
Abstract
This study compared the parameter estimates of pulmonary oxygen uptake (VO2p), heart rate (HR) and muscle deoxygenation (Δ[HHb]) kinetics when several moderate-intensity exercise transitions (MODs) were performed during a single visit versus several MODs performed during separate visits. Nine subjects (24 ± 5 years, mean ± SD) each completed two successive cycling MODs on six occasions (1-6A and 1-6B) from 20 W to a work rate corresponding to 80% estimated lactate threshold with 6 min recovery at 20 W. During one visit, subjects completed two series of three MODs (6A-F), separated by 20 min rest. VO2p time constants (τVO2p; 27 ± 10 s, 25 ± 12 s, 25 ± 11 s) were similar (p > 0.05) for MODs 1-6A, 1-6B and 6A-F, respectively. τVO2p had reproducibility 95% confidence intervals (CI95) of 8.3, 8.2, 4.7, 4.9 and 4.7 s when comparing single (1A vs. 2A), the average of two (1-2A vs. 3-4A), three (1-3A vs. 4-6A), four (1-2AB vs. 3-4AB) and six (1-3AB vs. 4-6AB) MODs, respectively. The effective Δ[HHb] response time (τ′Δ[HHb]) was unaffected across conditions (1-6A: 19 ± 2 s, 1-6B: 19 ± 3 s, 6A-F: 17 ± 4 s) with reproducibility CI95 of 5.3, 4.5, 3.1, 2.9 and 3.3 s when a single, two, three, four and six MODs were compared, respectively. τHR was reduced in MODs 6A-F compared to 1-6A and 1-6B (23 ± 5 s, 25 ± 5 s, 27 ± 6 s, respectively). This study showed that parameter estimates of VO2p, HR and Δ[HHb] kinetics are largely unaffected by data collection sequence, and the day-to-day reproducibility of τVO2p and τ′Δ[HHb] estimates, as determined by the CI95, was appreciably improved by averaging of at least three MODs.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The first studies to specifically characterize the profile of pulmonary oxygen uptake (VO2p) after the onset of exercise were performed in the early 1970s (Linnarsson 1974; Whipp 1971; Whipp and Wasserman 1972); what emerged was a domain within exercise physiology known as VO2 kinetics. A description of the multi-phase exponential increase in VO2p (Whipp and Ward 1990; Whipp and Wasserman 1972) following a square-wave transition in exercise intensity was instrumental in the study of VO2p kinetics. However, perhaps of equal importance to the development of the field were the numerous paper published with the intent to describe and standardize data collection and analyses techniques (Bell et al. 2001b; Paterson and Whipp 1991; Rossiter et al. 1999; Whipp et al. 1982). As well, Lamarra et al. (1987) first demonstrated the importance of averaging data from multiple similar transitions (in order to maximize the signal-to-noise ratio) when modeling VO2p data. While the practice of averaging multiple transitions to generate a single, representative response for an individual has since become widely accepted (Bell et al. 2001a; Jones et al. 2003; Koga et al. 2005), the day-to-day reliability and reproducibility of the VO2p time constant (τVO2p) has not been established. Describing the inherent measurement error associated with τVO2p [by determining the reliability coefficients and reproducibility 95% confidence intervals (CI95)] may have important implications for studies in which an intervention is expected to change τVO2p; to date, only the CI95 describing the quality of the fit of a given mathematical model to a data set has been reported.
Data from previous studies (Burnley et al. 2000; Ferreira et al. 2005; Gerbino et al. 1996) designed to examine the physiological mechanisms underlying the regulatory features of oxidative phoshphorylation suggests that a single bout of moderate-intensity exercise (MOD) does not affect the VO2p profile, and specifically τVO2p of a subsequent MOD. However, these studies derived their VO2p kinetic parameters by fitting exponential models to data from only one (Gerbino et al. 1996) or the average of two (Burnley et al. 2000; Ferreira et al. 2005) similar transitions. In light of the recommendations that multiple transitions be averaged together when assessing VO2p kinetics (Lamarra et al. 1987), this study aimed to further investigate the effect of prior MODs on parameter estimates of VO2p kinetics, but with an improved ability to detect whether variations were present. Furthermore, it remains unknown whether there is a cumulative effect of performing several (e.g. three to six rather than two) sequential MODs within a single laboratory visit.
