
Abstract
Prior to the emergence of agriculture in southwest Asia, increasingly sedentary human communities were experimenting with a diverse range of wild plants over a prolonged period. In some cases, this involved the cultivation of taxa that would go on to be domesticated and form the foundation of future agricultural economies. However, many forms of plant use did not follow this trajectory, and in multiple places farming was only taken up later as an established ‘package’ of crops and management practices. In this paper, we present new archaeobotanical evidence from the Early Neolithic site of Sheikh-e Abad in the central Zagros mountains of western Iran. Sheikh-e Abad is unique in being the only settlement known to date within southwest Asia that lies at an altitude above 1,000 m and which has occupation spanning the agricultural transition (9800–7600 bc). Thus, it provides a rare opportunity to examine pre-agricultural plant management strategies in an upland zone. Our analyses of the plant remains from Sheikh-e Abad suggest that from its earliest occupation the site’s inhabitants were ‘auditioning’ a group of locally available wild grasses, which ultimately were never domesticated. We discuss the possible reasons for this from a socio-ecological perspective, considering both the biology and ecology of the plant species in question, as well as the ways in which they were potentially managed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
It is increasingly accepted that agriculture had multiple ‘centres of origin’ within southwest Asia (Salamini et al. 2002; Fuller et al. 2012; Willcox 2013). This perspective is supported by a growing body of archaeobotanical data that documents the independent development of cultivation and the domestication of plants in several different regions during the earliest phases of the Neolithic, and possibly as far back as the Late Palaeolithic at Ohalo II, Israel (Snir et al. 2015). Pre-domestication cultivation—the cultivation of wild crop progenitors—has been proposed for 14 archaeological sites to date (Fig. 1a; Hopf 1983; van Zeist and de Roller 1994; Kislev 1997; Colledge 1998, 2001; Hillman 2000; Edwards et al. 2004; Weiss et al. 2006; Willcox et al. 2008; Kuijt and Finlayson 2009; Willcox 2012a; White and Makarewicz 2012; Arranz-Otaegui et al. 2016b), including recently for the first time the eastern Fertile Crescent (Riehl et al. 2013). However, pre-domestication cultivation represents only one of a number of possible pre-agricultural plant management strategies; moreover, the taxa that were domesticated and formed part of the ‘Neolithic crop package’ (Zohary and Hopf 2000; Weiss and Zohary 2011) account for only a small number of the wild plants that people were gathering, consuming and potentially managing during this time.

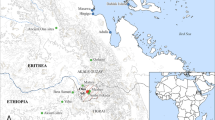
a Location of Sheikh-e Abad in the central Zagros mountains of western Iran and other key Early Neolithic sites across the Fertile Crescent; b elevation profile of a straight line transect between Chogha Golan and Sheikh-e Abad
While the process of domestication can be documented using morphological markers, such as tough rachis or seed size (Harlan et al. 1973; Nesbitt 2002; Fuller 2007), and the identification of arable weed floras used to infer the cultivation of wild crops (Colledge 1998; Weiss et al. 2006; Willcox 2012a), other potential plant management strategies may have involved markedly different forms of crop husbandry and methods of wild plant food production, including practices for which we have no modern analogues. Thus, reconstructing ‘non-domestication’ pre-agricultural plant management strategies remains problematic, especially given the complex nature of archaeobotanical assemblages and the multiple routes by which plants or their parts may have arrived at archaeological sites and been preserved. Moreover, charred plant assemblages are known to significantly under-represent wild plant resources (Matthews 2010, 2016; Colledge and Conolly 2014).
The interpretation of wild plant foods at Early Neolithic sites cannot rely on the uncritical assumption that the plant remains within an archaeobotanical assemblage are representative of those consumed. However, this inference may be strengthened if a taxon is recorded in large numbers and when other routes of entry into the assemblage are excluded, such as dung burning, crafts or construction. Data provided by modern ethnographic studies (e.g. Ertug-Yaras 1997) and information on the use of taxa recorded within regional floras can also help direct interpretations of wild plant foods, while more robust arguments may be made on the basis of archaeological evidence for food processing. For example at Jerf el Ahmar, Syria, the remains of charred seed cakes of Brassica/Sinapis sp. have been identified in situ on a saddle quern (Willcox 2002), suggesting their use as a food and/or oil. Similarly, at Ohalo II, the clustering of the remains of several wild grasses around a grinding stone has been cited in support of their interpretation as food at the site (Weiss et al. 2008). Morphological evidence for grains being broken prior to charring, consistent with the processing of cereals for consumption (e.g. by grinding) has also been reported at a number of sites across the region (Willcox 2002; Arranz-Otaegui et al. 2016a). In the absence of direct evidence for consumption from gut contents or coprolites, none of these other lines of evidence demonstrate unequivocally that a particular plant was used and managed as a food. However, such inferences may be substantiated on the basis of multiple lines of evidence.
A broader understanding of pre-agricultural plant management requires not only a critical evaluation of the archaeobotanical evidence available for wild plant foods, but also a consideration of the socio-ecological context in which experimentation with wild plants took place. Variation in local climate, vegetation and socio-cultural practices would all have been key factors in shaping early plant management, affording communities with different opportunities and constraints. In this paper, we present new evidence from the Early Neolithic site of Sheikh-e Abad in western Iran, where analysis of the charred macrobotanical assemblage has allowed us to trace long-term plant use over a 2,200-year period, including a sustained focus on a group of wild grasses with medium-sized grains as food. Notably, none of these wild grasses are crop progenitors and domesticated crops were only adopted later in the site’s occupation. At present, Sheikh-e Abad is unique amongst contemporary sites in southwest Asia, being the only settlement located in an upland zone, here defined as land above 1,400 m a.s.l., and with occupation spanning the agricultural transition (Fig. 1a). Furthermore, in contrast to other sites occupied during the initial phases of the Early Neolithic (PPNA or equivalent), Sheikh-e Abad is also located a considerable distance away from lower land. A straight-line distance of ca. 150 km separates it from elevations consistently below 1,000 m (Fig. 1b). The results we present here are therefore significant for our understanding of local variability in pre-agricultural plant management strategies, particularly for upland zones which are not represented within current models of agricultural origins.
Environmental setting
Sheikh-e Abad is located on a fertile alluvial plain at 1,430 m a.s.l. in a semi-arid region of the central Zagros mountains, 38 km north of the modern city of Kermanshah (Fig. 1a, b). The site lies at the boundary of the lower and elevated Zagros and is bounded by mountain peaks up to 3,000 m, providing access to a range of different environments. The climate of the study area is characterised as semi-arid with a Mediterranean component (Köppen–Geiger: Csa) and topography plays a key role in determining how this is expressed at a local level (Zohary 1973; Kehl 2009). Present-day precipitation around Sheikh-e Abad averages 437 mm per year and is sufficient for dry farming, with much of the land around the site currently under cultivation. However, precipitation is unevenly distributed throughout the year, with no rain falling during the hot summer months (June–October). Average temperatures range from 25.9 °C in July to a low of 1.3 °C in January, and between December and February minimum temperatures drop below freezing point.
