
Abstract
This study examined whether circadian rhythms affect lactate threshold (Thlac) during rowing exercise. Eleven male, endurance-trained athletes [mean (SD) age 29.5 (6.1) years] rowed at 0200, 0600, 1000, 1400, 1800 and 2200 hours under the same experimental conditions. Capillary blood (25 μl) was obtained from the tip of the toe during the last 30 s of a continuous, multi-stage, 3-min, incremental protocol on the Concept II ergometer. To determine Thlac, a curve-fitting procedure (the D max method), a visual method (Thlac-vis) and the fixed blood lactate concentration of 4.0 mmol l−1 (Thlac-4 mM) were used. Circadian rhythms were apparent for oxygen consumption and heart rate at Thlac using the D max method (P=0.02 and P=0.04 respectively), with the acrophases at 2139 hours and 2032 hours respectively coinciding in phase with that of core body temperature. The conclusion is that tests should be completed at the same time of day at which the athlete usually trains, to ensure precision of Thlac determination, especially when the D max method is used to determine Thlac.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Circadian rhythms constitute changes in physiological activity displaying a period of ~24 h (Minors and Waterhouse 1981). The most distinctive rhythm observed is that of core body temperature, with a peak occurring at ~1800 hours and a trough at ~0600 hours (Åkerstedt 1979). The circadian rhythm in temperature persists during submaximal exercise (Reilly and Brooks 1982), as does that of heart rate (HR; Reilly et al. 1984), oxygen consumption (V̇Ȯ2; Reilly and Brooks 1982; Giacomoni et al. 1999) and pulmonary ventilation (V̇ E; Reilly and Brooks 1982). The lactate threshold (Thlac), defined as the point at which blood lactate increases exponentially with exercise intensity (Yeh et al. 1983), has not been investigated thoroughly with respect to circadian rhythms.
Marti et al. (1998) tested 20 well-trained runners and found no significant differences in submaximal running speed at Thlac when subjects were tested between 0830 hours and 0900 hours compared to responses between 1030 hours and 1130 hours. It is unlikely that a diurnal variation will be revealed since only two time points were compared, and a difference of only 2 h separated these time points. In addition, Thlac was recorded as the running velocity inducing a blood lactate concentration of 4 mmol l−1 (Thlac-4 mM). The usefulness and reproducibility of using this method for representing a maximum lactate steady state, and distinguishing differences between individuals have been questioned (Stegmann et al. 1981). It may be more appropriate to assess whether the shape of the lactate response curve changes rhythmically with time of day, rather than measuring a fixed blood lactate concentration, which could be affected by changes in diet or training status (Hughes et al. 1982; Yoshida 1984). Sekir et al. (2002) studied the effect of time of day on the relationship between Thlac and the ventilatory threshold (Thvent). They reported no significant time of day effects for Thlac using the log–log method, or for Thvent using the V-slope method. There was, however, some variation due to time of day in the strength of the relationship between Thlac and Thvent (determined using correlation analysis) when exercise time and V̇ E were used as variables to describe the thresholds. Only three measurements were taken at 0700 hours, 1400 hours and 1900 hours.
If a circadian rhythm exists for blood lactate response to exercise, then it might be explained by increases in catecholamine activity as suggested by Deschenes et al. (1998), since catecholamines, particularly adrenaline, follow very similar patterns in response to exercise as those found for blood lactate (Mazzeo and Marshall 1989). A pronounced circadian variation in catecholamine activity has been observed at rest (Åkerstedt 1979) with a peak in the early afternoon and a trough occurring during the night, independent of sleep patterns, although this rhythm may be obscured by exercise (Stephenson et al. 1989). An alternative explanation for a circadian rhythm in blood lactate concentration, offered by Dalton et al. (1997), was that it might be caused partly by circadian changes in core body temperature. An elevation in body temperature would increase activity of enzymes such as phosphofructokinase and lactate dehydrogenase, which could in turn increase lactate production during exercise.
