
Abstract
The aim of this study was to investigate the effects of acute caffeine ingestion on intermittent high-intensity sprint performance after 5 days of creatine loading. After completing a control trial (no ergogenic aids, CON), twelve physically active men were administered in a double-blind, randomized crossover protocol to receive CRE + PLA (0.3 g kg−1 day−1 of creatine for 5 days then followed by 6 mg kg−1 of placebo) and CRE + CAF (0.3 g kg−1 day−1 of creatine for 5 days and followed by 6 mg kg−1 of caffeine), after which they performed a repeated sprint test. Each test consisted of six 10-s intermittent high-intensity sprints on a cycling ergometer, with 60-s rest intervals between sprints. Mean power, peak power, rating of perceived exertion (RPE), and heart rates were measured during the test. Blood samples for lactate, glucose, and catecholamine concentrations were drawn at specified intervals. The mean and peak power observed in the CRE + CAF were significantly higher than those found in the CON during Sprints 1 and 3; and the CRE + CAF showed significantly higher mean and peak power than that in the CRE + PLA during Sprints 1 and 2. The mean and peak power during Sprint 3 in the CRE + PLA was significantly greater than that in the CON. Heart rates, plasma lactate, and glucose increased significantly with CRE + CAF during most sprints. No significant differences were observed in the RPE among the three trials. The present study determined that caffeine ingestion after creatine supplements augmented intermittent high-intensity sprint performance.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A wide variety of supplementary products are used by athletes to enhance performance. Though not new to the arena of sports science, creatine monohydrate is currently one of the most widely utilized supplements by those seeking to improve athletic performance (Buford et al. 2007). Briefly, creatine supplementation improves performance by enhancing cellular homeostasis and buffering acidosis, resulting from exercise. This is accomplished through the utilization of hydrogen ions during the creatine kinase reaction, and through rephosphorylation of adenosine diphosphate (ADP) into adenosine triphosphate (ATP) (Bemben and Lamont 2005). Vandenberghe et al. (1996) suggested that intramuscular creatine and phosphocreatine (PCr) levels might be even further elevated by adding a direct stimulus to the membrane creatine transport system during administration of creatine. Direct stimulation would have to include the creatine transporter in the sarcolemma (Greenhaff et al. 1994), the influence of hormones such as catecholamine or insulin (Snow and Murphy 2001), and Na+-K+- adenosine triphosphatase (Clausen and Flatman 1977). A crucial fact to understand is that the sarcolemmic creatine transport is known to be critically dependent on extracellular Na+ gradient (Loike et al. 1986).
Brosnan and Brosnan (2007) noted that a specific creatine transporter can be detected in both skeletal muscle and liver tissue, and can be activated by Na+-K+-adenosine triphosphatase, which could presumably improve the uptake of creatine by anabolic hormones, such as insulin. In their study, anabolic hormones appeared to increase concentrations of creatine and creatine transport levels; whereas catabolic hormones (for example, catecholamine) improved the utilization of creatine in an anaerobic system by means of creatine transporter. For this reason, adrenergic stimulation might increase the driving force behind the delivery of creatine to muscle tissue via Na+-K+-adenosine triphosphatase pump activity, while raising Na+ sarcolemmic gradients (Clausen 1996, 2003). Caffeine (1,3,7-trimethylxanthine) may perform intracellular actions such as stimulating the secretion of adrenalin (Jackman et al. 1996). Increased adrenalin levels can induce a series of secondary metabolic reactions to produce ergogenic action (Stuart et al. 2005) and increase concentrations of lactate in the blood (Anselme et al. 1992; Greer et al. 1998) during the performance of exercise. In addition, the chemical structure of caffeine is very similar to that of adenosine, and is able to block the action of adenosine by binding with the receptors of the cell membrane (Graham 2001).
Vandenberghe et al. (1996) hypothesized that ergogenic action in performance might be elevated through combined supplementation of creatine with caffeine. Interestingly, they found that when creatine is ingested simultaneously with caffeine, the ergogenic benefit of creatine is lost, even though the level of intramuscular creatine is increased. Subsequent studies (Hespel et al. 2002; Vanakoski et al. 1998) have reported similar results in which the simultaneous ingestion of creatine with caffeine could interfere with the ergogenic effects of creatine. However, Doherty et al. (2002) investigated the effect of acute ingestion of caffeine on treadmill running until exhaustion at an exercise intensity equivalent to 125% \( \dot{V}{\text{O}}_{2\max } \), after a period of creatine supplementation with abstinence from caffeine. They found that total oxygen consumption and running time on a treadmill were significantly higher for those subjects who had been administered creatine and caffeine than for subjects administered a placebo. Although Doherty et al. (2002) suggested that ingesting creatine and caffeine at different times may inhibit opposition effects; no evidence exists to clarify whether or not such a supplemental regime could facilitate a “stop-and-go” type of exercise performance.