Many studies that consider VO2p kinetics also investigate changes in both the microvascular O2 extraction [estimated from near-infrared spectroscopy (NIRS)] and heart rate (HR) responses (Chin et al. 2007; DeLorey et al. 2004); at present, the impact of several (i.e., >2) repeated MODs on the parameter estimates of VO2p, NIRS-derived muscle deoxygenation (Δ[HHb]) and HR kinetics of subsequent MODs has yet to be systematically addressed. Additionally, information about intra-individual reproducibility of the effective response time of the Δ[HHb] signal (τ′Δ[HHb]; i.e., overall time course of adjustment) is lacking. Therefore the purpose of this study was: (1) to determine in a group of healthy, young subjects whether the parameter estimates of VO2p, Δ[HHb] and HR kinetics differ when the average of several MODs performed during a single visit are compared to the average of several single MODs performed during separate visits, and (2) to determine the intra-individual reproducibility CI95 of τVO2p and τ′Δ[HHb] estimates. It was hypothesized that the parameter estimates of VO2p kinetics would not be altered by serial MODs in 1 day. Despite potential effects of prior exercise on the baseline cardiovascular and muscle metabolic responses, it was hypothesized that the profiles of both HR and ∆[HHb] would be unaffected by serial repetitions. Finally, it was hypothesized that intra-individual (day-to-day) measures of τVO2p and τ′Δ[HHb] would be reliable (Pearson’s correlation coefficient) and reproducible (as evidenced by narrow reproducibility CI95 values).
Methods
Subjects
Nine young adults (4 women and 5 men, 24 ± 5 year; 70 ± 7 kg; 179 ± 6 cm; mean ± SD) volunteered and gave written consent to participate in the study. All procedures were approved by The University of Western Ontario Research Ethics Board for Health Sciences Research Involving Human Subjects. All subjects were non-obese (body mass index ≤ 30 kg/m2), non-smokers, and were physically active. Additionally, no subjects were taking medications that would affect the cardiorespiratory or hemodynamic responses to exercise.
Protocol
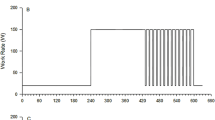
On day 1, subjects reported to the laboratory to perform a fatigue-limited ramp incremental test (20–25 W/min) on a cycle ergometer (model: H-300-R Lode; Lode B.V., Groningen, Holland) for determination of peak VO2 (VO2peak) and the estimated lactate threshold (θ L). θ L was defined as the VO2 at which CO2 output (VCO2) began to increase out of proportion to VO2 with a systematic rise in minute ventilation-to-VO2 ratio and end-tidal PO2 whereas minute ventilation-to-VCO2 ratio and end-tidal PCO2 were stable. Subsequent to the incremental test, subjects returned to the laboratory on six occasions. During each visit, subjects completed two successive MODs from a 6 min baseline work rate (WR) of 20 W to a 6 min WR corresponding to 80% θ L (termed MODs 1-6A and 1-6B; Fig. 1). During one randomly assigned visit, subjects continued on to complete two series of three successive MODs, with each series separated by 20 min of seated recovery (termed MODs 6A-F; Fig. 1). Each visit to the laboratory was separated by at least 24 h.

Schematic of moderate intensity exercise step-transitions from 20 W baseline to a WR corresponding to 80% θ L showing the first of six transitions performed on separate days (labelled 1-6A), the second of six transitions performed on separate days (labelled 1-6B), and serial transitions performed all in one day (labelled 6A-F). The order of the visits was randomized
Measurements
Gas-exchange measurements were similar to those previously described (Babcock et al. 1994). Briefly, inspired and expired flow rates were measured using a low dead space (90 mL) bidirectional turbine (Alpha Technologies VMM 110) which was calibrated before each test using a syringe of known volume. Inspired and expired gases were continuously sampled (50 Hz) at the mouth and analyzed for concentrations of O2, CO2, and N2 by mass spectrometry (Innovision, AMIS 2000, Lindvedvej, Denmark) after calibration with precision-analyzed gas mixtures. Changes in gas concentrations were aligned with gas volumes by measuring the time delay for a square-wave bolus of gas passing the turbine to the resulting changes in fractional gas concentrations as measured by the mass spectrometer. Data were transferred to a computer, which aligned concentrations with volume information to build a profile of each breath. Breath-by-breath alveolar gas exchange was calculated by using algorithms of Beaver et al. (1981).
HR was monitored continuously by electrocardiogram (three-lead arrangement) using PowerLab (ML132/ML880; ADInstruments, Colorado Springs, CO, USA). Data were recorded using LabChart v4.2 (ADInstruments, Colorado Springs, CO, USA) on a separate computer.
Local muscle deoxygenation profiles of the quadriceps vastus lateralis muscle were made with NIRS (Hamamatsu NIRO 300, Hamamatsu Photonics, Hamamatsu, Japan). Optodes were placed on the belly of the muscle midway between the lateral epicondyle and greater trochanter of the femur. The optodes were housed in an optically-dense plastic holder, secured on the skin surface with tape, and covered with an optically-dense, black vinyl sheet, thus minimizing the intrusion of extraneous light. The thigh was wrapped with an elastic bandage to minimize movement of the optodes.