It has not been firmly established whether a circadian variation exists for blood lactate concentration in response to exercise, or whether exercise intensity at Thlac is altered with time of day. The finding of changes in metabolism (V̇Ȯ2 during light to moderate exercise, e.g. Reilly and Brooks 1982) leaves open the possibility of a circadian rhythm in blood lactate. Circadian rhythms could influence evaluation of fitness, establishment of training intensities and evaluation of performance improvement, when these are assessed using blood lactate concentration as criterion. The purpose of this study was to investigate the existence of circadian rhythms in Thlac. The investigation required the measurement of a number of variables [exercise intensity, V̇Ȯ2, HR, ratings of perceived exertion (RPE) and blood lactate concentration] at Thlac at six different times throughout the day during an incremental exercise protocol on a rowing ergometer. Rowing was selected, since exercise intensity at Thlac is an important determinant of 2,000-m rowing performance (Ingham et al. 2002).
Methods
Eleven healthy male subjects, with a mean (SD) age of 29.5 (6.1) years, height of 1.80 (0.05) m and body mass of 79.5 (6.7) kg, were recruited to take part in the study. Seven subjects were endurance-trained athletes who regularly used the Concept II rowing ergometer (Model C, Nottingham, UK) as part of their training, and the remaining four were competitive, club-standard rowers. Subjects were free of medication and injury, and were not experiencing sleep deprivation or insomnia at the time of testing. Oral and written consent was obtained, after having fully informed subjects of the procedures. The study was approved by the Human Ethics Committee at Liverpool John Moores University. This experiment complies with the current laws of the countries of the UK.
Procedures and protocol
Subjects were tested on six separate occasions at 0200, 0600, 1000, 1400, 1800 and 2200 hours. A preliminary test session was carried out, in order to familiarise the subjects fully with the procedures, and to determine the protocol to be used on subsequent occasions. The order of testing was counter-balanced between subjects, and trials were separated by a minimum of 2 days to allow time for recovery. For any one subject the six tests were completed within 3 months, with no changes in habitual physical activity or training being noted during this period. Testing was also carried out on different days of the week to minimise any possible circaseptan influence. Subjects were encouraged to adhere as closely as possible to their usual sleep–wake cycle, with a minimum of 6 h of sleep being taken on the night preceding each test. Subjects did not sleep before the test at 0200 hours, but did sleep before that at 0600 hours and were awake for at least 1 h prior to the commencement of testing.
Subjects were encouraged to refrain from strenuous physical activity, to avoid alcohol and caffeine and to ensure adequate food intake and hydration within the 48 h preceding each test. Physical activity over 48 h and dietary intake 24 h prior to each of the six sessions were recorded, and diet and activity patterns were replicated as closely as possible. The last meal before testing was a light snack of approximately 1,670 kJ (400 kcal) consumed at least 3 h before testing and taken with 250 ml of pure orange juice (Reilly and Brooks 1982; Dalton et al. 1997). Before the test session at 0600 hours, subjects consumed the snack prior to going to bed. During the familiarisation visit, subjects completed questionnaires to ascertain chronotype (Horne and Östberg 1976), usual time of day of training, motivation for exercise and ratings of fatigue and sleep.
On reporting to the laboratory, subjects rested in a seated position for 10 min, and data (ventilatory variables, HR, core body temperature and blood lactate concentration) were collected in the final minute of rest. The protocol used was the same as that described by Forsyth and Reilly (2003). Subjects completed a warm-up for 5 min at an intensity corresponding to between 50% and 60% of maximum oxygen consumption (V̇Ȯ2max). This intensity and the initial intensity of the protocol were determined during the familiarisation visit. After the warm-up subjects rested for 5 min, remaining seated. The mean initial intensity for the protocol was 152.4 (SD 14.7, range 128.2–179.4) W, which corresponded to 57.4 (SD 4.1)% V̇Ȯ2max. The intensity was then increased by 18.4 (SD 2.4) W, corresponding to a mean increase in V̇Ȯ2 of 4.06 (SD 0.63) ml kg−1 min−1, every 3 min, until subjects reached volitional exhaustion or were unable to maintain the required power. Mean test duration was 19.8 (SD 2.4, range 16–24) min. Subjects were encouraged to maintain a stroke rate of between 24 and 32 strokes min−1, this range being wide enough to cater for differences in rowing ability and style, yet restrictive enough to allow for comparability. Mean stroke rate was 26.4 (SD 2.92) strokes min−1. The performance monitor on the Concept II ergometer displays the stroke rate, as well as exercise intensity (500-m split time), which subjects were able to monitor and control. The drag factor was 121 for lightweights and 132 for heavyweights.