Vegetation around Sheikh-e Abad is strongly influenced by the combined ecological factors of moisture and temperature, with both the cold winter period and hot dry summer inhibiting plant growth (Zohary 1973). In phytogeographical terms the study area falls into the xerophilous forest sub-division of the Irano-Turanian zone (Zohary 1973, p 87; Badripour 2006). Here, remnants of pistachio-oak woodland steppe are observed, rich in herbaceous plants and wild grasses, particularly perennials which form the basis of the vegetation from ca. 1,500 m (Badripour 2006, p 20). Bromus (B. tomentellus, B. cappadocicus), Festuca and Stipa (particularly S. barbata) are especially numerous, and other perennial grasses recorded here include Piptatherum holciforme (syn. Oryzopsis holciformis), Hordeum bulbosum, H. fragile, H. violaceum and Poa spp. The annual crop progenitor H. spontaneum (wild barley) is rare above 1,500 m due to its limited cold tolerance (Zohary and Hopf 2000, p 67), and the Zagros mountains represent a significant disruption in its natural range (Morrell and Clegg 2011). Triticum boeticum (wild einkorn) and T. dicoccoides (wild emmer) also prefer lower elevations (Bor 1968), placing Sheikh-e Abad outside their natural habitat areas, as illustrated in modern distribution maps for the wild cereals (Zohary and Hopf 2000; Willcox 2005; Fuller 2007).
It is unlikely that modern vegetation around the site provides a direct analogue for the archaeological past, especially in light of human and animal activity, particularly agriculture, which has significantly altered the natural vegetation of the study area (Zohary 1973, pp 608–610). Recent studies have also emphasised the role of varying moisture and CO2 availability in determining the distribution and yield of wild plants during the early Holocene (Cunniff et al. 2010; Riehl 2016). However, after the end of the Younger Dryas, as in the present day, wild grasses would have formed a major component of the vegetation across the Fertile Crescent (Riehl et al. 2012), replacing the existing chenopod-Artemisia steppe (Stevens et al. 2001). At the same time, Pistacia spp. (pistachio) and Quercus spp. (oak) pollen both appear in relatively low percentages in regional pollen cores, which correspond to those found today in the open oak woodland of the Zagros (Stevens et al. 2001), suggesting a similar level of arboreal cover.
Excavations at Sheikh-e Abad
Sheikh-e Abad was first recorded during a 2006 survey of the region by Yaghoub Mohammadifar and Abbass Motarjem and excavated in 2008 under phase 1 of the Central Zagros Archaeological Project (Matthews et al. 2013). At over 1 ha in extent, the site is dominated by the archaeological mound at its centre, which comprises approximately 10 m of almost exclusively Early Neolithic deposits. Excavations in 2008 focused on three areas that span this sequence: Trench 1 at the base of the mound, Trench 2 halfway up the mound and Trench 3 on the mound’s summit (Fig. 2a). Repeated layers of ash and debris from the earliest levels excavated attest to long-term continuity of activities at the site. Radiocarbon determinations on plant material from the lowest levels of Trench 1 returned a range of 9810–9240 cal bc, making occupation here broadly contemporary with the end of the Younger Dryas and the beginning of the Levantine Pre-Pottery Neolithic A (PPNA) cultural period (Kuijt and Goring-Morris 2002), while dates from Trench 2 (7960 cal bc) and Trench 3 (7590 cal bc) are coeval with the Levantine PPNB (Table 1). This makes the site one of the oldest Neolithic settlements in southwest Asia, and the only one currently in the central Zagros region to record occupation spanning the entire agricultural transition. Within the eastern Fertile Crescent only Chogha Golan (ca. 9700–7600 bc), about 170 km away in the lower foothills of the southern Zagros, has provided similar dates (Riehl et al. 2013). Currently, the nearest sites to Sheikh-e Abad from which archaeobotanical remains have been analysed are Ganj Dareh (8240–7840 bc) and Abdul Hosein (8300–7800), both dated to the later phases of the Early Neolithic, when we see the widespread emergence of farming across the region.

a Location of Trenches 1, 2 and 3 at Sheikh-e Abad; b plan of Buildings 1 and 2 in Trench 3; c wild sheep/goat skulls from Building 2, Trench 3
The plant macroremains analysed for this study were collected from deposits across all three trenches. Trench 1 (4.15 m deep) and Trench 2 (2.5 m deep), were both excavated as 2 × 2 m step-trenches and represent stratigraphic sections into the mound. Samples from these areas derive from a range of deposit types, including external midden, possible areas of mudbrick/surface packing, deposits characterised by heavily burnt sequences of ash and charred plant remains. In Trench 2, several deposits with pisé features (clay building material) interpreted as building debris/collapse were also uncovered. In Trench 3 (10 × 10 m plus a 6 × 3 m extension), where open area excavations took place, a considerable expanse of architecture was revealed including a T-shaped mudbrick building (Building 2; Fig. 2b). This single-roomed, arguably ‘symbolic’ building contained several significant features, for example, ground and unworked stone, a fire installation and most strikingly of all in its southeast part, an installation of wild sheep and goat skulls, one bearing traces of red ochre (Fig. 2c). A second building constructed of pisé (Building 1) was recorded to the east of the trench and included an area (Space 8) which is interpreted as an animal pen, based on the identification of a sequence of inter-bedded layers of compacted herbivore dung in micromorphological thin section (Matthews 2013; Shillito et al. 2013). Preliminary analysis of animal bones from the site documents the presence of goat and sheep and to a lesser extent red deer, cattle, fox and hare. Also identified from the faunal assemblage are birds, rodents, tortoise and crab (Bendrey et al. 2013).
Materials and methods
A total of 57 soil samples from 50 contexts, with an average volume of 36.8 litres, were collected from Sheikh-e Abad in 2008, as part of a systematic sampling strategy in which every archaeological unit was sampled. Of these, 41 samples were analysed for this study, corresponding to 1,893 litres of soil. Samples were processed by machine-assisted water flotation with the light fractions (flots) collected in a mesh with an aperture of ca. 250 µm, while the heavy residues were collected in an internal mesh with an aperture of 1 mm. Light fractions were air-dried in the shade and were separated before sorting by passing them through a stack of nested Endecott sieves with aperture sizes of 4, 2, 1 and 0.3 mm. Heavy residues were sorted separately and any plant material that had failed to float was removed and stored alongside its corresponding flot for further sorting and identification.