Therefore, the purpose of this study was to determine the effect of acute ingestion of caffeine 1 h prior to performing an intermittent high-intensity sprint (IHIS) exercise after pre-loading (i.e., ingesting) creatine for 5 days. We hypothesized that the acute ingestion of caffeine after short-term creatine supplementation would increase both anaerobic performance and lactate concentrations during repeated sprint cycling exercises, and creatine-only supplementation would contribute to anaerobic energy compared with no ergogenic aid during the stop-and-go exercise.
Methods
Participants
Twelve physically active men (age 19 ± 0.6 years, height 174.2 ± 7.4 cm, and body mass 69.9 ± 6.5 kg) were recruited for this study, which was approved by the National Taiwan Sport University Ethics Committee. All participants received written and verbal instructions regarding the medicine to be ingested during the study, including the potential side effects. Subjects were also asked to complete an ergogenic aids history questionnaire. Prior to the first trial, the questionnaire results showed that the participants had not taken any ergogenic aids or medicines in the previous 3 months, and that their volume of caffeine ingestion was lower than 50 ppm day−1. Moreover, they were required to abstain from caffeine-containing products and record physical activity (that is, exercise duration and rating of perceived exertion in exercise intensity) during the period of creatine loading, and were asked to avoid altering their level of physical activity. Subjects were also asked to abstain from exercise for 24 h and to fast at least 4 h prior to visiting the laboratory to reduce the effects of exercise and food interference.
Experimental design
A double-blind, randomized crossover, and counterbalancing experimental design was used in this study. Each participant visited the laboratory on four separate occasions. The purpose of the first visit was to familiarize participants with the equipment and the 6 × 10-s exercise protocol. Visit 2 was the control (CON) trial without any ergogenic aids, which included the 6 × 10-s IHIS test and taking blood samples during the IHIS test. The experimental procedure in visit 3 was similar to visit 4: following creatine loading for 5 days, participants ingested caffeine (CRE + CAF) or a placebo (CRE + PLA) 1 h, before the 6 × 10-s IHIS test on the sixth day.
A standardized testing process was used (Fig. 1). When participants arrived at the laboratory, body weight (BW) was measured to determine the load of the IHIS test on a cycling ergometer and for analysis of variations in BW after 5 days of creatine loading. Blood samples were drawn from each participant’s earlobe at baseline (pre-test), during exercise, and post-test for plasma lactate and glucose analysis; and blood samples were drawn from the forearm vein at baseline (pre-test), post-caffeine ingestion, and post-test for plasma epinephrine (Ep) and plasma norepinephrine (NEp) analysis.

Schematic of the exercise and blood samples collection timeline. Down arrow plasma lactate and glucose. Open square body weight measurement. Concentric circle epinephrine and norepinephrine. IHIS intermittent high-intensity sprints
Heart rate (HR) was estimated by the Polar heart rate monitor (Polar S810iTM, Polar Electro Inc, Finland) during the IHIS test, and the 6-20 Borg rating of perceived exertion (RPE) scale was also obtained immediately after every 10-s sprint throughout the IHIS test. The RPE scale was applied in the study due to the reliable linear response during high-intensity exercise (Graham et al. 2000) and the intermittent sprint test (Spriet et al. 1992).
Intermittent high-intensity sprint test
One hour after the caffeine or placebo had been ingested, participants performed a standardized warm-up session comprising 4 min of sub-maximal cycling at 50 W and three repetitions of brief unloaded sprints (5 s), followed by a 5-min routine stretching exercise. Subjects were asked to remain continuously seated on a cycling ergometer (Avantronic Cyclus II, h/p Cosmos®, Germany) during the IHIS test to prevent the recruitment of the other muscle groups. During the test, all participants were encouraged to pedal as fast as possible against the given load for every sprint on the cycling ergometer. The load of the IHIS test was calculated from BW by the equation of [(0.7 × BW)/0.173], which was designed within the Cyclus II ergometer program for inducing near-maximal power output of the active male population in our study. The power output data were then transferred to a personal computer for calculation of the peak and mean power. The IHIS test in this study consisted of 6 × 10-s sprints, with 60 s of active recovery on a cycling ergometer. This regimen was designed to repeatedly stress the PCr/ATP system (Balsom et al. 1995; Yquel et al. 2002). During the 60-s recovery phases, participants pedaled at 55–60 rpm, against the load of 50 watts, for the active recovery, and were instructed to prepare for the next sprint during the last 10-s interval. Consistent verbal encouragement and appropriate feedback were offered during each sprint, but no performance information was provided.