Analysis of variables
Ventilatory variables were recorded at rest and throughout the incremental test using an on-line, open-circuit gas analyser (Aerosport TEEM 100, Ann Arbor, Mich.), calibrated prior to exercise with known concentrations of gas, and by a 3-l syringe (GDX Corporation). The low-flow pneumotachograph was used to collect resting data, and the medium-flow pneumotachograph was used during exercise.
HR was monitored throughout by means of short-range radio telemetry (Cardiosport Excel Sport PC), and RPE (Borg 1970) was rated at the end of each increment. Rectal temperature (T rec) was measured at rest using a rectal probe (mini-thermistor, Grant Instruments, Shepreth, UK) inserted to a distance of approximately 10 cm beyond the anal sphincter. The data were recorded continuously through a Squirrel 1000 data logger (Grant Instruments). Rectal temperature was chosen since it is a reliable marker of circadian rhythm (Minors and Waterhouse 1981).
Capillary blood (25 μl) was removed from the tip of a pre-warmed toe during the last 30 s of each increment. Removing blood from the toe enables the subject to carry on rowing, without interruption to the protocol (Forsyth and Farrally 2000). Samples were assessed immediately from lysed blood using a 1500 YSI lactate analyser (Yellow Springs Instruments, Ohio), which was calibrated with 5, 15 and 30 mmol l−1 standards.
Determination of Thlac
Methods used to detect Thlac included the D max method (Cheng et al. 1992), the visual method (Thlac-vis) and Thlac-4 mM. For the D max method, third order polynomial regressions of the variables used (V̇Ȯ2, HR, power and RPE) against blood lactate concentration were determined. The slope of the straight line formed by the two endpoints of each curve was calculated, and D max was defined as the maximal perpendicular distance from the curve to the straight line. The Thlac-vis method involved plotting blood lactate concentration against power, and was defined as the highest exercise intensity before a curvilinear increase in blood lactate concentration occurred. Two independent observers were consulted, with a third observer used, if the initial two observers were not in agreement as to where Thlac-vis occurred. Plots were coded to avoid investigator bias. This “threshold” was also described according to V̇Ȯ2, HR, RPE and blood lactate concentration. For Thlac-4 mM determination, plots of blood lactate concentration against all variables were drawn and values were interpolated from the curve. On the six test occasions, the protocol and procedures were replicated as closely as possible. Mean environmental temperature for all trials was 19.0 (SD 1.5)°C, and mean humidity was 50.6 (10.6)%.
Statistical analysis
All variables were analysed using a repeated measures analysis of variance (ANOVA) with time of day and subjects as factors. A cosinor analysis was then performed on group data using the curve-fitting model to detect circadian characteristics of the variables (Nelson et al. 1979). The analysis of variance was used to identify non-symmetrical time of day effects and to uncover irregularities that did not conform to a cosinor analysis, and to determine variability among subjects. The cosinor analysis was used to optimise statistical power, since data are smoothed, and to give a complete description of rhythm characteristics. The mesor of the rhythm was defined as that which occurred mid-way between the highest (the peak) and the lowest point (the trough or nadir). The amplitude of a rhythm was defined as the difference between mesor and either extreme, peak or trough, and the acrophase as the time of the occurrence of the peak (Minors and Waterhouse 1981). Individual acrophases for the different variables were compared using a paired samples t-test. Limits of agreement (Bland and Altman 1986) were used to compare measurements taken 12 h apart between 0200 hours and 1400 hours, 0600 hours and 1800 hours, and between 1000 hours and 2200 hours, and effect sizes were determined as a means of further describing data (Mullineaux et al. 2001). A probability of 0.05 was used to represent significance.