The physical principles of tissue spectroscopy are described in detail by Elwell (1995) and the manner in which these are applied have been explained by DeLorey et al. (2003). Briefly, one fiber optic bundle carried the NIR-light produced by the laser diodes to the tissue of interest while a second fiber optic bundle (interoptode spacing = 5 cm) returned the transmitted light from the tissue to a photon detector (photomultiplier tube) in the spectrometer. Four laser diodes (λ = 775, 810, 850, and 910 nm) were pulsed in a rapid succession and the light was detected by the photomultiplier tube for online estimation and display of the concentration changes from the resting baseline for oxyhaemoglobin, deoxyhaemoglobin (Δ[HHb]), and total haemoglobin. Changes in light intensities were recorded continuously at 2 Hz and transferred to a computer for later analysis. The NIRS-derived signal was zero set with the subject sitting at rest on the cycle ergometer prior to the onset of baseline exercise (MOD A and MOD D; see Fig. 1) and changes in the concentration are reported as a delta (∆) in arbitrary units (a.u.).
Data analysis
VO2p data were filtered by removing aberrant data points that lay outside 4 SD of the local mean; the justification for this filtering process was provided by Lamarra et al. (1987), who demonstrated that ″noise″ observed within the VO2p signal conformed to a predictable Gaussian distribution, independent of work rate. The data for each transition were linearly-interpolated to 1 s intervals and time-aligned such that time zero represented the onset of exercise. Data from transitions across days (1-6A; 1-6B) and within a day (6A-F) were ensemble-averaged to yield three averaged responses for each subject. These transitions were further time-averaged into 5 s bins to provide three time-averaged responses for each subject. Baseline VO2p (VO2pbsln) was calculated as the average VO2p collected between 40 and 10 s before an increase in WR; this 30 s baseline was selected in order to minimize any possible effects of slow off-transient kinetics. The on-transient responses for VO2p were modeled using the following equation:
where Y(t) represents the VO2p for any given time; YB is the VO2p at baseline; A is the amplitude of the VO2p response; t is a given amount of time; τ represents the time required to attain 63% of the steady-state amplitude; and TD represents the time delay.
After excluding the initial 20 s of data (i.e., cardiodynamic phase) from the model, while still allowing TD to vary freely (in order to optimize accuracy of parameter estimates), VO2p data were modeled from the beginning of phase II to 4 min (240 s) of the step-transition; this ensured that each subject had attained a VO2p steady-state, yet did not bias the model fit during the on-transient (Bell et al. 2001b). The model parameters were estimated by least-squares nonlinear regression (Origin, OriginLab Corp., Northampton, MA, USA) in which the best fit was defined by minimization of the residual sum of squares and minimal variation of residuals around the Y axis (Y = 0). The 95% confidence interval of the model fit (CI95-model) for the estimated time constant was determined after preliminary fit of the data with Y B, A, and TD constrained to the best-fit values and the τ allowed to vary.
HR data were determined from the R–R interval on a second-by-second basis and edited and modeled in the same manner as the VO2p data described above. The on-transient HR response was modeled from the onset of exercise to 240 s using the exponential model described in Eq. 1.
Three NIRS-derived ∆[HHb] responses (1-6A, 1-6B and 6A-F) were generated for each subject. The Δ[HHb] profile has been described to consist of a time delay at the onset of exercise, followed by an increase in the signal with an “exponential-like” time-course. The time delay for the ∆[HHb] response (TD Δ[HHb]) was determined using second-by-second data and corresponded to the time, after the onset of exercise, at which the ∆[HHb] signal began a systematic increase from its nadir value. Determination of the TD ∆[HHb] was made on individual trials and averaged to yield a value for each individual in each of the three “conditions.” The ∆[HHb] data were modeled from the end of the TD ∆[HHb] to 90 s of the transition using an exponential model as described in Eq.1. As previously described by duManoir et al. (2010), different fitting strategies (i.e., 90–180 s) resulted in minimal differences (<2 s) in estimates of τ∆[HHb] and the early exponential increase in ∆[HHb] was well-characterized in the 90 s following exercise onset. The τ∆[HHb] described the time course for the increase in ∆[HHb], while the overall change of the effective ∆[HHb] (τ′∆[HHb] = TD ∆[HHb] + τ∆[HHb]) described the overall time course of the ∆[HHb] from the onset of exercise.
Statistics
Data are presented as mean ± SD. Repeated measures analysis of variance (ANOVA) was used to determine statistical significance for the dependent variables. A Tukey post-hoc analysis was used when significant differences were found for the main effects of each dependent variable. Pearson’s product-moment (reliability) correlation coefficients and intra-individual CI95 were computed for τVO2p and τ′∆[HHb] estimates. All statistical analyses were performed using SPSS Version 16.0, (SPSS Inc., Chicago, IL, USA). Statistical significance was declared when p < 0.05.