Analysis of samples was undertaken at the School of Archaeology, University of Oxford, using a Leica EZ4HD and a CETI varizoom stereomicroscope at magnifications of 6× to 40×. Scanning electron microscopy using a Jeol JSM-5910 was used to provide magnifications up to 250,000× where necessary. Identifications were made according to morphological characteristics, surface texture and size, and verified by comparison to modern reference material. Archaeological specimens were compared to both modern uncharred and modern charred material wherever possible, to take account of the potential effects of charring on seed morphology. Plant remains were quantified using the principle of recording the ‘minimum number of individuals’ (MNI) following Jones (1991).
Similarities and differences in the botanical composition of samples were explored within correspondence analysis (CA) (Lange 1990; Jones 1991), which was carried out using the CANOCO for Windows package and the results were plotted using CANODRAW (Šmilauer 1992, 1997). Axis 1 (which explains the most variation) was plotted horizontally and axis 2 vertically. To explore the possibility that wild taxa may have entered the archaeobotanical assemblage within animal dung burnt as a fuel, the bias that ruminant (sheep/goat) digestion imposes on the composition of plant remains surviving within dung was also considered. This has been demonstrated to be closely related to the size and hardness of plant remains (Wallace and Charles 2013). Wild plant taxa were classified as having a low, medium, or high likelihood of surviving in dung, based on (1) their toughness according to the hardness/thickness of their seed coat and whether they are enclosed, and (2) their size in relation to the reticulo-omasal orifice of the ruminant digestive system of sheep, with seeds less than 1.18 mm in all dimensions considered to pass easily through this (Poppi et al. 1985). At present, no attempt has been made to include the complex factor of plant toxicity within our model.
Categorising wild grasses in the archaeological record
In archaeobotanical reports, wild grasses are commonly referred to in terms of non-taxonomic groupings based on the relative size of their seeds, for example ‘large-seeded’ and ‘small-seeded’. However, there is little uniformity in how these categories are applied, making comparisons problematic and leading to a degree of ambiguity in wider discussion. Some scholars have used the term ‘large-seeded’ to refer primarily to the wild progenitor taxa and, despite considerable variation in size, have lumped all remaining taxa together as ‘small-seeded’ (Weiss et al. 2004; Savard et al. 2006). Elsewhere, ‘large-seeded’ has been used to cover a much broader category of wild grasses, including many non-progenitor taxa, such as Taeniatherum caput-medusae and Bromus spp., with the ‘small-seeded’ category reserved for more minute grasses such as Poa spp. (Riehl et al. 2012, 2013; Spengler and Willcox 2013). In some publications a ‘medium-seeded’ category provides an intermediate class that bridges the divide between the large- and small-seeded types (Nesbitt 2006; Riehl et al. 2013). The criteria for these groupings and/or the full list of taxa that fall within them are rarely reported by authors.
Within this paper, we explicitly differentiate between five groups of wild grasses, small-, medium(small)-, medium(medium)-, medium(large)- and large-seeded, based on their relative grain size (Fig. 3). Grain size was calculated from measurements of modern uncharred specimens reported by Nesbitt (2006) in order to avoid potential inconsistencies resulting from the morphological effects of charring (Charles et al. 2015). Grain volume was chosen in preference to length as a comparative measure, due to the latter placing undue emphasis on a single dimension, which can be misleading; for example, Bromus and Stipa grains tend to be long and thin, meaning that length measurements overestimate their relative size (Fig. 3). Ellipsoid volume was selected as the most appropriate volume determination for grass caryopses following Forster et al. (personal communication) and was calculated using the following formula:

The dimensions (mm) and ellipsoid volume (mm3) of wild grasses identified at Sheikh-e Abad along with other common Poaceae (values shown on y-axis). Measurements calculated using values in Nesbitt (2006) as described in text. *Taxa identified at Sheikh-e Abad, †Taxa identified at Sheikh-e Abad that potentially fall into multiple categories (Bromus and Stipa)
where:
While still subjective, the five groupings we propose provide a more nuanced categorisation of wild grasses that avoids giving undue prominence to wild crop progenitors and also recognises the wide range of seed sizes found within the Poaceae family. This final point is of particular significance when considering how seeds were potentially collected, processed and regarded by people in the past. Large-seeded grasses (mainly crop progenitors) still form a distinct group, but other grasses are now divided between four groups, with only the most minute of these (Poa, Crypsis, Puccinellia, Sporobolus) labelled as small-seeded. Grasses whose grain falls between these size divisions are referred to as medium-seeded grasses, within one of three sub-divisions (small, medium and large), with the largest of these also including crop progenitors such as Secale and progenitor-related taxa such as Aegilops.
Results
For the present study, approximately 13,044 charred plant macroremains, representing 130 different taxa/types, were identified across the 41 samples analysed from Trenches 1, 2 and 3 at Sheikh-e Abad (Table 2). Preservation was good and in some cases exceptional. Dominating the assemblage in terms of both frequency and abundance are the wild grasses (67.2% of the assemblage), followed by other wild and weedy taxa (29.4%). Crops, namely Hordeum vulgare (domestic-type barley), Triticum ‘new type’/monococcum/dicoccum (glume wheat) and a range of pulses including Pisum sativum (pea), Lens spp. (lentil) and Vicia ervilia (bitter vetch) represent only a minor component of the plant remains recovered, together accounting for just 2.7% of the total Sheikh-e Abad assemblage (355 out of 13,044 items). Moreover, these are restricted to Trenches 2 and 3 at the site, with no evidence of crops or their wild progenitors recovered from the earlier Trench 1 deposits. Also present at low levels in the assemblage, although recorded in over half of all samples, are the remains of nut/fruit taxa (1.7% of the assemblage), including both Prunus amygdalus (almond) and Pistacia sp. (pistachio).
Within the wild grass category, small-seeded grasses (Poa spp., Crypsis sp. and Sporobolus sp.) are particularly abundant, occurring in 68.3% of samples and accounting for ca. 50% (6,250) of all the macrobotanical items identified from the site. However, there are clear differences in the distribution of small-seeded grasses through time, with the lowest proportions recorded from Trench 1 and the highest from Trench 3, where they dominate every sample analysed (Fig. 4a). Medium-seeded grasses, meaning those within the small, medium and large subdivisions (Fig. 3), are present repeatedly through the site’s occupation but are particularly numerous prior to the establishment of crops (Fig. 4a). Together they account for 25.4% of all the plant remains recovered from Trench 1, 22.8% of plant remains from Trench 2, but only 1.7% of plant remains from Trench 3. Four grasses in particular dominate the assemblage in terms of both ubiquity and abundance: Lolium perenne/rigidum, Piptatherum holciforme, Taeniatherum caput-medusae and Stipa spp. None of these taxa formed part of the original ‘founder suite’ of domesticated cereals that emerged by the 8th millennium bc across the Fertile Crescent.