Results
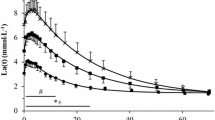
A circadian rhythm was observed for T rec, with the acrophase occurring at 1728 hours (Table 1). Cosinor analysis and analysis of variance revealed circadian rhythms for V̇Ȯ2 and HR at Thlac using the D max method of determination (Figs. 1, 2). HR and V̇Ȯ2 at D max coincided in phase with that of T rec.

Mean and SD (error bars) in heart rate using the D max method (Cheng et al. 1992) at the six different testing occasions

Mean and SD (error bars) in oxygen consumption (V̇Ȯ 2 ) using the D max method at the six different testing occasions
All other methods for determining Thlac were independent of time of day, as were power, RPE and blood lactate concentration using the D max method. The effect size for RPE at Thlac, determined using the D max method, was large (0.90) and limits of agreement were wide, especially between 0600 hours and 1800 hours (Table 2). Mean values for RPE using the D max method were lower at 0600 hours [13.6 (SD 1.7)] than they were at 1800 hours [15.2 (SD 1.9)], although differences were not significant. Mean values for blood lactate concentration at Thlac using the D max method were also lowest at 0600 hours [3.31 (SD 0.68) mmol l−1] and highest at 1800 hours [3.59 (SD 0.91) mmol l−1], but again differences did not reach significance. Hence, the D max method for determining Thlac seemed to yield most variation due to time of day compared to when using the other methods (Thlac-vis and Thlac-4 mM), although these variations only followed a typical circadian pattern when V̇Ȯ2 and HR were used as variables to describe the threshold.
There was little difference in how subjects rated their motivation for exercise, although four of the subjects stated that their motivation was poor at 0200 hours, compared to only one subject stating this at other times. Six subjects also reported feeling more fatigued at 0200 hours compared to the other times of testing. Mean sleep duration prior to the testing at 0600 hours was less (6 h) than that taken before the other test occasions, which were all greater than 6.6 h, although quality of sleep was rated similarly across all trials. The reason for mean sleep time being less before the testing at 0600 hours, was mainly because one of the subjects only slept for 5 h. All other subjects slept for the recommended minimum of 6 h.
According to the Horne and Östberg (1976) questionnaire, there was a mix of chronotypes, with four of the subjects rated as “neither a lark nor an owl but intermediate in chronotype”. Four of the subjects were rated as “moderately a lark”, two as “moderately an owl”, and one as “definitely an owl”. One subject trained habitually in the afternoon. The remaining subjects trained in the evening, between 1700 hours and 2000 hours.
Discussion
Circadian rhythms were apparent for V̇Ȯ2 and HR at Thlac when the D max method was used, with the acrophases of these rhythms occurring at 2139 hours and 2032 hours respectively (Table 1). For HR using the D max method, mean values were lowest at 0600 hours [157 (SD 10) beats min−1] and highest at 2200 hours [163 (SD 5) beats min−1]. The finding that HR at Thlac when using D max varies according to time of day is of importance, since this variable is commonly used as an indicator of training intensity. Hence, if using HR at D max to determine Thlac, training intensity could be set too high when the test is carried out in the late evening compared to when carried out at other times of the day. However, the amplitude of this rhythm was only 2 beats min−1, and the mean difference in HR between the test session at 0600 hours and that at 2200 hours was 6 beats min−1. These differences are small when considering that HR may alter by much more than this amplitude as a result of, for instance, anxiety experienced during competition. Nevertheless, in order to ensure accuracy is as high as possible, it is important to complete a test to determine Thlac using the D max method at the same time of day at which the athlete would normally train.