Results
Mean VO2peak and peak power output during the ramp incremental tests were 3.5 ± 0.8 L min−1 and 303 ± 61 W, respectively. The mean power output during MODs was 119 ± 34 W. Table 1 shows the parameters of VO2p kinetics. None of τVO2p, the amplitude of the VO2p response (VO2pamp), VO2 gain (ΔVO2p/ΔWR), the CI95-model τVO2p, nor the VO2p time delay (TD VO2p) were different when comparing transitions 1-6A, 1-6B and 6A-F. Figure 2 presents the dispersion of individual data around the mean, along with Bland–Altman plots and further corroborates the similarities in τVO2p between transitions 1-6A and 1-6B (r = 0.97, p < 0.05), 1-6A and 6A-F (r = 0.91, p < 0.05), and 1-6B and 6A-F (r = 0.90, p < 0.05), respectively. The VO2pbsln was significantly elevated in MODs 1-6B and 6A-F compared to 1-6A, and steady state VO2p (VO2pss) was significantly higher in MODs 6A-F compared to 1-6A or 1-6B.

Comparison of individual (open circle) and mean (closed circle) τVO2p values and associated Bland–Altman plots from MODs 1-6B vs. 1-6A (r = 0.97; a, d); 6A-F vs. 1-6A (r = 0.91; b, e); and 6A-F vs. 1-6B (r = 0.90; c, f). The line of identity (a–c) and limits of agreement (d–f) are represented by the dotted lines
The day-to-day reliability and reproducibility of VO2p kinetics parameter estimates were also investigated by comparing the responses from a single transition (MODs 1A vs. 2A), the average of two (1-2A vs. 3-4A), three (1-3A vs. 4-6A), four (1-2AB vs. 3-4AB) and six MODs (1-3AB vs. 4-6AB), respectively. No significant differences were observed for any of these comparisons for VO2pbsln, VO2pamp, VO2pss, τVO2p, CI95-model τVO2p and TD VO2p (Table 2). Reliability coefficients ranged from r = 0.87–0.89, while the associated CI95 were notably reduced in comparisons of three or more MODs rather than a single or the average of two MODs (MOD 1A vs. 2A: r = 0.87, CI95 = 8.3 s; MODs 1-2A vs. 3-4A: r = 0.88, CI95 = 8.2 s; MODs 1-3A vs. 4-6A: r = 0.87, CI95 = 4.7; MODs 1-2AB vs. 3-4AB: r = 0.89, CI95 = 4.9 s; MODs 1-3AB vs. 4-6AB: r = 0.87, CI95 = 4.7 s). These data suggest that at least three MODs are required in order to reduce the influence of normal day-to-day variability in estimates of τVO2p. Figure 3 depicts the reliability and reproducibility and the dispersion of individual τVO2p data around the mean for each of the five comparison groups described above.

Comparison of individual (open circle) and mean (closed circle) τVO2p values from a MOD 2A vs. 1A (r = 0.87, CI95 = 8.3 s); b MODs 3-4A vs. 1-2A (r = 0.88, CI95 = 8.2 s); c) MODs 4-6A vs. 1-3A (r = 0.87, CI95 = 4.7); d MODs 3-4AB vs. 1-2AB (r = 0.89, CI95 = 4.9 s); and e MODs 4-6AB vs. 1-3AB (r = 0.87, CI95 = 4.7 s). Error bars are SD. The line of identity is represented by the dotted line
The mean τHR for MODs 6A-F was significantly lower than for MODs 1-6A and 1-6B (Table 3). Baseline HR (HRbsln) was higher in MODs 1-6B (p < 0.05) but not MODs 6A-F (p = 0.066) when compared with MODs 1-6A; however, the amplitude of the HR response (HRamp) for both MODs 1-6B and MODs 6A-F were significantly less than that of MODs 1-6A. Thus, the steady state HR (HRss) was similar amongst all three responses. No differences were identified for either the CI95-model τHR or the HR time delay (TD HR) when comparing MODs 1-6A, 1-6B and 6A-F.
There were no differences in τΔ[HHb], Δ[HHb] amplitude (Δ[HHb]amp), TD Δ[HHb], or τ′Δ[HHb] amongst MODs 1-6A, 1-6B and 6A-F (Table 4). Baseline Δ[HHb] (Δ[HHb]bsln) for MODs 6A-F was significantly greater than MODs 1-6B, and both the steady state Δ[HHb] (Δ[HHb]ss) and CI95-model τΔ[HHb] were significantly different when comparing MODs 1-6A and 6A-F. Like τVO2p, reproducibility CI95 for τ′Δ[HHb] (n = 8) were reduced when comparing three or more MODs with a single or the average of two MODs (MOD 1A vs. 2A CI95 = 5.3 s; MODs 1-2A vs. 3-4A CI95 = 4.5 s; MODs 1-3A vs. 4-6A CI95 = 3.1; MODs 1-2AB vs. 3-4AB CI95 = 2.9 s; MODs 1-3AB vs. 4-6AB CI95 = 3.3 s). Figure 4 shows the dispersion of individual τ′Δ[HHb] data around the mean for each of these groups.