Supplementation
Subjects were asked to prepare for each testing session in a similar manner and to refrain from consuming alcoholic beverages for 24 h prior to the experiment. Five days before visits 3 and 4, participants ingested 0.3 g kg−1 day−1 of creatine (Phosphagen HPTM, EAS, Golden, CO) and the dosage was spread out over four times per day (that is, 0.075 g/kg per dose) for 5 days. Subjects were instructed to mix the creatine powder in warm water (~250 ml) for better dissolution (Harris et al. 1992) and to ingest the solution with their morning, mid-day, afternoon, and evening meals. It has been well established that this creatine supplementary method is effective for increasing the contents of creatine in the muscle of young males (Harris et al. 1992; Preen et al. 2001). During the period of creatine supplementation, all participants were required to refrain from drinking caffeinated beverages (Doherty et al. 2002) and to keep a detailed record of their dietary intake. The aim of the five-day dietary record was to determine whether or not the participants ingested caffeine-containing products. After 5 days of creatine supplementation, participants were asked to consume either 6 mg kg−1 of caffeine (Sigma–Aldrich, Sydney, Australia) or an equivalent placebo (maltodextrin; Starmax Nutrition, Hereford, UK) 1 h before the IHIS test on the sixth day. Previous studies (Anselme et al. 1992; Spriet et al. 1992) have proved that this dosage of caffeine ingestion can elicit a positive ergogenic effect without serious side effects. In addition, visits 3 and 4 were separated by 1 month to allow a period for the washout of creatine, which was suggested in a previous study (Hultman et al. 1996).
Blood sampling and analysis
At the beginning of each trial, a catheter was placed percutaneously into a medial antecubital vein, and a normal saline drip (0.9% NaCl) was injected to maintain catheter patency to obtain 10 ml venous blood samples. The samples were taken while the participants were at rest, 60 min after ingesting caffeine or placebo capsules, and again at the post-test. The blood samples for catecholamine concentrations were drawn into tubes containing ethylenediaminetetraacetic acid, and centrifuged at 3,000 rpm. The plasma was stored at 2–8°C before immediate analysis of the plasma E and NEp concentrations via high performance liquid chromatography (HPLC) CLC 300 (Chromsystem®, München, Germany), following the method of Hue et al. (2000). In brief, the catecholamines were extracted by selective absorption using aluminum oxide (Chromsystem® HPLC Kit, Waters Corp, Milford, MA, USA) before the HPLC was run. Aluminum oxide was shaken briefly in an extraction buffer (50 μl) and 1 ml of plasma was added with a 50 μl internal standard solution (600 pg dihydroxybenzylamine). The aluminum oxide was washed three times, with a brief centrifugation between washes. The catecholamine was extracted with 120 μl elution buffers through brief shaking and subjecting it to a final centrifugation at 2,000g for 1 min, at which point 50 μl of the sample eluent was injected into a HPLC column and eluted with a mobile phase.
Capillary blood samples (10 μl) were taken from each the earlobe of each participant at pre-test, immediately after each cycling sprint, and 5 min after the IHIS test (post-test), for analysis of lactate and glucose concentrations using the Biosen C-Line Sport Analyser (EKF Diagnostics, Magdeburg, Germany) (Fig. 1).
Statistical analysis
Throughout the exercise, mean and peak power values were analyzed using two-way ANOVA with repeated measures for conditions (CON, CRE + CAF, and CRE + PLA) and time (sprint number). Plasma lactate, glucose, catecholamine, heart rate, and RPE were analyzed in the same manner. One-way ANOVA with repeated measures was used to compare body mass among the three conditions, and a paired t test was used to analyze physical activity of the 5 days between the CRE + CAF and CRE + PLA conditions. Statistical significance was set at a P value of ≤0.05 in all cases. Bonferroni post hoc analysis was used when the interaction was significant. The SPSS software package was used for statistical analysis (SPSS for Windows 11.0, SPSS, Inc., Chicago IL, USA). Data were expressed as means ± standard deviation (SD).
Results
Physical characteristics
Body mass was not different among the conditions of CON, CRE + CAF, and CRE + PLA (69.96 ± 6.58 vs. 70.15 ± 6.43 vs. 70.55 ± 6.29 kg; P > 0.05). No significant differences were noted with regard to physical activities between CRE + CAF (2.3 h day−1) and CRE + PLA (2.7 h day−1) during the period of creatine supplementation (P > 0.05). In addition, no caffeine-containing foods were detected in the dietary records during creatine supplementation. After all participants had ingested the caffeine capsules and completed the IHIS test, five of the participants experienced a transient, mild sense of anxiety; six participants exhibited slight trembling, and one participant experienced slight nausea.