There was no significant time of day effect for HR using the other methods (Thlac-vis and Thlac-4 mM) to determine Thlac. However, the D max method is more reliable in repeat tests (Cheng et al. 1992; Forsyth and Reilly 2003). The probability that variation occurs due to time of day is, therefore, greater when using the D max method, since relatively less variation occurs due to confounding factors, such as experimental error, or due to any subjective assessment of where the “threshold” lies, as is the case for the Thlac-vis method. The D max method is more sensitive in identifying circadian rhythms in lactate response to exercise than are other methods of assessing Thlac.
Mean values for RPE and blood lactate using the D max method were suggestive of a circadian rhythm, both being lowest at 0600 hours [RPE: 13.6 (SD 1.7); blood lactate: 3.31 (SD 0.68) mmol l−1] and highest at 1800 hours [RPE: 15.2 (SD 1.9); blood lactate: 3.59 (SD 0.91) mmol l−1], but these fluctuations were not significant. The large effect size for RPE at Thlac using Dmax suggests that time of day could explain some of the variance. Limits of agreement for RPE and blood lactate concentration using the D max method were also relatively wide, especially between the test sessions at 0600 hours and 1800 hours (Table 2). The D max method for determining Thlac yields most circadian variation compared to the other methods of assessing Thlac, although cosinor variations were only apparent when V̇Ȯ2 and HR were used as variables to describe the “threshold”.
If there is some variation due to time of day when using the D max method to determine Thlac, then the rhythm is partly linked with changes in core body temperature, since HR and V̇Ȯ2 using D max both coincided with the acrophase in T rec. However, the concordance in phase is in itself insufficient to attribute a causal link to the relationship.
The finding, that Thlac when determined using the D max method occurred at a higher HR and V̇Ȯ2 later in the day, means that either the shape of the polynomial curve of blood lactate concentration in response to exercise, or the straight line joining the two end points, or both, have changed. Reasons for this include the following: a change in blood lactate concentration at the first and final stage of exercise, which will change the slope of the straight line as well as changing the shape of the polynomial; any change in blood lactate concentration throughout the test, which will change the shape of the polynomial; and any change in the variable, HR or V̇Ȯ2, that is employed in describing the “threshold”. Hence, when using the D max method, it is difficult to determine which of these possibilities account for the change in the determination of Thlac. It might be that the reason why only HR and V̇Ȯ2 follow a circadian rhythm when used to describe the “threshold” is because these two variables exhibit rhythmicity when exercise is submaximal (Reilly et al. 1984; Giacomoni et al. 1999), and it is not the blood lactate concentration per se that changes to any great extent. To determine whether the actual amount of blood lactate changes during exercise due to circadian effects, the results of using the Thlac-4 mM can be examined. Mean power at a blood lactate concentration of 4 mmol l−1 was lowest at 1800 hours [220.7 (SD 40.7) W] and highest at 0200 hours [227.6 (SD 24.0) W], indicating that blood lactate concentration was generally elevated at 1800 hours. The relatively lower mean blood lactate concentration at 0200 hours may be due to the preference for fat as substrate for metabolism at night (Schlierf 1978). Dalton et al. (1997) suggested that blood lactate concentration during submaximal exercise peaked in the afternoon, which would lead to Thlac occurring at a lower exercise intensity if determined using a fixed blood lactate concentration of 4 mmol l−1. The results of the current study agree with this finding, although changes in power at Thlac-4 mM were not large enough to produce a significant cosinor rhythm. Reproducibility of using the fixed blood lactate concentration has been questioned due to changes that occur as a result of relatively minor alterations in dietary intake and physical activity (Hughes et al. 1982; Yoshida 1984). There is, therefore, a preference for examining the shape of the lactate curve, which is less influenced by external factors, and may be more sensitive to changes as a result of circadian rhythms.