Comparison of individual (open circle) and mean (closed circle) τ′Δ[HHb] values from a MOD 2A vs. 1A (CI95 = 5.3 s); b MODs 3-4A vs. 1-2A (CI95 = 4.5 s); c MODs 4-6A vs. 1-3A (CI95 = 3.1 s); d MODs 3-4AB vs. 1-2AB (CI95 = 2.9 s); and e MODs 4-6AB vs. 1-3AB (CI95 = 3.3 s). Error bars are SD. The line of identity is represented by the dotted line
Discussion
The primary purpose of this study was to determine whether the parameter estimates of VO2p, HR and Δ[HHb] kinetics differ when the average of several MODs performed during a single visit are compared to the average of single MODs performed during separate visits. The major findings are: (1) despite small but significant differences in VO2pbsln and VO2pss, neither τVO2p nor the VO2p gain were affected by repeated transitions within a single visit to the laboratory; (2) several of the HR kinetics parameter values were altered after performing either two (i.e., 1-6B; elevated HRbsln and reduced HRamp) or six (reduced τHR and HRamp) MODs in a single laboratory visit relative to when a single MOD was performed on separate visits; (3) few parameter measures of Δ[HHb] kinetics were affected by serial testing within a single laboratory visit. This study also established the day-to-day intra-individual reproducibility of τVO2p and τ′Δ[HHb] estimates; importantly, the reproducibility CI95 were noticeably reduced such that when three or more MODs were used, the intra-individual CI95 for τVO2p was <5 s and for τ′Δ[HHb] was ~3 s.
It has been suggested that a prior bout of moderate-intensity exercise does not affect the parameter estimates of VO2p kinetics (mean response time) during a subsequent MOD (Gerbino et al. 1996). Burnley et al. (2000) and Ferreira et al. (2005) also showed no effect of prior moderate exercise on τVO2p, VO2pbsln and VO2pamp during a subsequent MOD performed after 6 min of ‘recovery cycling’ at 20 W. Nevertheless, these studies used a single (Gerbino et al. 1996) or the average of two MODs (Burnley et al. 2000; Ferreira et al. 2005) to determine their VO2p kinetics parameters rather than multiple transitions, which have been shown to improve the signal-to-noise ratio and confidence in parameter estimates. This is the first study to use repeated transitions (either on different days or with serial tests on a single day) to show that estimates of τVO2p are unaffected by performance of serial MODs.
When the experimental design is facilitated by having multiple MODs performed in a single visit, estimates of τVO2p are valid; however, consideration must be given to the altered baseline and steady state metabolic rates. It was somewhat surprising that VO2pss was greater when six transitions were performed in sequence, rather than when only one or two MODs were completed within a single visit. Interestingly, this appears to be a reflection of differences in VO2pbsln, as the VO2 gain (and VO2pamp) was unaffected by the data collection sequence; that is, VO2p gain is independent of small, but significant variations in VO2pbsln (i.e., during 20 W exercise) when work is performed below θ L. That subjects may have an altered VO2pbsln even when MODs are performed on separate days may be inconsequential as this appears not to affect the other parameters of VO2p kinetics.
The significantly greater VO2pbsln seen in MODs 1-6B (by 0.04 L min−1) and 6A-F (by 0.06 L min−1) compared to MODs 1-6A differs from the unchanged VO2p kinetics parameters reported previously (Burnley et al. 2000; Ferreira et al. 2005). Since a more rapid on-transient response may suggest a faster off-transient response within the moderate intensity domain (Paterson and Whipp 1991) and return towards VO2pbsln, this difference between studies may be a by-product of the very small τVO2p (16 s) reported by Burnley et al. (2000) compared to the larger τVO2p response (27 s) in the present study. This is unlikely, however, as Ferreira et al. (2005) observed unchanged VO2pbsln with repeated MODs in individuals with ~23 s τVO2p kinetics.