High-intensity intermittent performance
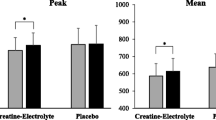
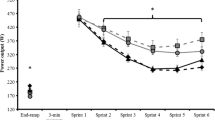
The effects of caffeine supplementation on the mean power and peak power of the IHIS test are respectively presented in Figs. 2 and 3. A significant interaction was observed for mean power (F = 5.03, P < 0.05). Post hoc analysis of the interaction indicated that CRE + CAF was significantly higher, 0.52 watt kg−1 during Sprint 1 and 0.72 watt kg−1 during Sprint 2 compared with CRE + PLA (P < 0.05). When compared with CON, CRE + CAF resulted in a 0.88 watt kg−1 increase during Sprint 1 and 0.52 watt kg−1 increase during Sprint 3 (P < 0.01). A significant increase of 0.41 watt kg−1 was observed in CRE + PLA relative to CON during Sprint 3 (P < 0.05) (Fig. 2).

The mean power of the 6 × 10-s intermittent high-intensity sprints. *CRE + CAF is significantly different from CON (P < 0.05); †CRE + CAF is significantly different from CRE + PLA (P < 0.05); #significant difference between CRE + PLA and CON (P < 0.05). Values are presented as mean ± SD

The peak power of the 6 × 10-s intermittent high-intensity sprints. *CRE + CAF is significantly different from CON (P < 0.05); †CRE + CAF is significantly different from CRE + PLA (P < 0.05); #significant difference between CRE + PLA and CON (P < 0.05). Values are presented as mean ± SD
A significant interaction was observed for peak power (F = 2.87, P < 0.05). Compared with the CRE + PLA, CRE + CAF showed a significant increase of 0.42 watt kg−1 and 0.58 watt kg−1 during Sprint 1 and Sprint 2 (P < 0.05), respectively. CRE + CAF also resulted in a 0.77 watt kg−1 increase during Sprint 1 and a 0.55 watt kg−1 increase during Sprint 3 relative to CON (P < 0.05). Additionally, CRE + PLA showed a significant increase by 0.40 watt kg−1 during Sprint 3 compared with CON (P < 0.05) (Fig. 3).
Plasma lactate and glucose concentrations
A significant main effect of condition and time (P < 0.01) for plasma lactate and glucose was detected; however, no significant interaction was observed (F = 1.49, P > 0.05). Plasma lactate and glucose concentrations were higher for CRE + CAF compared to CRE + PLA and CON for most of the sprints and the post-test (Table 1). Plasma lactate and glucose concentrations in the CRE + PLA did not significantly differ from the CON (P > 0.05) (Table 1).
Catecholamines
No significant interaction was found in plasma Ep (F = 0.15, P > 0.05), but a significant interaction was observed in plasma NEp (F = 5.48, P < 0.05). Figure 4 shows that no significant differences existed between CRE + CAF and CRE + PLA in plasma Ep and NEp at any point (P > 0.05). In addition, no significant difference in the time course on plasma Ep levels was apparent (P > 0.05). However, caffeine ingestion resulted in significantly higher concentrations of NEp in the pre-test and in post-test compared to levels measured 1 h after caffeine ingestion (P < 0.05) (Fig. 4).

Plasma epinephrine (Ep) and norepinephrine (NEp) concentrations in the CRE + CAF and CRE + PLA at the pre-test, post-caffeine ingestion, and after the 6 × 10-s intermittent high-intensity sprints test. The Ep and NEp are not significantly different between CRE + CAF and CRE + PLA at every point. #The NEp in CRE + CAF during pre-test and post-test is significantly greater than post-caffeine ingestion (P < 0.05). Values are mean ± SD
Rating of perceived exertion (RPE)
No significant interaction for RPE scores was noted (F = 1.99, P > 0.05). No significant differences were noted in the RPE measured after each sprint between CON, CRE + CAF and CRE + PLA during the IHIS test (P > 0.05). The results of RPE analysis indicated that caffeine ingestion did not affect the level of perceived effort by the athletes during the IHIS test.
