
Abstract
Oxidative stress occurs at altitude, and physical exertion might enhance this stress. In the present study, we investigated the combined effects of exercise and moderate altitude on redox balance in ten endurance exercising biathletes, and five sedentary volunteers during a 6-week-stay at 2,800 m. As a marker for oxidative stress, hydrogen peroxide (H2O2) was analyzed by the biosensor measuring system Ecocheck™, and 8-iso prostaglandin F2α (8-iso PGF2α) was determined by enzyme immunoassay in exhaled breath condensate (EBC). To determine the whole blood antioxidative capacity, we measured reduced glutathione (GSH) enzymatically using Ellman’s reagent. Exercising athletes and sedentary volunteers showed increased levels of oxidative markers at moderate altitude, contrary to our expectations; there was no difference between both groups. Therefore, all subjects’ data were pooled to examine the oxidative stress response exclusively due to altitude exposure. H2O2 levels increased at altitude and remained elevated for 3 days after returning to sea level (p ≤ 0.05). On the other hand, 8-iso PGF2α levels showed a tendency to increase at altitude, but declined immediately after returning to sea level (p ≤ 0.001). Hypoxic exposure during the first day at altitude resulted in elevated GSH levels (p ≤ 0.05), that decreased during prolonged sojourn at altitude (p ≤ 0.001). In conclusion, a stay at moderate altitude for up to 6 weeks increases markers of oxidative stress in EBC independent of additional endurance training. Notably, this oxidative stress is still detectable 3 days upon return to sea level.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Altitude training is frequently used by endurance athletes to improve competition performance at sea level (Levine and Stray-Gundersen 1997). Three to four weeks at moderate altitude is the minimal recommended period required to maximize the erythropoietic response (Chapman and Levine 2007; Heinicke et al. 2005). However, not all aspects of altitude acclimatization may be beneficial for competing athletes. In particular, it is known that exposure to altitude combined with exercise (Araneda et al. 2005; Bailey et al. 2001; Chao et al. 1999; Pfeiffer et al. 1999; Schmidt et al. 2002), and even exposure to altitude in resting conditions (Bailey et al. 2001), increases reactive oxygen species (ROS) levels/formation, especially in lung and blood. On the other hand, acute physical work per se is a contributor to oxidative stress where mitochondria represent the major source of free radicals (Bailey et al. 2001; Bailey et al. 2004; Sen 1995). Apart from exercise, additional stressors increase oxidative stress at altitude, such as exposure to ultra violet light, lack of dietary antioxidants, cold climate, increased xanthine oxidase activation and catecholamine production, as well as increased reductive stress and generation of free radicals through anoxia/reoxygenation (Askew 2002). Excessive generation of ROS can induce both, a detrimental effect on lipids, proteins, and DNA as well as an adaptive response. Several studies demonstrate the pathophysiological significance of oxidative stress molecules such as hydrogen peroxide (H2O2) and 8-iso prostaglandin F2α (8-iso PGF2α) in relation to cellular inflammation and lung diseases (Makris et al. 2007; Montuschi 2005; Psathakis et al. 2006). H2O2 is produced after converting superoxide anions O ·−2 to H2O2 by superoxide dismutase and is less stable than 8-iso PGF2α that is a product of arachidonic acid oxidation by ROS. Collection of exhaled breath condensate (EBC) offers the opportunity to measure these markers in the lower respiratory tract in a noninvasive manner (Horvath et al. 2005). In addition, maintenance of redox state depends on the efficiency of antioxidant defense systems and the amount of ROS generated. As such, reduced glutathione (GSH) is one of the most important cellular antioxidants that plays a central role in the regulation of redox sensitive signal transduction (Sen 1999).
Despite its impact, there is little knowledge as to whether oxidative stress increases during a prolonged period of exercise at moderate altitude. We hypothesized that a cumulated effect of exercise and hypoxia exists that causes up-regulation of free radical production, and consequently, an increase in oxidative stress markers. Therefore, using EBC and blood we determined oxidative stress response parameters upon exercise or rest during a 6-week-sojourn at moderate altitude (2,800 m).