Regardless of the number of MODs averaged together, the parameter estimates of VO2p kinetics were shown to be reproducible (Table 2). Lamarra et al. (1987) noted that having subjects perform multiple MODs improves the confidence in τVO2p estimates; the data presented in Table 4 substantiate this analysis. A progressive decrease in both CI95-model τVO2p and the SD associated with each mean τVO2p estimate is observed as the number of MODs increases (i.e., one, two, three, four and six); however, the quality (confidence) of the modeling with averaging of additional MODs only reflects the reduction in signal noise (greater signal-to-noise ratio), not the intra-individual reliability of the measure itself. Determining the day-to-day reliability and reproducibility of τVO2p estimates generated from few (one or two) or many (three to six) MODs has potentially important implications in the interpretation of study results. For instance, a reduction in τVO2p of 6 s following a training intervention may be observed, yet if this parameter were estimated based upon the averaged response from two MODs for each individual, the change would be within the CI95 of day-to-day reproducibility. Although the reliability coefficients were virtually unaffected by the number of MODs (ranging from r = 0.87–0.89), the reproducibility CI95 was appreciably reduced (from >8 s to <5 s) when three or more MODs were averaged together compared to only one or two. It must be noted that the reproducibility CI95 for a given number of MODs is a finite value with no variance, and as such no statistical inferences can be drawn. Nonetheless, these finding suggests that at least three MODs are required in order to reduce the influence of normal day-to-day variability in estimates of τVO2p. That there was no further reduction in the reproducibility CI95 when four or six MODs were compared to three suggests that performing these additional transitions may have been unnecessary, particularly in light of the relatively small changes in CI95-model τVO2p (confidence of the model fit). It is important to consider the influence of subject fitness on these conclusions, however. The mean power output for subjects tested in the present study was 119 ± 34 W, leading to a mean VO2pamp of approximately 0.97 L min−1; in populations where the signal amplitude would be expected to be lower than that observed in the present study, more repeats could likely be required as demonstrated by Lamarra et al (1987). Additionally, recognizing that the VO2p data in the present study were collected using a mass spectrometer and volume turbine (steady state measurement error: SD = 0.12 L min−1) is of importance when considering this conclusion, as this method of data acquisition (“alveolar” gas exchange with measured inspiratory and expiratory volumes and nitrogen concentrations ([N2]) in order to estimate changes in lung gas stores dynamically) reduces breath-by-breath “noise” (greater signal-to-noise ratio) when compared to some metabolic carts, for example.
Additionally, it was of interest to note that compared to the day-to-day variability of τVO2p assessed using six MODs (1-3AB vs. 4-6AB; r = 0.87), τVO2p estimates generated using the serial testing in a single day (MODs 6A-F) actually displayed slightly less variability in comparison to six MODs conducted one per day (MODs 1-6A, r = 0.91; Fig. 2). This finding supports the suggestion that estimates of τVO2p that were generated by averaging the VO2p profile from several serial transitions (i.e. 6A-F) are valid.
As observed with τVO2p, the CI95 values determined for τ′Δ[HHb] also suggest that averaging responses from at least three MODs is required in order to improve day-to-day reproducibility, with no further improvement observed with the inclusion of additional MODs (i.e., four and six, respectively). The day-to-day CI95 was ~3 s compared to ~5 s for τVO2p when averaging the responses from at least three MODs. Interestingly, meaningful reliability coefficients could not be calculated for this variable as they were for τVO2p. The subjects in the present study presented with a broad range of τVO2p values (SD ~11 s), thus allowing for easy discrimination amongst individuals when calculating a correlation coefficient. The τ′Δ[HHb] values observed in the present study, however, were much more closely clustered (SD ~3 s) and as such, a small change in the parameter estimate from day-to-day (e.g., 3 s) would necessarily imply a dramatic weakening in the observed correlation coefficient. The tightly clustered τ′Δ[HHb] values are not unique to this study, however, as similar SD values have previously been reported in young subjects (Murias et al. 2010).
To date, the effects of performing several prior MODs on the kinetics of HR or local muscle O2 extraction during subsequent moderate exercise have not been systematically studied; Ferreira et al. (2005) considered the effect of a single MOD on a subsequent MOD on the Δ[HHb] response. Given the potential effects of prior exercise on the cardiovascular (e.g., vasodilation, elevated HRbsln) and muscle metabolic (e.g., enzyme activity) responses, consideration of the effects of performing multiple MODs on HR and Δ[HHb] kinetics is warranted. Indeed, several parameters of HR kinetics were affected by repeated MODs within a single laboratory visit. Like VO2p, HRbsln was greater in MODs 1-6B and 6A-F than in MODs 1-6A. In demonstrating the asymmetry of the on- and off-transient cardiac output (Q) responses during moderate exercise, Yoshida and Whipp (1994) pointed out that the slower post-exercise response acts to maintain sufficient O2 supply to muscles during a phase where metabolic demand remains high. Thus, the fact that HR remained elevated 6 min after the cessation of a bout of moderate exercise (i.e. HRbsln preceding MODs 1-6B and 6A-F) may simply be a reflection of the relatively higher VO2pbsln.