Heart rate
During the intermittent sprints, peak heart rates in CRE + CAF from Sprint 3 to Sprint 5 were significantly higher than those in CON and CRE + PLA (P < 0.05) (Table 2). Compared with the CON and CRE + PLA, the mean heart rate also significantly increased in the CRE + CAF throughout the IHIS test (CRE + CAF vs. CON vs. CRE + PLA; 158 ± 9 vs. 151 ± 12 vs. 152 ± 15 bpm, P < 0.05) (Table 2).
Discussion
This was the first study to investigate the acute effects of caffeine ingestion after short-term creatine loading on the 6 × 10-s IHIS performance. The main findings of the present study showed that after 5 days of creatine loading (0.3 g kg−1), a single dose (6 mg kg−1) of caffeine ingestion increased the peak and mean power outputs of the IHIS. In addition, short-term creatine supplementation (CRE + PLA) resulted in greater mean and peak power than control. However, caffeine might also augment metabolic and cardiovascular stress during exercise; although the perceptions of participants regarding the efforts they expended were unaltered.
A limitation of this study is that the control condition was not randomized along with the CRE + PLA and CRE + CAF conditions, because we consider that another washout period would have been required and this was not feasible given the possibility that fitness level of the participants could change over a longer experimental period.
Our results provide further evidence to support the assertion that the ergogenic effects of creatine on high-intensity performance are not negatively influenced by acute caffeine ingestion. This determinate finding is similar to that of a previous study (Doherty et al. 2002). Additionally, Izquierdo et al. (2002) and Ziegenfuss et al. (1998) have demonstrated that BW did not increase in conjunction with creatine supplementation, which was likely due to participants engaging in regular physical activity. Conversely, a single dose of caffeine ingestion did not interfere with the effects of short-term creatine loading on performance. This result did not correspond with previous studies (Hespel et al. 2002; Vanakoski et al. 1998). Although the effects of caffeine ingestion on short-term, intense exercise are not well understood, previous studies have documented that maximal anaerobic power (Anselme et al. 1992) and intermittent sprinting ability (Schneiker et al. 2006) were enhanced by caffeine ingestion. This was supported by the results of our study.
Graham et al. (2000) had subjects perform 1 h of exercise at the intensity of approximately 70% \( \dot{V}{\text{O}}_{2\max } \) after ingesting caffeine, the results of which indicated that caffeine could alter blood lactate concentrations during exercise. In our study, the ingestion of caffeine increased peak and mean power output in the first three sprints; however, no differences in peak or mean power was noted between Sprint 4 and Sprint 6. It is possible that the improvements in IHIS performance induced by caffeine ingestion could result from increased levels of intracellular Ca+ to facilitate excitation–contraction coupling in the skeletal muscles (Ogawa 1994) or an increase in the sarcolemmic creatine transporter levels (Loike et al. 1986). Additionally, Williams (1991) indicated that caffeine ingestion could function directly on the central nervous system (CNS) by inhibiting the enzyme phosphodiesterase (PDE), which is responsible for the degradation of the second messenger, cyclic-3′,5′-adenosine monophosphate (cyclic AMP). By inhibiting PDE mediated breakdown, cyclic AMP levels remain elevated, resulting in increased neurotransmitter release, neuronal activation, and subsequent stimulation of the CNS (Sutherland and Rall 1958). The increases in peak/mean power output in the CRE + PLA in Sprint 3 might be attributed to augmentation of creatine and PCr content in the skeletal muscles (Harris et al. 1992; Pulido et al. 1998); however, the IHIS performance was not enhanced by creatine compared with CON during other sprints. A similar finding was reported by McKenna et al. (1999), who observed that peak power, or cumulative work output, had small increases of <2–3% after creatine supplementation during a 5 × 10-s intermittent sprint test protocol, despite the observation that muscle total creatine content and PCr were elevated significantly in the creatine group. These results correspond with other studies (Izquierdo et al. 2002; Wright et al. 2007; Ziegenfuss et al. 2002); suggesting that short-term creatine loading appears to significantly affect intermittent high-intensity performance. The benefits of creatine loading on intermittent sprint performance are not clear, due to the differences in experimental models used in other studies (Balsom et al. 1995; Glaister et al. 2006), such as double-blind fashion and crossover design. This area deserves future consideration.
Although the ingestion of caffeine improved IHIS performance, metabolic and cardiovascular stress was also increased due to an increase in plasma lactate levels and heart rate. However, the present study noted no significant differences in RPE among the three trials during the 6 × 10-s IHIS test. In a previous study, caffeine was found to be effective for improving cycling performance and lowering perceptual response, specifically characterized by a constant 100% maximal power output for 2 min, followed by sprinting for 1 min (Doherty et al. 2004); however, those results were not similar to ours, or to those of another study (Schneiker et al. 2006). Therefore, caffeine might increase tolerance to high-intensity exercise because of the improvements in intermittent sprint performance, without causing any change in the perception of effort. In other words, caffeine ingestion may have no impact on motor or sensory pathways during performance of intermittent sprint exercise.