Methods
Subjects and protocol
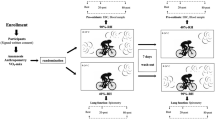
Ten healthy, endurance trained biathletes, and five sedentary, control subjects, all 15 volunteers being members of the Chilean Military “Escuela de Montaña”, who usually live at sea level, were investigated during a 6-week-training camp or stay at moderate altitude (2,800 m) in Portillo, Chile. The subjects gave their written informed consent to participate in this study, which was approved by the Ethical Committee for Humans from the Faculty of Medicine, University of Chile, Santiago and supported by the headquarter of the Chilean Military. The daily training time of the athletes [four females and six males; mean age (SD), 24.7 ± 1.3 years, 167.2 ± 9.5 cm, 63.1 ± 8.0 kg, BMI 22.5 ± 1.5 kg/m2] was 4–6 h with one rest day per week. Training included extensive cross country skiing, strength training, and shooting technique training. The sedentary control subjects (five males, 30.2 ± 3.3 years, 168.2 ± 5.3 cm, 79.8 ± 7.1 kg, BMI 28.2 ± 1.7 kg/m2) performed mainly office work and regular daily life tasks with no heavy work load. Diet composition and caloric intake was monitored 1 week before and throughout the study. Dietary characteristics were balanced and identical for both test groups. All test subjects were members of the army, a fact that helped to standardize the experimental conditions. EBC (see below) was collected initially 2 weeks prior at sea level, on day 2 (24 h after arrival at altitude) and day 44 at altitude as well as at sea level 3 days after returning from altitude. Capillary blood samples for determination of hemoglobin concentration ([Hb]) and cubital-venous blood samples for reduced GSH were taken initially 2 weeks prior at sea level (670 m), and on days 2 (24 h after arrival at altitude), 13, 24, 34 and 44 at altitude (2,800 m). All data were collected in the morning after a minimum period of 12 h rest for the test subjects.
Exhaled breath condensate (EBC) collection and analysis
Exhaled breath condensate was collected with a self-constructed device as described earlier (Araneda et al. 2005). In brief, prior to the sampling procedure, the subject’s mouth was rinsed with distilled water and the nostrils were blocked by cotton stoppers. Expired air was led from a face mask with attached heated tube (40°C) into a glass collector placed in ice at −5°C. Using a trap, saliva was collected in the bottom of the face mask that was not connected to the condenser. About 1.5 ml EBC was collected during a period of 20 min and immediately stored in liquid nitrogen until analyzed.
Hydrogen peroxide (H2O2)
Hydrogen peroxide concentration was measured in EBC with the Ecocheck™ (Flit GmbH, Berlin, Germany) by using a biosensor as described previously (Gerritsen et al. 2005). In brief, 500 μl EBC are mixed with 500 μl dilution buffer to adjust the phosphate concentration. H2O2 is reduced enzymatically by a peroxidase on the sensor. The electrons are transferred to the sensor electrode by a redox pair where they induce a voltage change, which is proportional to the H2O2 concentration.
8-iso prostaglandin F2α (8-iso PGF2α)
8-iso PGF2α concentration was determined by an enzyme immunoassay kit (Cayman Chemical, Ann Arbor, MI, USA), previously tested for its utility to assess 8-iso PGF2α in EBC (Montuschi et al. 2000). The test kit was modified according to the manufacturer’s instructions to reach a lower limit of detection of 1 pg/ml.
Blood analysis
Capillary blood samples were immediately analyzed with the GEM Oxygenation Portable Laboratory (OPL), (Instrumentation Laboratory, Munich, Germany) to measure [Hb].