a Relative percentages of crops, small-seeded grasses, medium-seeded wild grasses (including small, medium and large sub-divisions) and other potentially edible taxa through time at Sheikh-e Abad, based on 24 samples containing 30 or more items. Y-axis gives sample numbers arranged stratigraphically with lowest/oldest at bottom; in brackets the total number of items within each sample. Taxa occurring at proportions < 5% are indicated by +. Presence of rodent droppings in samples indicated by open circles. Samples 510, 617, 613 and 612 potentially missing fine flot (0.3 mm) material and therefore small seeds may be under-represented; b relative percentages of four major medium-seeded grass taxa (Lolium perenne/rigidum, Piptatherum holciforme, Taeniatherum caput-medusae, Stipa spp.) in 24 samples, calculated as percentage of total number of all medium-seeded grass taxa. Y-axis shows sample numbers, arranged stratigraphically with lowest/oldest at bottom; in brackets the total number of medium-seeded grasses within each sample
When comparing the relative proportions of these four taxa over time at the site (Fig. 4b), it is clear that P. holciforme, T. caput-medusae and to a greater extent L. perenne/rigidum all decline through time relative to Stipa spp.; L. perenne/rigidum is absent from Trench 3 deposits altogether. Other medium-seeded grasses, including Bromus spp., are found more frequently from later levels at the site, as is the large-seeded crop progenitor Hordeum spontaneum, which first appears alongside domesticated crops in Trench 2 (Fig. 4a). In addition to grain, fragments of Stipa awn (Fig. 5a) and T. caput-medusae rachis (Fig. 5b) have also been recovered within samples from Trenches 1 and 2, suggesting that these grasses were brought to the site before processing, while Aegilops sp. is represented solely by spikelet bases. Some wild grasses, for example specimens of Stipa, also show clear evidence of having been broken before charring, as demonstrated by the distinctive ‘oozing’ of the endosperm and bulging-smooth appearance of the grain at the broken surfaces (Fig. 5c), a pattern that is consistent with these having been processed for consumption (Willcox 2002; Antolín and Buxó 2011; Valamoti 2011). P. holciforme grains meanwhile have a distinct tapering appearance that was not observed in modern uncharred material (Fig. 5d). Experimental charring undertaken by the authors suggests that this is a result of the grains having been charred in their chaff, plausibly as a result of parching to release the grain, as documented ethnographically for species of the Piptatherum/Oryzopsis complex (Adams 1999).

Examples of major wild grass taxa from Sheikh-e Abad as mentioned in text. a Stipa awn fragment; b Taeniatherum caput-medusae rachis; c Stipa grains showing morphological evidence consistent with having been ground for consumption before charring as discussed in text; d Piptatherum holciforme grains charred in their chaff and demonstrating a distinctive tapering appearance consistent with parching
Other wild taxa identified at Sheikh-e Abad that are significant in terms of their ubiquity and/or abundance include Bolboschoenus glaucus and Chenopodium spp., both of which decline proportionally in later levels of the site (Fig. 4a). In contrast, Brassica/Sinapis sp. and Neslia paniculata as well as Papaver spp. all increase through time (Fig. 4a), a pattern that may relate to their role as arable weeds during later occupation. Figure 6 illustrates the bi-plot for a CA run on all taxa and types occurring in three or more samples (44), after first removing any outliers that obscured underlying trends, and all samples with three or more items of these (14). These three taxa clearly form a group with crops and other wild plants that we interpret as arable weeds (Fig. 6), based on this association as well as their fruiting times which suggest they would have been in seed at the time of harvest (Townsend et al. 1966). Several of these weedy taxa (Bellevalia sp., Papaver spp., Vaccaria pyramidata, Glaucium sp. and Coronilla sp.) also correspond to the potential arable weed taxa listed by Willcox (2012b) for southwest Asian Neolithic assemblages. In contrast, Chenopodium spp. and Bolboschoenus glaucus separate out from crops along the first (horizontal) and second (vertical) axis, suggesting that they did not arrive on site as part of this crop/weed package. This inference is further supported in the case of B. glaucus by its fruiting period, which falls outside of the assumed harvest time (Townsend et al. 1966). Also, diverging from this crop/weed group along the first axis are medium-seeded wild grasses, which form a distinct group in CA unrelated to most other taxa. This suggests that they did not arrive on site as crop weeds or (annual) cultivars, since cultivation would have entailed regular tillage and re-sowing, creating a cultural niche for ruderal plants to invade. However, we do not rule out other forms of crop husbandry such as watering and weeding, especially for perennial plants that may have suited alternative management strategies.

Correspondence analysis plot of 44 taxa based on their occurrences in 14 samples, showing axis 1 (horizontal) and axis 2 (vertical), with taxa coded according to type and labelled as mentioned in text
Several non-botanical remains have also been identified at Sheikh-e Abad, including amorphous charred matter, which requires further investigation, and charred rodent droppings that are restricted to Trench 2 and found in association with crops (Fig. 4a), suggesting that crops or their products were being stored at the site during this time. The remains of potential sheep/goat dung have also been recovered from Trench 2, and were identified in morphological thin sections from Trench 3 together with traces of herbivore and omnivore faecal material identified by gas chromatography mass spectrometry (GC-MS) (Matthews 2013; Shillito et al. 2013).
The burning of animal dung as fuel is a major route by which plant remains can enter a charred macrobotanical assemblage (Miller 1996; Charles 1998; Matthews 2010). At present, there is no unequivocal evidence for any plant remains having a dung origin at Sheikh-e Abad, since no plant macrofossils have been identified within whole dung pellets. However, when considering the potential of seeds for surviving in dung (Table 3), it is possible that many wild plants could have entered the archaeobotanical record in this way. This is particularly the case for those taxa that are unlikely to have arrived as arable weeds based on their seasonality and divergence from the crop/weed group in CA, and for which another explanation is therefore required (e.g. Chenopodium spp. and B. glaucus). Similarly, small-seeded grasses, including Poa spp. which dominate the assemblage, are also ranked as having a high chance of surviving ruminant digestion intact. The dominance of small-seeded grasses from the upper levels of Trench 2 and across Trench 3 (Table 2; Fig. 4a) may relate to an increasing intensity of animal management at the site, corresponding with more widespread trends in the management and emergence of domestic livestock across southwest Asia in the Early Neolithic (Zeder 2011). In contrast, medium- and large-seeded wild grasses are increasingly less likely to have entered the assemblage within animal dung. Furthermore, in the case of specimens that were broken prior to charring such as Stipa grains, and which are therefore reduced in size, it remains unlikely that these came from dung, as despite their endosperm being exposed they are clearly undigested.