Notably, τHR was significantly larger in MODs 1-6A and 1-6B than in MODs 6A-F. This finding is unexpected since increases in HRbsln may increase the relative contribution of (slower) sympathetic activation, rather than the more rapid withdrawal of parasympathetic activity that together govern the adjustment of HR. Regardless, the faster HR adjustment in the absence of changes to τVO2p in MODs 6A-F implies that bulk delivery of O2 is not a limiting factor in the phase II τVO2p response to moderate intensity exercise. Although not seen in the present study, other studies have consistently reported faster HR kinetics compared to VO2p kinetics, and as Poole et al. (2008) assert, it is difficult to envision a faster process limiting a slower one. The altered HR response nevertheless highlights cardiovascular adjustments in response to prior moderate intensity exercise that deserve consideration.
The parameters of Δ[HHb] kinetics appeared to be largely unaffected when comparing the average response of MODs performed on a single visit versus the average of MODs performed during separate visits. Importantly, none of τΔ[HHb], Δ[HHb]amp, TD Δ[HHb], nor τ′Δ[HHb] were affected by serial testing within a single visit. These findings are in opposition to those reported by Ferreira et al. (2005), who observed a decreased TD Δ[HHb] and increased τΔ[HHb] (with no change in τ′Δ[HHb]) in ‘MODs 1-2B’ compared to ‘MOD 1-2A’ (as they would be identified in the present paper). However, it must be noted that the TD Δ[HHb] reported by Ferreira et al. (2005) was derived in a different fashion than that reported in the present study. Whereas our method has been to plot second-by-second data for each individual transition to identify the time, after the onset of exercise, at which the ∆[HHb] signal begins a systematic increase from its nadir value (indicating a physiological change within the tissue that is specific to an individual transition), Ferreira et al. (2005) reported the TD Δ[HHb] derived from a mathematical model of the ensemble-averaged data where the mono-exponential component is projected to intersect with the linear (i.e., baseline) component. As their Fig. 1 shows, this modeled TD Δ[HHb] can be influenced by the time window selected for inclusion in the model as well as the dispersion of the data; our approach has no such bias. Furthermore, a change in the modeled TD Δ[HHb] would be expected to impact the estimation of τΔ[HHb] (such that an increase in one parameter is often accompanied by a decrease in the other), which makes accurate identification of the TD Δ[HHb] of critical importance in the calculation of the Δ[HHb] parameter estimates.
Although the Δ[HHb]bsln and Δ[HHb]ss (the latter exclusively related to differences in the former in the absence of changes in Δ[HHb]amp) estimates for MODs 6A-F differed from 1-6B and 1-6A respectively, this is not necessarily indicative of an altered physiological response; that is, the elevated Δ[HHb]bsln in MODs 6A-F should not be interpreted as an increased reliance on O2 extraction. Rather, because the measurement units are arbitrary, small differences in probe placement amongst the six separate visits might explain the altered baseline parameter values. That the baseline and steady state values in MODs 1-6A and 1-6B were virtually identical is consistent with this interpretation. Interestingly, MODs 6A-F, in which probe placement was unchanged, produced a smaller CI95-model τΔ[HHb] than MODs 1-6A. These data suggest that performing serial MODs within a single visit to the laboratory will not significantly affect the parameter estimates of Δ[HHb] kinetics.
In conclusion, this study has shown that neither τVO2p nor the VO2p gain were affected by repeated transitions within a single visit to the laboratory, in spite of small but significant differences in VO2pbsln and VO2pss. Both τVO2p and τ′Δ[HHb] were shown to be stable variables that can be reliably measured from day-to-day, but with a narrowed reproducibility CI95 when three or more MODs are used in their estimation. While few parameters of Δ[HHb] kinetics were altered by the sequence of data acquisition, some parameter estimates for HR kinetics were affected. While the potential effects of prior exercise on the cardiovascular (e.g., vasodilation) and muscle metabolic (e.g., enzyme activity) responses cannot be ignored, data from the present study suggest that performing multiple, serial MODs does not alter the response to the MOD exercise transition and does not adversely influence the estimation of VO2p kinetics parameters.