Reduced glutathione (GSH)
GSH was determined in whole blood by analyzing the concentration in erythrocytes that is about 500 times higher than in plasma (Jones 2002). Hundred microliter venous blood was added to 800 μl of precipitation solution containing: 1.67 g of glacial metaphosphoric acid, 0.2 g Na2EDTA and 30 g NaCl in 100 ml ddH2O. Samples were left at room temperature for 10 min to reach complete hemoglobin denaturation and were then frozen in liquid nitrogen.
Ellman’s reagent (DTNB) was used to evaluate the nonprotein thiol GSH as described earlier (Tietze 1969). After centrifugation (15 min at 5,000g), supernatants were assayed for nonprotein thiols. GSH was measured using a Lambda 25 UV/VIS spectrometer (Perkin Elmer). Absorption of complexes of GSH with DTNB was measured at 412 nm and a calibration curve was constructed for each experiment using a commercially available GSH standard (Sigma).
Statistics
Statistical analysis was carried out using GraphPad InStat software version 3.06 (San Diego, CA). The nonparametric Friedman test for repeated measures was performed to compare groups by time followed by Dunn’s post-hoc test. Comparisons between the single groups (trained vs. sedentary) were analyzed by applying the Mann–Whitney test. The level of statistical significance was set at p ≤ 0.05.
Results
Increased levels of oxidative stress markers were found at altitude in both groups, trained and sedentary (Table 1). However, we did not detect any significant differences in H2O2, 8-iso PGF2α and GSH levels between the trained and sedentary group at sea level or at altitude. Therefore, the subjects’ data were pooled to examine the effect of hypoxia on redox balance. As depicted in Fig. 1, H2O2 significantly increased after 44 days at altitude from 60 nmol/l (median), 40–106 nmol/l (interquartile range) at sea level to 131, 118–167 nmol/l, p ≤ 0.05. The level of H2O2 remained elevated even 3 days after returning to sea level (129, 92–194 nmol/l, p ≤ 0.05) when compared to sea level (-pre) values. 8-iso PGF2α showed a tendency to increase during the stay at altitude (day 2 vs. day 44, from 2.48, 1.90–3.24 to 3.17, 2.37–4.31, pg/ml, Fig. 2) and significantly decreased upon returning to sea level (1.84, 1.58–2.23, pg/ml, p ≤ 0.05). Finally, Fig. 3 demonstrates that an acute exposure to hypoxia resulted in significantly elevated GSH levels [sea level (-pre) vs. day 2, from 4.82, 4.12–5.33 to 5.38, 5.01–6.08, μmol/g Hb, p ≤ 0.05]. GSH levels significantly decreased throughout the stay at altitude (day 2 vs. day 44, from 5.38, 5.01–6.08 to 3.87, 3.32–4.66 μmol/g Hb, p ≤ 0.001).

Changes in hydrogen peroxide (H2O2) in response to prolonged hypoxia. Sea level (-pre) values, 2 weeks prior to altitude exposure; sea level (-post) values, 3 days post altitude exposure. Values are given as box plots indicating the median (center horizontal line), interquartile range from the 25th to the 75th percentile (box), and the 10th and 90th percentiles (whiskers). Outliers are presented as circles. Asterisk indicates significant differences from sea level (-pre) values, p ≤ 0.05. Pooled data from ten trained and five sedentary control subjects are shown

Changes in 8-iso prostaglandin F2α (8-iso PGF2α) in response to prolonged hypoxia. Sea level (-pre) values, 2 weeks prior to altitude exposure; sea level (-post) values, 3 days post altitude exposure. Values are given as box plots indicating the median (center horizontal line), interquartile range from the 25th to the 75th percentile (box), and the 10th and 90th percentiles (whiskers). Outliers are presented as circles. Asterisk indicates significant differences from sea level (-pre) values, hash different from altitude values—day 2, plus different from altitude values—day 44, p ≤ 0.05. Pooled data from ten trained and five sedentary control subjects are shown

Time course of reduced glutathione (GSH) at moderate altitude. Sea level (-pre) values, 2 weeks prior to altitude exposure; sea level (-post) values, 3 days post altitude exposure. Values are given as box plots indicating the median (center horizontal line), interquartile range from the 25th to the 75th percentile (box), and the 10th and 90th percentiles (whiskers). Outliers are presented as circles. Asterisk indicates significant differences from sea level (-pre) values, p ≤ 0.05; hash different from altitude values—day 2, p ≤ 0.05, double hash p ≤ 0.01, triple hash p ≤ 0.001. Pooled data from ten trained and five sedentary control subjects are shown
Discussion
Contrary to our hypothesis, the present study determined that the effects of two combined oxidative stimuli, chronic hypoxia and training, did not demonstrate a cumulative effect on the redox state in endurance trained athletes compared to sedentary controls. We observed, however, altitude-induced alterations with significantly increased levels of H2O2 in EBC at altitude and after return to sea level, a tendency to increased 8-iso PGF2α levels in EBC at altitude as well as a depletion of the antioxidant GSH in blood. Our results suggest that living at moderate altitude for an extended period of time is accompanied by a long-term increase in lung oxidative stress independent of strenuous physical exertion in trained subjects.