Discussion
Identification of wild plant foods
Analysis of the plant remains from Sheikh-e Abad clearly demonstrates that a diverse range of wild taxa was used at the site. This includes a suite of medium-seeded wild grasses that were most likely gathered for consumption. Four taxa, Lolium perenne/rigidum, Piptatherum holciforme, Taeniatherum caput-medusae and Stipa spp., account for the majority of medium-seeded wild grasses recovered, both in terms of ubiquity and abundance. We interpret these grasses as staple foods at Sheikh-e Abad based on the following five lines of independent evidence: (1) their frequent occurrence throughout the assemblage and their presence in significant numbers, for instance 100 or more specimens of a single taxon in a single sample; (2) that they are unlikely to have been crop weeds, given their ripening times in comparison to the harvest period and/or their divergence from crops and crop weeds in CA; (3) their low likelihood of surviving animal digestion intact and having entered the assemblage in animal dung burnt as a fuel; (4) morphological evidence for grains being processed in a manner consistent with their consumption by being ground or parched and (5) the decrease in these taxa (except Stipa spp.) in later levels of occupation following the appearance of domesticated crops.
Other potentially edible wild plant foods found in deposits at Sheikh-e Abad include Chenopodium spp. and Papaver spp., the seeds of which can be used as a condiment, and the nut/fruit group, mainly composed of Pistacia sp. and Prunus amygdalus. Bolboschoenus glaucus may also have been collected for consumption and has been cited as a major food source at other Early Neolithic sites in the region, for example Qermez Dere and Hallan Çemi in southeast Anatolia (Savard et al. 2006). Experimental work on the tubers of B. glaucus has demonstrated how these might have been processed for consumption (Wollstonecroft et al. 2008). Meanwhile, the mustards Brassica/Sinapis sp. and Neslia paniculata could have been used as flavouring and/or for their oil; charred plant assemblages from Neolithic sites in western Asia provide evidence for mustard seeds having been stored and processed for such purposes (Willcox 2002; Bogaard et al. 2009). We use the phrase ‘potentially edible’ here, as unlike the medium-seeded wild grasses discussed, there is no convincing evidence to support the hypothesis that these wild plants were consumed. Indeed, many may have entered the assemblage along other routes, with Papaver spp., N. paniculata and Brassica/Sinapis sp. all noted as potential crop weeds, while Chenopodium spp. and B. glaucus may derive from burnt dung. If this latter inference is supported, it would have significant ramifications for our understanding of fuel use and animal management during the earliest phases of the Neolithic, given current debates regarding the availability of dung fuel during this time and the ubiquity of these taxa in Trench 1 deposits, which date to the 10th millennium bc (Hesse 1984; Miller 1996). We also leave open the possibility that these wild taxa may have arrived on site in myriad ways, have been used for multiple purposes simultaneously (including for medicine or crafts) and that their use(s) could have varied over time. Nor do we exclude the possibility that other taxa found at the site might have been consumed.
Pre-agricultural plant management strategies and ‘auditioning’ of medium-seeded wild grasses
The medium-seeded wild grasses Lolium perenne/rigidum, Piptatherum holciforme, Taeniatherum caput-medusae and Stipa spp. form a significant component of the Sheikh-e Abad assemblage, both in terms of their ubiquity and abundance, and are most reasonably interpreted as processed or staple foods. This repeated and intensive focus on these wild grasses over a prolonged period of time is, we argue, consistent with the concept of ‘auditioning’ (Smith 2007, 2011, 2016), insofar as these taxa appear to have been actively trialled for key roles in food production as part of newly emerging sedentary lifeways during the Early Neolithic. We do not imply by the use of this term that people were trialling plants with any long-term view to, or prior knowledge of, farming, but rather that ‘auditioning’ took place within the broader context of niche construction (sensu Rowley-Conwy and Layton 2001; Smith 2007, 2016) and in pursuit of short-term advantage(s). Critically however, unlike pre-domestication cultivation, this pre-agricultural plant management strategy did not result in the domestication of these taxa and it was eventually supplanted by a strategy of food production based on the cultivation of domesticated crops.
In contrast to other wild grasses whose pre-domestication cultivation is well documented at a number of archaeological sites, for example wild barley (Kislev 1997; Willcox and Stordeur 2012; White and Makarewicz 2012), the absence of a weed flora associated with this group of wild grasses at Sheikh-e Abad suggests that their grain was harvested from wild populations growing in dense, natural stands, as opposed to cultivated plots. Of the four prominent taxa recorded at the site, at least two, P. holciforme and Stipa spp. and potentially also L. perenne/rigidum, are perennials, and thus, while plausibly managed by weeding, watering, protection etc., this would not have provided the same degree of control over the timing and abundance of the harvest as sowing the seeds of annual crops allows (Nesbitt 2006, p 12; van Tassel et al. 2010). It is therefore unlikely that such a strategy would have created the selection pressures necessary for domestication. These medium-seeded wild grasses therefore occupied a fundamentally different niche to that of annual crop progenitors, one that is identified clearly here for the first time in southwest Asia.
It is also possible that a dominance of perennial grasses in the local environment would have made it more difficult for annual crops to become established in the absence of tillage and re-sowing, as they cannot compete with perennial grasses and the dense vegetation that develops; an observation that has been made in modern field experiments on Triticum boeoticum thaoudar (Willcox 1999). This lends further support to the theory that domesticated crops arrived at Sheikh-e Abad as part of an integrated farming ‘package’ that included, by then, established methods of cultivation and crop husbandry. Once adopted, this would have opened up a new agroecological niche, in which perennials as poor and unreliable seed setters would have struggled to compete (Harlan 1992, p 120; Nesbitt 2006, p 13; van Tassel et al. 2010). It is worth noting that there is no evidence to suggest that any attempt was made to integrate these ‘auditioned’ wild grasses within this farming ‘package’ and manage them as annuals; though they overlap in the Trench 2 sequence, their distribution and abundances suggest different routes of entry. Indeed, retaining an alternative strategy of plant management may have helped reduce some of the risks associated with early crop cultivation and food storage, and is consistent with patterns noted across western Asia for the continued use of wild plant foods through the Early Neolithic.