References
Babcock MA, Paterson DH, Cunningham DA, Dickinson JR (1994) Exercise on-transient gas exchange kinetics are slowed as a function of age. Med Sci Sports Exerc 26:440–446
Beaver WL, Lamarra N, Wasserman K (1981) Breath-by-breath measurement of true alveolar gas exchange. J Appl Physiol 51:1662–1675
Bell C, Paterson DH, Kowalchuk JM, Moy AP, Thorp DB, Noble EG, Taylor AW, Cunningham DA (2001a) Determinants of oxygen uptake kinetics in older humans following single-limb endurance exercise training. Exp Physiol 86:659–665
Bell C, Paterson DH, Kowalchuk JM, Padilla J, Cunningham DA (2001b) A comparison of modelling techniques used to characterise oxygen uptake kinetics during the on-transient of exercise. Exp Physiol 86:667–676
Burnley M, Jones AM, Carter H, Doust JH (2000) Effects of prior heavy exercise on phase II pulmonary oxygen uptake kinetics during heavy exercise. J Appl Physiol 89:1387–1396
Chin LM, Leigh RJ, Heigenhauser GJ, Rossiter HB, Paterson DH, Kowalchuk JM (2007) Hyperventilation-induced hypocapnic alkalosis slows the adaptation of pulmonary O2 uptake during the transition to moderate-intensity exercise. J Physiol 583:351–364
DeLorey DS, Kowalchuk JM, Paterson DH (2003) Relationship between pulmonary O2 uptake kinetics and muscle deoxygenation during moderate-intensity exercise. J Appl Physiol 95:113–120
DeLorey DS, Kowalchuk JM, Paterson DH (2004) Effect of age on O2 uptake kinetics and the adaptation of muscle deoxygenation at the onset of moderate-intensity cycling exercise. J Appl Physiol 97:165–172
duManoir GR, DeLorey DS, Kowalchuk JM, Paterson DH (2010) Kinetics of VO2 limb blood flow and regional muscle deoxygenation in young adults during moderate intensity, knee-extension exercise. Eur J Appl Physiol 108:607–617
Elwell C (1995) A practical users guide to near infrared spectroscopy. Hamamatsu Photonics KK, London
Ferreira LF, Lutjemeier BJ, Townsend DK, Barstow TJ (2005) Dynamics of skeletal muscle oxygenation during sequential bouts of moderate exercise. Exp Physiol 90:393–401
Gerbino A, Ward SA, Whipp BJ (1996) Effects of prior exercise on pulmonary gas-exchange kinetics during high-intensity exercise in humans. J Appl Physiol 80:99–107
Jones AM, Wilkerson DP, Koppo K, Wilmshurst S, Campbell IT (2003) Inhibition of nitric oxide synthase by L-NAME speeds phase II pulmonary VO2 kinetics in the transition to moderate-intensity exercise in man. J Physiol 552:265–272
Koga S, Poole DC, Shiojiri T, Kondo N, Fukuba Y, Miura A, Barstow TJ (2005) Comparison of oxygen uptake kinetics during knee extension and cycle exercise. Am J Physiol Regul Integr Comp Physiol 288:R212–R220
Lamarra N, Whipp BJ, Ward SA, Wasserman K (1987) Effect of interbreath fluctuations on characterizing exercise gas exchange kinetics. J Appl Physiol 62:2003–2012
Linnarsson D (1974) Dynamics of pulmonary gas exchange and heart rate changes at start and end of exercise. Acta Physiol Scand Suppl 415:1–68
Murias JM, Kowalchuk JM, Paterson DH (2010) Speeding of VO2 kinetics with endurance training in old and young men is associated with improved matching of local O2 delivery to muscle O2 utilization. J Appl Physiol 108:913–922
Paterson DH, Whipp BJ (1991) Asymmetries of oxygen uptake transients at the on- and offset of heavy exercise in humans. J Physiol 443:575–586
Poole DC, Barstow TJ, McDonough P, Jones AM (2008) Control of oxygen uptake during exercise. Med Sci Sports Exerc 40:462–474
Rossiter HB, Ward SA, Doyle VL, Howe FA, Griffiths JR, Whipp BJ (1999) Inferences from pulmonary O2 uptake with respect to intramuscular [phosphocreatine] kinetics during moderate exercise in humans. J Physiol 518:921–932
Whipp BJ (1971) Rate constant for the kinetics of oxygen uptake during light exercise. J Appl Physiol 30:261–263
Whipp BJ, Ward SA (1990) Physiological determinants of pulmonary gas exchange kinetics during exercise. Med Sci Sports Exerc 22:62–71
Whipp BJ, Wasserman K (1972) Oxygen uptake kinetics for various intensities of constant-load work. J Appl Physiol 33:351–356
Whipp BJ, Ward SA, Lamarra N, Davis JA, Wasserman K (1982) Parameters of ventilatory and gas exchange dynamics during exercise. J Appl Physiol 52:1506–1513
Yoshida T, Whipp BJ (1994) Dynamic asymmetries of cardiac output transients in response to muscular exercise in man. J Physiol 480:355–359
Acknowledgments
We would like to express our gratitude to the subjects in this study and to acknowledge the assistance provided by Brad Hansen. This study was supported by Natural Sciences and Engineering Research Council of Canada (NSERC) research and equipment grants. Additional support was provided by Standard Life Assurance Company of Canada. J. M. Murias was supported by a doctoral research scholarship from the Canadian Institutes of Health Research (CIHR).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Susan Ward.
Rights and permissions
About this article
Cite this article
Spencer, M.D., Murias, J.M., Lamb, H.P. et al. Are the parameters of VO2, heart rate and muscle deoxygenation kinetics affected by serial moderate-intensity exercise transitions in a single day?. Eur J Appl Physiol 111, 591–600 (2011). https://doi.org/10.1007/s00421-010-1653-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-010-1653-x