Earlier studies have shown increased levels of oxidative stress associated with physical work at moderate altitude over a shorter period of time than our study, 11–24 days (Araneda et al. 2005; Chao et al. 1999; Pfeiffer et al. 1999; Schmidt et al. 2002; Vasankari et al. 1997), and 12 hypoxic training sessions during 6 weeks (Pialoux et al. 2006). However, in those studies no sedentary control groups were investigated at altitude and the objective was mainly to investigate the influence of antioxidant supplementation.
It is known that acute exercise in hypoxia (Araneda et al. 2005; Bailey et al. 2001; Joanny et al. 2001; Moller et al. 2001; Pialoux et al. 2006) and normoxia (Bailey et al. 2001; Bailey et al. 2004) increases oxidative stress compared to resting conditions. On the other hand, regular intermittent hypoxic training was leading to attenuated increases in lipid peroxidation and reduced plasma malondialdehyde due to mobilization of α-tocopherol compared with normoxic training (Bailey et al. 2001). The observation that there was no difference between sedentary and endurance trained subjects might be explained by the lower exercise-induced oxidative stress after intermittent hypoxic training. This suggests that controlled ROS generation is required to initiate adaptation to physical stress.
There is evidence that normoxic exercise training results in increased cellular concentrations of antioxidants (Powers et al. 1999). Endurance training can elevate antioxidant enzyme activities in erythrocytes (Miyazaki et al. 2001). It also seems to be capable of reducing DNA damage in white blood cells after exhaustive exercise (Niess et al. 1996), and strengthens GSH dependent tissue antioxidant defense by increasing cellular concentrations of GSH in skeletal muscles (Powers et al. 1999; Sen 1999). However, we found no differences in GSH sea level values between our trained and sedentary subjects. At moderate altitude, both groups followed the same antioxidant pattern. Hypoxic exposure did lead to an increase in GSH-concentration after 24 h; and, GSH levels decreased to normoxic levels during the stay at altitude. Earlier we described a rapid increase in mouse erythrocytes GSH levels on 0.5% O2 hypoxic exposure in vitro (Bogdanova et al. 2003), and others showed an increase in mouse muscle reduced GSH with acute hypoxia equivalent to an altitude of 7,000 m (Magalhaes et al. 2004), a result that correlates with the observation of suppressed ROS production in hepatocytes primary cultures in response to acute hypoxia (Bogdanova et al. 2005). Chronic hypoxic exposure reduces erythrocytes GSH content (Singh et al. 2001) and triggers activation of glutathione cycle related enzyme activity (Magalhaes et al. 2005) leading to improved antioxidant capacity of blood.