Caffeine ingestion can cause an increase in heart rate during exercise (Bell et al. 1998). The strong sympathetic stimulus with caffeine ingestion may cause an increase in plasma Ep at exhaustion (Spriet et al. 1992; Van Soeren and Graham 1998). Although the Ep and NEp registered no significant differences between the conditions in the present study, the NEp concentrations in CRE + CAF at post-test were higher than concentrations after caffeine ingestion (Fig. 4). An alternative explanation for the changes in NEp concentration in CRE + CAF at post-test might be due to the relationship between short-term, high-intensity exercise and increases in the level of plasma catecholamines. However, the single dose (6 mg kg−1) of caffeine ingested in our study might not be adequate to induce observable differences from the ingestion of the placebo. Interpreting the results of the plasma Ep and NEp in our study was difficult due to the large variations observed, and higher plasma catecholamines levels appeared unlikely to have been a major factor in enhancing performance (Collomp et al. 1991; Greer et al. 1998). Thus, further research is required to determine the relationship between caffeine-induced changes in catecholamines and the IHIS exercise.
Study data indicate that acute caffeine ingestion after short-term creatine loading altered the mobilization of plasma lactate and glucose (Table 1). As previously demonstrated, caffeine intake in humans can increase blood glucose concentrations (Doherty et al. 2002; Raguso et al. 1996). Raguso et al. (1996) determined that the appearance rate of glucose was not influenced by theophylline, with subjects at rest or during 30 min of exercise at 75% \( \dot{V}{\text{O}}_{2\max } \), but the disappearance rate of glucose was decreased. They concluded that this phenomenon was related to lower glucose uptake by active muscles. However, the exact mechanism of the lowering glucose uptake of muscle cells reaction requires additional research and investigation for a more complete understanding.
The present study revealed that plasma lactate levels measured during the interval between Sprints 1 and 4, 5 min after the IHIS test, in the CRE + CAF trial, were significantly greater than those measured in the CRE + PLA trial. These results are in agreement with the other studies (Anselme et al. 1992; Collomp et al. 1992; Crowe et al. 2006; Doherty et al. 2002). Anselme et al. (1992) reported that ingesting caffeine increased the concentration of blood lactate at the completion of, and 5 min after, 4 × 6-s cycling sprints, and indicated that this could be related to the levels of Ca+. This further suggests that Ca+ released from the sarcoplasmic reticulum during the contraction of muscles probably activated the enzymatic transformation of the glycogen phosphorylase β to the more active form α, thereby accelerating glycogenolysis. Another possible explanation for the increase in plasma lactate concentrations might be that the duration of the rest interval (60 s) during the 6 × 10-s sprints was not long enough for the liver to metabolize lactate and/or intramuscular mitochondria when ingesting caffeine. Greer et al. (1998) suggested that caffeine ingestion enhanced performance during the initial efforts, and might be ergolytic as fatigue developed, possibly due to an increase in by-products of anaerobic metabolism. Furthermore, the observed accumulation of plasma lactate might have been the result of a deficient oxidation of pyruvate in the skeletal muscles (Graham 2001) or due to muscular glycogenolysis (Holloszy 1982). Crowe et al. (2006) suggested that elevated blood lactate concentrations could be the result of changes in lactate clearance rather than production, and did not necessarily reflect an increase in glycolytic metabolism or ergogenic effects of caffeine. Graham et al. (2000) directly proved that caffeine ingestion increased the levels of arterial lactate during exercise, but the levels of both muscle lactate and the blood lactate released from the exercising leg were unaltered. Hence, the ingestion of creatine plus caffeine resulting in increased blood lactate levels could possibly be related to the inhibition of lactate clearance, because no significant differences in the catecholamine levels between the treatments were present in this study.
In summary, this study indicated that after short-term creatine supplementation with caffeine abstinence, the ingestion of a moderately high dose of caffeine 1 h before exercise could improve the subsequent intermittent high-intensity cycling sprint performance, but might stimulate the metabolic responses in plasma lactate and glucose. This study also determined that caffeine might not attenuate the ergogenic benefits of creatine, if the ingestion of caffeine occurred after the creatine loading phase. The practical recommendation of this study is to supplement with caffeine before intense exercise, if coaches or athletes would like to augment the ergogenic effects of creatine. However, one should consider the resulting increase in the accumulation of lactate in the blood and elevation of heart rate during intermittent sprint exercises.