Emergence of farming
Established crops, including domestic-type barley, glume wheat and a range of pulses, first appear in later 8th millennium bc levels of the site. Here, they are associated with a range of wild plant taxa that are best interpreted as weeds of cultivation, including several that are absent from earlier levels, such as Adonis sp. and Ajuga sp. Also, appearing for the first time together with crops are charred rodent droppings, indicating that crops or their products were being stored at the site, although no direct evidence for storage such as containers or bins has been identified. From this evidence, we infer that the later inhabitants of Sheikh-e Abad were engaged in a strategy of food production based on the cultivation of a domesticated crop ‘package’ from at least 7960 cal bc. Significantly, no crop remains (domesticated taxa or their wild progenitors) were recovered from earlier 10th millennium bc levels at the site, suggesting that domesticated crops were introduced to the settlement from an external source, providing a new alternative to existing food procurement strategies. This is in contrast to contemporary Chogha Golan in the low foothills of the southern Zagros mountains, where cultivation of wild crop progenitors such as wild barley, lentil and grass pea is evidenced from the earliest levels of occupation (Riehl et al. 2013; Weide et al. 2017).
Conclusions
Analysis of the Sheikh-e Abad macrobotanical assemblage has allowed us to trace for the first time long-term plant management strategies across the agricultural transition in an upland setting. Significantly, we identify a pre-agricultural strategy of plant food production centred on locally available wild plants, including a group of medium-seeded wild grasses. We observe that several of these wild grasses are perennials, as opposed to the wild progenitors of cereal crops which are all annuals, and hypothesise that the life history of favoured wild grasses, as well as their distribution, was a key factor in diverging plant management trajectories during the Early Neolithic. It is likely that other factors relating to plant availability, seasonality, local climate, animal husbandry and socio-cultural practices at the site also played a role in precluding the in situ development of farming at Sheikh-e Abad. The appearance of domesticated crops in later levels of the site we interpret as the result of a farming ‘package’ being brought in from a distant lowland source. We suggest that the arrival of this farming ‘package’ opened up a new agroecological niche in this upland zone, as cultivation practices that had evolved alongside domesticated annuals allowed these crops to out-compete perennial plants that had previously dominated the local environment. The spread of this new agricultural niche into the central Zagros also corresponds to, and may in part explain, the appearance of other early farming sites here during the later phases of the Early Neolithic, such as Ganj Dareh, Abdul Hosein and Asiab.
Continued sampling and analysis of plant material from the site, including additional radiocarbon dating to refine the chronology of occupation here, will allow us to test these inferences further. It should be stated that the ubiquity values provided in Table 2 have been calculated from a limited number of samples and should be treated with some caution. It should also be noted that while the upper levels of Trench 2 overlap with the lower levels of Trench 3, the top of Trench 1 and base of Trench 2 are separated by a depth of approximately 5 m of deposit, corresponding to some 1,000 years in time (ca. 9000–8000 bc). These deposits remain to be investigated along with other areas of the mound, and are likely to provide additional data on the complexity and diversity of plant management and consumption strategies at the site.
References
Adams KR (1999) Macrobotanical remains. In: Varien MD (ed) The Sand Canyon archaeological project: site testing (Version 1.0, CD-ROM). Crow Canyon Archaeological Center, Cortez
Antolín F, Buxó R (2011) Proposal for the systematic description and taphonomic study of carbonized cereal grain assemblages: a case study of an early Neolithic funerary context in the cave of Can Sadurni (Begues, Barcelona province, Spain). Veget Hist Archaeobot 20:53–66. https://doi.org/10.1007/s00334-010-0255-1
Arranz-Otaegui A, Colledge S, Ibañez JJ, Zapata L (2016a) Crop husbandry activities and wild plant gathering, use and consumption at the EPPNB Tell Qarassa North (south Syria). Veget Hist Archaeobot 25:629–645. https://doi.org/10.1007/s00334-016-0564-0
Arranz-Otaegui A, Colledge S, Zapata L et al (2016b) Regional diversity on the timing for the initial appearance of cereal cultivation and domestication in southwest Asia. Proc Natl Acad Sci 113:14,001–14,006. https://doi.org/10.1073/pnas.1612797113
Badripour H (2006) Country pasture/forage resource profiles. FAO, Islamic Republic of Iran
Bendrey R, Cole G, Tvetmarken CL (2013) Zooarchaeology: preliminary assessment of the animal bones. In: Matthews R, Matthews M, Mohammadifar Y (eds) The earliest Neolithic of Iran: 2008 Excavations at Sheikh-e Abad and Jani. British Institute of Persian Studies and Oxbow Books, Oxford, pp 147–158
Bogaard A, Charles M, Twiss K et al (2009) Private pantries and celebrated surplus: storing and sharing food at Neolithic Çatalhöyük, Central Anatolia. Antiquity 83:649–668
Bor NL (1968) Flora of Iraq, vol 9, Gramineae. Ministry of Agriculture of the Republic of Iraq, Baghdad
Charles M (1998) Fodder from dung: the recognition and interpretation of dung-derived plant material from archaeological sites. Environ Archaeol J Hum Palaeoecol 1:111–122
Charles M, Forster E, Wallace M, Jones G (2015) “Nor ever lightning char thy grain”: establishing archaeologically relevant charring conditions and their effect on glume wheat grain morphology. STAR Sci Technol Archaeol Res 1:1–6. https://doi.org/10.1179/2054892315Y.0000000008
Colledge S (1998) Identifying pre-domestication cultivation using multivariate analysis. In: Damania AB, Valkoun J, Willcox G, Qualset CO (eds) The origins of agriculture and the domestication of crop plants in the Near East. ICARDA, Aleppo, pp 121–131
Colledge S (2001) Plant exploitation on Epipalaeolithic and Early Neolithic sites in the Levant. (British Archaeological Reports, International Series 986). Archaeo Press, Oxford