Permanent residence at high altitude was accompanied by increased oxidative stress (Jefferson et al. 2004), and 1 day of high altitude hypoxia in sedentary lowlanders was associated with increased steady-state levels of oxidative DNA damage (Moller et al. 2001). However, prolonged hypoxic exposure in lowlanders showed an adaptive response to oxidative stress. After 8 weeks at high altitude, similar oxidative DNA damage was found as it was seen at sea level (Lundby et al. 2003). Moreover, after 13 months, the antioxidant defense was upregulated, e.g. as identified by elevated whole blood GSH levels (Vij et al. 2005).
In our study, oxidative stress persisted upon returning to sea level for 3 days. Indeed, there are reports of continued increased levels of lipid peroxidase (Joanny et al. 2001), increased levels of H2O2 (Araneda et al. 2005) after a prolonged stay at high altitude, and increased polarity of the erythrocyte membrane bilayer surface with increased formation of malondialdehyde after a short-term stay at moderate altitude (Gonzalez et al. 2005).
In conclusion, 6 weeks of endurance training at moderate altitude did not result in an additional long-term increase in lung oxidative stress in trained compared to sedentary subjects. In addition, oxidative stress remained elevated upon returning to sea level. These observations suggest that the increased oxidative stress is due to exposure to moderate altitude. Our data imply that a protective adaptation to physical stress occurred in athletes that adjusts the systemic homoeostasis. However, the physiological consequences of increased lung oxidative stress at moderate altitude remain to be elucidated.
References
Araneda OF, Garcia C, Lagos N, Quiroga G, Cajigal J, Salazar MP, Behn C (2005) Lung oxidative stress as related to exercise and altitude lipid peroxidation evidence in exhaled breath condensate: a possible predictor of acute mountain sickness. Eur J Appl Physiol 95:383–390. doi:10.1007/s00421-005-0047-y
Askew EW (2002) Work at high altitude and oxidative stress: antioxidant nutrients. Toxicology 180:107–119. doi:10.1016/S0300-483X(02)00385-2
Bailey DM, Davies B, Young IS (2001) Intermittent hypoxic training: implications for lipid peroxidation induced by acute normoxic exercise in active men. Clin Sci (Lond) 101:465–475. doi:10.1042/CS20010065
Bailey DM, Young IS, McEneny J, Lawrenson L, Kim J, Barden J, Richardson RS (2004) Regulation of free radical outflow from an isolated muscle bed in exercising humans. Am J Physiol Heart Circ Physiol 287:H1689–H1699. doi:10.1152/ajpheart.00148.2004
Bogdanova AY, Ogunshola OO, Bauer C, Gassmann M (2003) Pivotal role of reduced glutathione in oxygen-induced regulation of the Na(+)/K(+) pump in mouse erythrocyte membranes. J Membr Biol 195:33–42. doi:10.1007/s00232-003-2042-8
Bogdanova A, Grenacher B, Nikinmaa M, Gassmann M (2005) Hypoxic responses of Na+/K+ ATPase in trout hepatocytes. J Exp Biol 208:1793–1801. doi:10.1242/jeb.01572
Chao WH, Askew EW, Roberts DE, Wood SM, Perkins JB (1999) Oxidative stress in humans during work at moderate altitude. J Nutr 129:2009–2012
Chapman R, Levine BD (2007) Altitude training for the marathon. Sports Med 37:392–395. doi:10.2165/00007256-200737040-00031
Gerritsen WB, Zanen P, Bauwens AA, van den Bosch JM, Haas FJ (2005) Validation of a new method to measure hydrogen peroxide in exhaled breath condensate. Respir Med 99:1132–1137. doi:10.1016/j.rmed.2005.02.020