The limits of agreement shown in Table 2 give some indication of the amount of variation that is possible when determining Thlac. To highlight an example, limits of agreement for power at Thlac-vis ranged from −63.8 to +55.7 W between 0600 hours and 1800 hours. Since the mean increment for the test protocol was 18.4 W, the worst case scenario for an individual when using Thlac-vis could be that Thlac is over-estimated or under-predicted by as much as three exercise stages depending on what time of day the test is carried out. An error by this amount is unacceptable, especially when considering most individuals in this study only completed six to seven stages of exercise until exhaustion was reached. Determining Thlac visually is prone to errors in judgement over where the threshold lies and is therefore not as reliable as more objective methods. In this study, changes in power at Thlac-vis that occur due to time of day may have further added to the problem of reliability when using this method to determine Thlac.
Factors that might have influenced the present observations include individual levels of motivation and fatigue, and reported chronotype. At 0200 hours, four of the subjects stated their motivation for exercise was poor, compared to only one who reported poor motivation at all other times. This may have had some influence on test duration, and the ability to reach maximum exercise intensity at this time. The maximum mean power achieved was lowest at the 0200 hours and 0600 hours test sessions, being 250.0 (SD 25.8) W and 249.9 (SD 23.9) W respectively. The highest mean power achieved was at 1400 hours [256.6 (SD 30.7) W], although none of these differences was significant. Mean sleep duration is also a potential source of influence on performance at 0600 hours, since mean sleep duration was significantly less than that prior to the other test occasions. However, usually endurance performance is maintained even with much greater losses of sleep than this amount (Reilly and Deykin 1983). Hill et al. (1988) suggested that chronotype may affect performance depending on time of day, although Burgoon et al. (1992) found that HR and performance time exhibited a significant diurnal effect, but not an effect of chronotype during an incremental test to exhaustion. Most people fall within the range defined as intermediate in chronotype, although according to the results of Horne and Östberg’s questionnaire used in this study, seven subjects were outside this range, with one subject falling in the extreme of “definitely an owl”. This range could have led to discrepancies between individuals in attainment of peak performance. Although the subject described as “definitely an owl” did perform better at 0200 hours than he did on the other test occasions, generally those subjects described as “moderate owls” did not perform better in the evening, and the “moderate larks” did not perform better in the morning. There were no significant correlations between chronotype score and individual acrophases of peak power (r=−0.40) and test duration (r=−0.31). It seems that in general, these minor, non-significant differences in motivation, test duration, fatigue and chronotype did not have any major impact on the results.
This investigation has demonstrated the occurrence of time-of-day effects for V̇Ȯ2 and HR at Thlac using the D max method, which were found to be in phase with T rec. It is recommended that when using Thlac as a marker of performance change, tests are carried out at the same time of day to minimise circadian influences. In particular, when establishing training intensities using the D max method to determine Thlac, the test should be conducted at the same time of day at which the athlete usually trains or competes.
References
Åkerstedt T (1979) Altered sleep/wake patterns and circadian rhythms. Acta Physiol Scand Suppl 469:1–48
Bland JM, Altman DG (1986) Statistical methods for assessing agreement between two methods of clinical measurement. Lancet i:307–310
Borg G (1970) Perceived exertion as an indicator of somatic stress. Scand J Rehab Med 2–3:92–98.