Colledge S, Conolly J (2014) Wild plant use in European Neolithic subsistence economies: a formal assessment of preservation bias in archaeobotanical assemblages and the implications for understanding changes in plant diet breadth. Quat Sci Rev 101:193–206. https://doi.org/10.1016/j.quascirev.2014.07.013
Cunniff J, Charles M, Jones G, Osborne CP (2010) Was low atmospheric CO(2) a limiting factor in the origin of agriculture? Environ Archaeol 15:113–123. https://doi.org/10.1179/146141010x12640787648469
Edwards PC, Meadows J, Sayej G, Westway M (2004) From the PPNA to the PPNB: new views from the Southern Levant after excavations at Zahrat adh-Dhra’ 2 in Jordan. Paléorient 30:21–60. https://doi.org/10.3406/paleo.2004.1010
Ertug-Yaras F (1997) An ethnoarchaeological study of subsistence and plant gathering in central Anatolia. Doctoral Dissertation, Washington University
Fuller DQ (2007) Contrasting patterns in crop domestication and domestication rates: Recent archaeobotanical insights from the Old World. Ann Bot 100:903–924
Fuller DQ, Willcox G, Allaby RG (2012) Early agricultural pathways: moving outside the “core area” hypothesis in Southwest Asia. J Exp Bot 63:617–633. https://doi.org/10.1093/jxb/err307
Harlan JR (1992) Crops and man, 2nd edn. American Society of Agronomy, Madison
Harlan JR, de Wet JMJ, Price EG (1973) Comparative evolution of cereals. Evolution 27:311–325
Hesse B (1984) These are our goats: the origins of herding in west central Iran. In: Clutton-Brock J, Grigson C (eds) Animals in archaeology, vol 3. Early herders and their flocks. (British Archaeological Reports, International Series 227). Archaeo Press, Oxford, pp 243–262
Hillman GC (2000) Plant food economy of Abu Hureyra: Abu Hureyra 1: The Epipalaeolithic. In: Moore AMT, Hillmann GC, Legge AJ (eds) Village on the Euphrates, from foraging to farming at Abu Hureyra. Oxford University Press, Oxford, pp 327–399
Hopf M (1983) Appendix B: Jericho plant remains. In: Kenyon K, Holland TA (eds) Jericho, vol 5. The pottery phases of the tell and other finds. British School of Archaeology in Jerusalem, London, pp 576–621
Jones G (1991) Numerical analysis in archaeobotany. In: Van Zeist W, Wasylikowa K, Behre K-E (eds) Progress in Old World palaeoethnobotany. Balkema, Rotterdam, pp 63–80
Kehl M (2009) Quaternary climate change in Iran—the state of knowledge. Erdkunde 63:1–17
Kislev ME (1997) Early agriculture and paleoecology of Netiv Hagdud. In: Bar-Yosef O, Gopher A (eds) An early Neolithic village in the Jordan valley. Peabody Museum of Archaeology and Ethnology. Harvard University, Cambridge, pp 209–236
Kuijt I, Finlayson B (2009) Evidence for food storage and predomestication granaries 11,000 years ago in the Jordan Valley. Proc Natl Acad Sci USA 106:10,966–10,970. https://doi.org/10.1073/pnas.0812764106
Kuijt I, Goring-Morris N (2002) Foraging, farming, and social complexity in the Pre-Pottery Neolithic of the Southern Levant: a review and synthesis. J World Prehist 16:361–440
Lange AG (1990) De Horden near Wijk bij Durstede. Plant remains from a native settlement at the Roman frontier: a numerical approach. (Nederlandse Oudheden 13). ROB, Amersfoort
Matthews W (2010) Geoarchaeology and taphonomy of plant remains and microarchaeological residues in early urban environments in the Ancient Near East. Quat Int 214:98–113
Matthews W (2013) Investigating Early Neolithic materials, ecology and sedentism: micromorphology and microstratigraphy. In: Matthews R, Matthews W, Mohammadifar Y (eds) The Earliest Neolithic of Iran: 2008 Excavations at Sheikh-e Abad and Jani. British Institute of Persian Studies and Oxbow Books, Oxford, pp 64–108
Matthews W (2016) Humans and fire: changing relations in early agricultural and built environments in the Zagros, Iran, Iraq. Anthr Rev 3:107–139. https://doi.org/10.1177/2053019616636134
Matthews R, Matthews W, Mohammadifar Y (2013) The earliest Neolithic of Iran: 2008 excavations at Sheikh-e Abad and Jani. British Institute of Persian Studies and Oxbow Books, Oxford
Miller NF (1996) Seed eaters of the ancient Near East: human or herbivore? Curr Anthropol 37:521–528
Morrell PL, Clegg MT (2011) Hordeum, Chap. 6. In: Kole C (ed) Wild crop relatives: genomic and breeding resources. Springer, Berlin, pp 309–319
Nesbitt M (2002) When and where did domesticated cereals first occur in southwest Asia? In: Cappers R, Bottema S (eds) The dawn of farming in the Near East. Ex Oriente, Berlin, pp 113–132
Nesbitt M (2006) Identification guide for Near Eastern grass seeds. Institute of Archaeology, University College London, London
Poppi DP, Hendricksen RE, Minson DJ (1985) The relative resistance to escape of leaf and stem particles from the rumen of cattle and sheep. J Agric Sci 105:9–14. https://doi.org/10.1017/S0021859600055623
Riehl S (2016) The role of the local environment in the slow pace of emerging agriculture in the Fertile Crescent. J Ethnobiol 36:512–534. https://doi.org/10.2993/0278-0771-36.3.512
Riehl S, Benz M, Conard N et al (2012) Plant use in three Pre-Pottery Neolithic sites of the northern and eastern Fertile Crescent: a preliminary report. Veget Hist Archaeobot 21:95–106. https://doi.org/10.1007/s00334-011-0318-y
Riehl S, Zeidi M, Conard NJ (2013) Emergence of agriculture in the foothills of the Zagros Mountains of Iran. Science 341:65–67. https://doi.org/10.1126/science.1236743
Rowley-Conwy P, Layton R (2011) Foraging and farming as niche construction: stable and unstable adaptations. Philos Trans R Soc B 366:849–862
Salamini F, Özkan H, Brandolini A et al (2002) Genetics and geography of wild cereal domestication in the near east. Nat Rev Genet 3:429–441. https://doi.org/10.1038/nrg817
Savard M, Nesbitt M, Jones MK (2006) The role of wild grasses in subsistence and sedentism: new evidence from the northern Fertile Crescent. World Archaeol 38:179–196. https://doi.org/10.1080/00438240600689016
Shillito LM, Matthews W, Bull ID, Williams J (2013) Biomolecular investigations of faecal biomarkers at Sheikh-e Abad and Jani. In: Matthews R, Matthews W, Mohamadifar Y (eds) The earliest Neolithic of Iran: 2008 excavations at Sheikh-e Abad and Jani. British Institute of Persian Studies and Oxbow Books, Oxford, pp 105–116
Šmilauer P (1992) CANODRAW 3.0 user’s guide. Microcomputer Power, New York
Smith BD (2007) Niche construction and the behavioral context of plant and animal domestication. Evol Anthropol 16:188–199
Smith BD (2011) General patterns of niche construction and the management of “wild” plant and animal resources by small-scale pre-industrial societies. Philos Trans R Soc B Biol Sci 366:836–848. https://doi.org/10.1098/rstb.2010.0253