Gonzalez G, Celedon G, Escobar M, Sotomayor C, Ferrer V, Benitez D, Behn C (2005) Red cell membrane lipid changes at 3,500 m and on return to sea level. High Alt Med Biol 6:320–326. doi:10.1089/ham.2005.6.320
Heinicke K, Heinicke I, Schmidt W, Wolfarth B (2005) A three-week traditional altitude training increases hemoglobin mass and red cell volume in elite biathlon athletes. Int J Sports Med 26:350–355. doi:10.1055/s-2004-821052
Horvath I, Hunt J, Barnes PJ, Alving K, Antczak A, Baraldi E, Becher G, van Beurden WJ, Corradi M, Dekhuijzen R, Dweik RA, Dwyer T, Effros R, Erzurum S, Gaston B, Gessner C, Greening A, Ho LP, Hohlfeld J, Jobsis Q, Laskowski D, Loukides S, Marlin D, Montuschi P, Olin AC, Redington AE, Reinhold P, van Rensen EL, Rubinstein I, Silkoff P, Toren K, Vass G, Vogelberg C, Wirtz H (2005) Exhaled breath condensate: methodological recommendations and unresolved questions. Eur Respir J 26:523–548. doi:10.1183/09031936.05.00029705
Jefferson JA, Simoni J, Escudero E, Hurtado ME, Swenson ER, Wesson DE, Schreiner GF, Schoene RB, Johnson RJ, Hurtado A (2004) Increased oxidative stress following acute and chronic high altitude exposure. High Alt Med Biol 5:61–69. doi:10.1089/152702904322963690
Joanny P, Steinberg J, Robach P, Richalet JP, Gortan C, Gardette B, Jammes Y (2001) Operation Everest III (Comex’97): the effect of simulated sever hypobaric hypoxia on lipid peroxidation and antioxidant defence systems in human blood at rest and after maximal exercise. Resuscitation 49:307–314. doi:10.1016/S0300-9572(00)00373-7
Jones DP (2002) Redox potential of GSH/GSSG couple: assay and biological significance. Methods Enzymol 348:93–112. doi:10.1016/S0076-6879(02)48630-2
Levine BD, Stray-Gundersen J (1997) “Living high-training low”: effect of moderate-altitude acclimatization with low-altitude training on performance. J Appl Physiol 83:102–112
Lundby C, Pilegaard H, van Hall G, Sander M, Calbet J, Loft S, Moller P (2003) Oxidative DNA damage and repair in skeletal muscle of humans exposed to high-altitude hypoxia. Toxicology 192:229–236. doi:10.1016/S0300-483X(03)00328-7
Magalhaes J, Ascensao A, Soares JM, Neuparth MJ, Ferreira R, Oliveira J, Amado F, Duarte JA (2004) Acute and severe hypobaric hypoxia-induced muscle oxidative stress in mice: the role of glutathione against oxidative damage. Eur J Appl Physiol 91:185–191. doi:10.1007/s00421-003-0972-6
Magalhaes J, Ascensao A, Marques F, Soares JM, Ferreira R, Neuparth MJ, Duarte JA (2005) Effect of a high-altitude expedition to a Himalayan peak (Pumori, 7,161 m) on plasma and erythrocyte antioxidant profile. Eur J Appl Physiol 93:726–732. doi:10.1007/s00421-004-1222-2
Makris D, Paraskakis E, Korakas P, Karagiannakis E, Sourvinos G, Siafakas NM, Tzanakis N (2007) Exhaled breath condensate 8-isoprostane, clinical parameters, radiological indices and airway inflammation in COPD. Respiration 75:138–144. doi:10.1159/000106377
Miyazaki H, Oh-ishi S, Ookawara T, Kizaki T, Toshinai K, Ha S, Haga S, Ji LL, Ohno H (2001) Strenuous endurance training in humans reduces oxidative stress following exhausting exercise. Eur J Appl Physiol 84:1–6. doi:10.1007/s004210000342
Moller P, Loft S, Lundby C, Olsen NV (2001) Acute hypoxia and hypoxic exercise induce DNA strand breaks and oxidative DNA damage in humans. FASEB J 15:1181–1186. doi:10.1096/fj.00-0703com
Montuschi P (2005) Exhaled breath condensate analysis in patients with COPD. Clin Chim Acta 356:22–34. doi:10.1016/j.cccn.2005.01.012
Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ (2000) Exhaled 8-isoprostane as an in vivo biomarker of lung oxidative stress in patients with COPD and healthy smokers. Am J Respir Crit Care Med 162:1175–1177