Burgoon PW, Holland GJ, Loy SF, Vincent WJ (1992) A comparison of morning and evening “types” during maximum exercise. J Appl Sport Sci Res 6:115–119
Cheng B, Kuipers H, Snyder AC, Keizer HA, Jeukendrup A, Hesselink M (1992) A new approach for the determination of ventilatory and lactate thresholds. Int J Sports Med 13:518–522
Dalton B, McNaughton L, Davoren B (1997) Circadian rhythms have no effect on cycling performance. Int J Sports Med 18:538–542
Deschenes MR, Sharma JV, Brittingham KT, Casa DJ, Armstrong LE, Maresh CM (1998) Chronobiological effects on exercise performance and selected physiological responses. Eur J Appl Physiol 77:249–256
Forsyth JJ, Farrally MF (2000) A comparison of lactate concentration in plasma collected from the toe, ear and fingertip after a simulated rowing exercise. Br J Sports Med 34:35–38
Forsyth JJ, Reilly T (2003) A rowing protocol to determine lactate threshold. In: Reilly T, Marfell-Jones M (eds) Kinanthropometry VIII. Routledge, London, pp 55–65
Giacomoni M, Benard T, Gavarry O, Altare S, Fulgairette G (1999) Dirunal variations in ventilatory and cardiorespiratory responses to submaximal treadmill exercise in females. Eur J Appl Physiol 80:591–597
Hill DW, Cureton KJ, Collins MA, Grisham SC (1988) Effect of the circadian rhythm in body temperature on oxygen uptake. J Sports Med Phys Fitness 28:310–312
Horne JA, Östberg O (1976) A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int J Chronobiol 4:97–110
Hughes EF, Turner SC, Brooks GA (1982) Effects of glycogen depletion and pedalling speed on anaerobic threshold. J Appl Physiol 52:1598–1607
Ingham SA, Whyte GP, Jones K, Nevill AM (2002) Determinants of 2,000 m rowing ergometer performance in elite rowers. Eur J Appl Physiol 88:243–246
Marti B, Zundel R, Held T (1998) Beeinflusst ein früher vs. später vormittäglicher Testzeitpunkt die Ergebnisse eines Ausdauerleistungstests? Schweiz Z Sportmed Sporttraumatol 46:155–158
Mazzeo RS, Marshall P (1989) Influence of plasma catecholamines on the lactate threshold during graded exercise. J Appl Physiol 67:1319–1322
Minors DS, Waterhouse JM (1981) Circadian rhythms and the human. Wright, Bristol
Mullineaux PR, Bartlett RM, Bennett S (2001) Research design and statistics in biomechanics and motor control. J Sports Sci 19:739–760
Nelson W, Tong YL, Lee JK, Halberg F (1979) Methods for cosinor rhythmometry. Chronobiologia 6:305–323
Reilly T, Brooks GA (1982) Investigation of circadian rhythms in metabolic responses to exercise. Ergonomics 25:1093–1107
Reilly T, Deykin T (1983) Effects of partial sleep loss on subjective states, psychomotor and physical performance tasks. J Hum Mov Stud 9:157
Reilly T, Robinson G, Minors DS (1984) Some circulatory response to exercise at different times of day. Med Sci Sports Exerc 16:477–482
Schlierf G (1978) Diurnal variations in plasma substrate concentration. Eur J Clin Invest 8:59–60
Sekir U, Ozyener F, Gur H (2002) Effect of time of day on the relationship between lactate and ventilatory thresholds: a brief report. J Sports Sci Med 1:136–140
Stegmann H, Kindermann W, Schnabel A (1981) Lactate kinetics and individual anaerobic threshold. Int J Sports Med 2:160–165
Stephenson LA, Kolka MA, Francesconi R, Gonzalez RR (1989) Circadian variations in plasma renin activity, catecholamines and aldosterone during exercise in women. Eur J Appl Physiol 58:756–764
Yeh MP, Gardner RM, Adams TD, Yanowitz FG, Crapo RO (1983) “Anaerobic threshold”; problems of determination and validation. J Appl Physiol 55:1178–1186
Yoshida T (1984) Effect of exercise duration during incremental exercise on the determination of anaerobic threshold and the onset of blood lactate accumulation. Eur J Appl Physiol 53:196–199
Acknowledgement
The time and effort of the subjects who participated in this investigation are greatly appreciated.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Forsyth, J.J., Reilly, T. Circadian rhythms in blood lactate concentration during incremental ergometer rowing. Eur J Appl Physiol 92, 69–74 (2004). https://doi.org/10.1007/s00421-004-1059-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-004-1059-8