004210100434
Proske U, Morgan DL (2001) Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications. J Physiol 537:333–345
Raastad T, Owe SG, Paulsen G et al (2010) Changes in calpain activity, muscle structure, and function after eccentric exercise. Med Sci Sports Exerc 42:86–95. doi:10.1249/MSS.0b013e3181ac7afa
Salomoni S, Tucker K, Hug F et al (2016) Reduced maximal force during acute anterior knee pain is associated with deficits in voluntary muscle activation. PLoS One 11:e0161487. doi:10.1371/journal.pone.0161487
Stephenson D, Wendt I (1984) Length dependence of changes in sarcoplasmic calcium concentration and myofibrillar calcium sensitivity in striated muscle fibres. J Muscle Res Cell Motil 5:243–272
Takahashi H, Kuno S, Miyamoto T et al (1994) Changes in magnetic resonance images in human skeletal muscle after eccentric exercise. Eur J Appl Physiol 69:408–413
Warren GL, Lowe DA, Armstrong RB (1999) Measurement tools used in the study of eccentric contraction-induced injury. Sports Med 27:43–59. doi:10.2165/00007256-199927010-00004
Whitehead NP, Weerakkody NS, Gregory JE et al (2001) Changes in passive tension of muscle in humans and animals after eccentric exercise. J Physiol 533:593–604
Acknowledgements
This study was supported by grants from the French Ministry of Sports (Contract No. 15i19) and the Région Pays de la Loire (QUETE Project No. 2015-09035).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
No conflicts of interest, financial or otherwise, are declared by the authors. The results of the study are presented clearly, honestly, and without fabrication, falsification, or inappropriate data manipulation. The results of the present study do not constitute endorsement by the American College of Sports Medicine.
Additional information
Communicated by Olivier Seynnes.
Rights and permissions
About this article

Cite this article
Lacourpaille, L., Nordez, A., Hug, F. et al. Early detection of exercise-induced muscle damage using elastography. Eur J Appl Physiol 117, 2047–2056 (2017). https://doi.org/10.1007/s00421-017-3695-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-017-3695-9