Niess AM, Hartmann A, Grunert-Fuchs M, Poch B, Speit G (1996) DNA damage after exhaustive treadmill running in trained and untrained men. Int J Sports Med 17:397–403. doi:10.1055/s-2007-972868
Pfeiffer JM, Askew EW, Roberts DE, Wood SM, Benson JE, Johnson SC, Freedman MS (1999) Effect of antioxidant supplementation on urine and blood markers of oxidative stress during extended moderate-altitude training. Wilderness Environ Med 10:66–74
Pialoux V, Mounier R, Ponsot E, Rock E, Mazur A, Dufour S, Richard R, Richalet JP, Coudert J, Fellmann N (2006) Effects of exercise and training in hypoxia on antioxidant/pro-oxidant balance. Eur J Clin Nutr 60:1345–1354. doi:10.1038/sj.ejcn.1602462
Powers SK, Ji LL, Leeuwenburgh C (1999) Exercise training-induced alterations in skeletal muscle antioxidant capacity: a brief review. Med Sci Sports Exerc 31:987–997. doi:10.1097/00005768-199907000-00011
Psathakis K, Mermigkis D, Papatheodorou G, Loukides S, Panagou P, Polychronopoulos V, Siafakas NM, Bouros D (2006) Exhaled markers of oxidative stress in idiopathic pulmonary fibrosis. Eur J Clin Invest 36:362–367. doi:10.1111/j.1365-2362.2006.01636.x
Schmidt MC, Askew EW, Roberts DE, Prior RL, Ensign WY Jr, Hesslink RE Jr (2002) Oxidative stress in humans training in a cold, moderate altitude environment and their response to a phytochemical antioxidant supplement. Wilderness Environ Med 13:94–105
Sen CK (1995) Oxidants and antioxidants in exercise. J Appl Physiol 79:675–686
Sen CK (1999) Glutathione homeostasis in response to exercise training and nutritional supplements. Mol Cell Biochem 196:31–42. doi:10.1023/A:1006910011048
Singh SN, Vats P, Kumria MM, Ranganathan S, Shyam R, Arora MP, Jain CL, Sridharan K (2001) Effect of high altitude (7,620 m) exposure on glutathione and related metabolism in rats. Eur J Appl Physiol 84:233–237. doi:10.1007/s004210170010
Tietze F (1969) Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues. Anal Biochem 27:502–522. doi:10.1016/0003-2697(69)90064-5
Vasankari TJ, Kujala UM, Rusko H, Sarna S, Ahotupa M (1997) The effect of endurance exercise at moderate altitude on serum lipid peroxidation and antioxidative functions in humans. Eur J Appl Physiol Occup Physiol 75:396–399. doi:10.1007/s004210050178
Vij AG, Dutta R, Satija NK (2005) Acclimatization to oxidative stress at high altitude. High Alt Med Biol 6:301–310. doi:10.1089/ham.2005.6.301
Acknowledgments
We thank the subjects from the Chilean army for their enthusiastic contribution and gratefully acknowledge technical support from Gisela Kuhn, Stephan Keller and Bianca Rohde-Saam. Johannes Vogel is acknowledged for fruitful discussions. This work has been supported by the Eidgenössische Sportkommission (ESK) des Bundesamtes für Sport (BASPO), by the Swiss National Science Foundation (3100A0-100214) and by the Fensterfabrik Albisrieden, Zurich, Switzerland (U. & G. Frei).
Author information
Authors and Affiliations
Corresponding author
Additional information
The contribution of Max Gassmann and Katja Heinicke who are the senior authors was equivalent.
Rights and permissions
About this article
Cite this article
Heinicke, I., Boehler, A., Rechsteiner, T. et al. Moderate altitude but not additional endurance training increases markers of oxidative stress in exhaled breath condensate. Eur J Appl Physiol 106, 599–604 (2009). https://doi.org/10.1007/s00421-009-1014-9
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-009-1014-9