References
Anselme F, Collomp K, Mercier B, Ahmaidi S, Ch Prefaut (1992) Caffeine increases maximal anaerobic power and blood lactate concentration. Eur J Appl Physiol Occup Physiol 65:188–191
Balsom PD, Soderlund K, Sjodin B, Ekblom B (1995) Skeletal muscle metabolism during short duration high-intensity exercise: influence of creatine supplementation. Acta Physiol Scand 154:303–310
Bell DG, Jacobs I, Zamecnik J (1998) Effects of caffeine, ephedrine and their combination on time to exhaustion during high-intensity exercise. Eur J Appl Physiol Occup Physiol 77:427–433
Bemben MG, Lamont HS (2005) Creatine supplementation and exercise performance: recent findings. Sports Med 35:107–125
Brosnan JT, Brosnan ME (2007) Creatine: endogenous metabolite, dietary, and therapeutic supplement. Annu Rev Nutr 27:241–261
Buford TW, Kreider RB, Stout JR, Greenwood M, Campbell B, Spano M, Ziegenfuss T, Lopez H, Landis J, Antonio J (2007) International Society of Sports Nutrition position stand: creatine supplementation and exercise. J Int Soc Sports Nutr 4:6
Clausen T (1996) The Na+, K+ pump in skeletal muscle: quantification, regulation and functional significance. Acta Physiol Scand 156:227–235
Clausen T (2003) Na+-K+ pump regulation and skeletal muscle contractility. Physiol Rev 83:1269–1324
Clausen T, Flatman JA (1977) The effect of catecholamines on Na+-K+ transport and membrane potential in rat soleus muscle. J Physiol Lond 270:383–414
Collomp K, Ahmaidi S, Audran M, Chanal JL, Prefaut CH (1991) Effects of caffeine ingestion on performance and anaerobic metabolism during the Wingate test. Int J Sports Med 12:439–443
Collomp K, Ahmaidi S, Chatard JC, Audran M, Ch Prefaut (1992) Benefits of caffeine ingestion on sprint performance in trained and untrained swimmers. Eur J Appl Physiol Occup Physiol 64:377–380
Crowe MJ, Leicht AS, Spinks WL (2006) Physiological and cognitive responses to caffeine during repeated high-intensity exercise. Int J Sport Nutr Exerc Metab 16:528–544
Doherty M, Smith PM, Davison RC, Hughes MG (2002) Caffeine is ergogenic after supplementation of oral creatine monohydrate. Med Sci Sports Exerc 34:1785–1792
Doherty M, Smith P, Hughes M, Davison R (2004) Caffeine lowers perceptual response and increases power output during high-intensity cycling. J Sports Sci 22:637–643
Glaister M, Lockey RA, Abraham CS, Staerck A, Goodwin JE, Mclnnes G (2006) Creatine supplementation and multiple sprint running performance. J Strength Cond Res 20:273–277
Graham TE (2001) Caffeine and exercise: metabolism, endurance and performance. Sports Med 31:785–807
Graham TE, Helge JW, MacLean DA, Kiens B, Richter EA (2000) Caffeine ingestion does not alter carbohydrate or fat metabolism in human skeletal muscle during exercise. J Physiol 529:837–847
Greenhaff PL, Bodin K, Siiderlund K, Hultman E (1994) Effect of oral creatine supplementation on skeletal muscle phosphocreatine resynthesis. Am J Physiol 266:E725–E730
Greer F, McLean C, Graham TE (1998) Caffeine, performance, and metabolism during repeated Wingate exercise tests. J Appl Physiol 85:1502–1508
Harris RC, Soderlund K, Hultman E (1992) Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation. Clin Sci (Lond) 83:367–374
Hespel P, Op’t Eijnde B, Van Leemputte M (2002) Opposite actions of caffeine and creatine on muscle relaxation time in humans. J Appl Physiol 92:513–518
Holloszy JO (1982) Muscle metabolism during exercise. Arch Phys Med Rehabil 63:231–234
Hue O, Le Gallais D, Boussana A, Galy O, Chamari K, Mercier B, Prefaut C (2000) Catecholamine, blood lactate and ventilatory responses to multi-cycle-run blocks. Med Sci Sports Exerc 32:1582–1586
Hultman E, Soderlund K, Timmons JA, Cederblad G, Greenhaff PL (1996) Muscle creatine loading in men. J Appl Physiol 81:232–237
Izquierdo M, Ibanez J, Gonzalez-Badillo JJ, Gorostiaga EM (2002) Effects of creatine supplementation on muscle power, endurance, and sprint performance. Med Sci Sports Exerc 34:332–343