Smith BD (2016) Neo-Darwinism, niche construction theory, and the initial domestication of plants and animals. Evol Ecol 30:307–324. https://doi.org/10.1007/s10682-015-9797-0
Snir A, Nadel D, Groman-Yaroslavski I et al (2015) The origin of cultivation and proto-weeds, long before Neolithic farming. PLoS One 10:e0131422. https://doi.org/10.1371/journal.pone.0131422
Spengler RN, Willcox G (2013) Archaeobotanical results from Sarazm, Tajikistan, an Early Bronze Age settlement on the edge: agriculture and exchange. Environ Archaeol 18:211–221. https://doi.org/10.1179/1749631413Y.0000000008
Stevens LR, Wright HE, Ito E (2001) Proposed changes in seasonality of climate during the Late glacial and Holocene at Lake Zeribar, Iran. Holocene 11:747–755. https://doi.org/10.1191/09596830195762
Ter Braak CFJ, Šmilauer P (1997) CANOCO for Windows v. 4.02. Centre for Biometry, Wageningen
Townsend CC, Guest ER, al-Rawi A (1966) Flora of Iraq, vols 2, 3, 4, 8, 9. Ministry of Agriculture and Agrarian Reform, Baghdad
Valamoti SM (2011) Ground cereal food preparations from Greece: the prehistory and modern survival of traditional Mediterranean “fast foods”. Archaeol Anthropol Sci 3:19–39. https://doi.org/10.1007/s12520-011-0058-z
Van Tassel DL, DeHaan LR, Cox TS (2010) Missing domesticated plant forms: can artificial selection fill the gap? Evol Appl 3:434–452. https://doi.org/10.1111/j.1752-4571.2010.00132.x
Van Zeist W, de Roller GJ (1994) The plant husbandry of aceramic Çayonü, S. E. Turkey. Palaeohistoria 33/34:65–96
Wallace M, Charles M (2013) What goes in does not always come out: the impact of the ruminant digestive system of sheep on plant material, and its importance for the interpretation of dung-derived archaeobotanical assemblages. J Environ Archaeol 18:18–30. https://doi.org/10.1179/1461410313Z.00000000022
Weide A, Riehl S, Zeidi M, Conard NJ (2017) Reconstructing subsistence practices: taphonomic constraints and the interpretation of wild plant remains at aceramic Neolithic Chogha Golan, Iran. Veget Hist Archaeobot 26:487–504. https://doi.org/10.1007/s00334-017-0607-1
Weiss E, Kislev ME, Hartmann A (2006) Autonomous cultivation before domestication. Science 312:1,608–1,610
Weiss E, Kislev ME, Simchoni O et al (2008) Plant-food preparation area on an Upper Paleolithic brush hut floor at Ohalo II, Israel. J Archaeol Sci 35:2,400–2,414
Weiss E, Kislev ME, Simchoni O, Nadel D (2004) Small-grained wild grasses as staple food at the 23,000-year-old site of Ohalo II, Israel. Econ Bot 58:S125–S134
Weiss E, Zohary D (2011) The Neolithic southwest Asian founder crops: their biology and archaeobotany. Curr Anthropol 52:S237-S254. https://doi.org/10.1086/658367
White CE, Makarewicz CA (2012) Harvesting practices and early Neolithic barley cultivation at el-Hemmeh, Jordan. Veget Hist Archaeobot 21:85–94. https://doi.org/10.1007/s00334-011-0309-z
Willcox G (1999) Agrarian change and the beginnings of cultivation in the Near East: evidence from wild progenitors, experimental cultivation and archaeobotanical data. In: Gosden C, Hather J (eds) The prehistory of food: appetites for change. Routledge, London, pp 478–500
Willcox G (2002) Charred plant remains from a 10th millenium bp kitchen at Jerf el Ahmar (Syria). Veget Hist Archaeobot 11:55–60
Willcox G (2005) The distribution, natural habitats and availability of wild cereals in relation to their domestication in the Near East: multiple events, multiple centres. Veget Hist Archaeobot 14:534–541
Willcox G (2012a) The beginnings of cereal cultivation and domestication in Southwest Asia. In: Potts DT (ed) A companion guide to the archaeology of the ancient Near East. Blackwell, Oxford, pp 163–180
Willcox G (2012b) Searching for the origins of arable weeds in the Near East. Veget Hist Archaeobot 21:163–167. https://doi.org/10.1007/s00334-011-0307-1
Willcox G (2013) The roots of cultivation in southwestern Asia. Science 341:39–40. https://doi.org/10.1126/science.1240496
Willcox G, Fornite S, Herveux L (2008) Early Holocene cultivation before domestication in northern Syria. Veget Hist Archaeobot 17:313–325
Willcox G, Stordeur D (2012) Large-scale cereal processing before domestication during the tenth millenium bc cal. in northern Syria. Antiquity 86:99–114
Wollstonecroft MM, Ellis PR, Hillman GC, Fuller DQ (2008) Advances in plant food processing in the Near Eastern Epipalaeolithic and implications for improved edibility and nutrient bioaccessibility: an experimental assessment of Bolboschoenus maritimus (L.) Palla (sea club-rush). Veget Hist Archaeobot 17:S19–S27
Zeder MA (2011) The origins of agriculture in the Near East. Curr Anthropol 52:221–235
Zohary M (1973) Geobotanical foundations of the Middle East, vols 1, 2. Fischer, Stuttgart
Zohary D, Hopf M (2000) Domestication of plants in the Old World: the origin and spread of cultivated plants in West Asia, Europe, and the Nile Valley, 3rd edn. Clarendon Press, Oxford
Acknowledgements
The research reported here was supported by the British Academy (BARDA-48993) and the UK Arts and Humanities Research Council (AH/H034315/1) as part of the Central Zagros Archaeological Project (CZAP), which has been supported by additional grants from the British Institute of Persian Studies, the University of Reading and University College London. We are grateful to our many colleagues in Iran for their support, including Dr. Hashemi and Dr. Mosavi (previously Directors of Research, Iran’s Cultural Heritage, Handicrafts and Tourism Organization—ICHHTO) and our partners at Bu Ali Sina University, Hamedan, as well as the Iranian Center for Archaeological Research (ICAR and its previous Director, Dr. Fazeli Nashli) and ICHHTO for assistance in exporting material for scientific analysis from Iran. Our thanks extend to the entire CZAP team for their work in and out of the field, and to the Institute of Archaeology (University of Oxford) where post-excavation analysis was carried out and where all archaeobotanical material is currently stored.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Fairbairn.
Rights and permissions
About this article

Cite this article
Whitlam, J., Bogaard, A., Matthews, R. et al. Pre-agricultural plant management in the uplands of the central Zagros: the archaeobotanical evidence from Sheikh-e Abad. Veget Hist Archaeobot 27, 817–831 (2018). https://doi.org/10.1007/s00334-018-0675-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-018-0675-x