Jackman M, Wendling P, Friars D, Graham TE (1996) Metabolic, catecholamine, and endurance responses to caffeine during intense exercise. J Appl Physiol 81:1658–1663
Loike JD, Somes M, Silverstein SC (1986) Creatine uptake, metabolism, and efflux in human monocytes and macrophages. Am J Physiol Cell Physiol 251:C128–C135
McKenna MJ, Morton J, Selig SE, Snow RJ (1999) Creatine supplementation increases muscle total creatine but not maximal intermittent exercise performance. J Appl Physiol 87:2244–2252
Ogawa Y (1994) Role of ryanodine receptors. Crit Rev Biochem Mol Biol 29:229–274
Preen D, Dawson B, Goodman C, Lawrence S, Beilby J, Ching S (2001) Effect of creatine loading on long-term sprint exercise performance and metabolism. Med Sci Sports Exerc 33:814–821
Pulido SM, Passaquin AC, Leijendekka WJ, Challet C, Wallimann T, Ruegg UT (1998) Creatine supplementation improves intracellular Ca2+ handling and survival in mdx skeletal muscle cells. FEBS Lett 439:357–362
Raguso CA, Coggan AR, Sidossis LS, Gastaldelli A, Wolfe RR (1996) Effect of theophylline on substrate metabolism during exercise. Metabolism 45:1153–1160
Schneiker KT, Bishop D, Dawson B, Hackett LP (2006) Effects of caffeine on prolonged intermittent-sprint ability in team-sport athletes. Med Sci Sports Exerc 38:578–585
Snow RJ, Murphy RM (2001) Creatine and the creatine transporter: a review. Mol Cell Biochem 224:169–181
Spriet LL, MacLean DA, Dyck DJ, Hultman E, Cederblad G, Graham TE (1992) Caffeine ingestion and muscle metabolism during prolonged exercise in humans. Am J Physiol Endocrinol Metab 262:E891–E898
Stuart GR, Hopkins WG, Cook C, Cairns SP (2005) Multiple effects of caffeine on simulated high-intensity team-sport performance. Med Sci Sports Exerc 37:1998–2005
Sutherland EW, Rall TW (1958) Fractionation and characterization of a cyclic adenine ribonucleotide formed by tissue particles. J Biol Chem 232:1077–1091
Van Soeren MH, Graham TE (1998) Effect of caffeine on metabolism, exercise endurance, and catecholamine responses after withdrawal. J Appl Physiol 85:1493–1501
Vanakoski J, Kosunen V, Meririnne E, Seppala T (1998) Creatine and caffeine in anaerobic and aerobic exercise: effects on physical performance and pharmacokinetic considerations. Int J Clin Pharmacol Ther 36:258–262
Vandenberghe K, Gillis N, Van Leemputte M, Van Hecke P, Vanstapel F, Hespel P (1996) Caffeine counteracts the ergogenic action of muscle creatine loading. J Appl Physiol 80:452–457
Williams JH (1991) Caffeine, neuromuscular function and high-intensity exercise performance. J Sports Med Phys Fit 31:481–489
Wright GA, Grandjean PW, Pascoe DD (2007) The effects of creatine loading on thermoregulation and intermittent sprint exercise performance in a hot humid environment. J Strength Cond Res 21:655–660
Yquel RJ, Arsac LM, Thiaudiere E, Canioni P, Manier G (2002) Effect of creatine supplementation on phosphocreatine resynthesis, inorganic phosphate accumulation and pH during intermittent maximal exercise. J Sports Sci 20:427–437
Ziegenfuss TN, Lowery LM, Lemon PWR (1998) Acute fluid volume changes in men during three days of creatine supplementation. J Exerc Physiol online 1(3). (http://faculty.css.edu/tboone2/asep/jan13d.htm)
Ziegenfuss TN, Rogers M, Lowery L, Mullins N, Mendel R, Antonio J, Lemon P (2002) Effect of creatine loading on anaerobic performance and skeletal muscle volume in NCAA Division I athletes. Nutrition 18:397–402
Acknowledgments
The authors thank all participants for their effort and dedication to the protocol. This study was supported by a research grant from National Science Council, Taiwan (NSC 97-2410-H-034-028).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Jean-René Lacour.
Rights and permissions
About this article
Cite this article
Lee, CL., Lin, JC. & Cheng, CF. Effect of caffeine ingestion after creatine supplementation on intermittent high-intensity sprint performance. Eur J Appl Physiol 111, 1669–1677 (2011). https://doi.org/10.1007/s00421-010-1792-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-010-1792-0