
Abstract
Forest vegetation composition, including dominant keystone species and floristic diversity, is driven by natural and anthropogenic disturbances. Disentangling these complex interactions to identify the role of species competition, climate and disturbances in boreal forest dynamics is challenging. Here, pollen and charcoal data are used to reconstruct Holocene vegetation and fire history at the local stand-scale within an old-growth Picea abies (Norway spruce) forest hollow in southern Finland. The aim is to reconstruct vegetation history with specific emphasis on the mid-Holocene expansion of Picea and the decline in deciduous species in relation to fire history. Early-Holocene forest development and vegetation dynamics are primarily driven by climatic variations. The spread of Picea at approximately 5,200 cal bp does not coincide with local natural or anthropogenic disturbance or a decline in deciduous species and is consistent with its regional expansion, suggesting climate as the most likely control over the late establishment of this taxon. The mid-Holocene decline in deciduous species corresponds to an increased fire frequency suggesting a higher anthropogenic disturbance (also considered as the primary reason for the loss of floristic diversity in southern Finland). The ‘natural’ fire frequency in this local stand-scale boreal forest is lower than that observed in the recent past (i.e. the time of significant anthropogenic impact), yet the present-day absence or low frequency of fire remains within the range of natural variability observed during the early- and mid-Holocene.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Boreal forest vegetation dynamics are driven by both biotic (e.g. species competition) and abiotic (e.g. climate) processes that are responsible for forest development, composition and structure. Understanding the main factors of vegetation change is important not only for forest conservation and silvicultural management (Birks 1996) but also for assessing, modelling and forecasting future long-term variability and forest structure (e.g. Stocks et al. 1998; Cowling et al. 2001). In addition to natural successional processes, disturbance has an ambiguous yet significant role in driving local vegetation dynamics. Subsequently, forest stands are frequently in a state of disturbance or recovery due to natural (e.g. fire, storms and pests) and anthropogenic (e.g. fire, selective cutting and forest management) drivers often controlling local-scale vegetation dynamics (Bradshaw 1993).
Fire is considered an important disturbance process in the circumboreal forest (e.g. Flannigan et al. 1998; Kasischke and Stocks 2000) and fire regime is driven by a complex interplay between climate, fuel and anthropogenic activity (Whitlock et al. 2010). Separating these drivers is challenging. However, recent attempts to investigate northern European fire regimes suggest that early-Holocene fire is mainly controlled by climate and fuel availability, while mid- to late-Holocene fire is mostly driven by fuel type and land-use change (Molinari et al. 2013; Clear et al. 2014). The significance of mid- to late-Holocene anthropogenic fire disturbance is further evident in dendrological fire scar records (e.g. Zackrisson 1977; Wallenius et al. 2007) which often show very high fire frequencies during periods of significant human impact followed by active fire suppression or a cessation of the use of fire as a land management tool (Wallenius 2011).
The late establishment and mid- to late-Holocene expansion of Picea in an east to west trajectory across Fennoscandia (Giesecke and Bennett 2004; Seppä et al. 2009b) led to a stepwise ecosystem transformation and shift in keystone species. While the cause of this delayed establishment remains uncertain, a shift in climatic condition, specifically continentality (Giesecke et al. 2008), during the early-Holocene, as well as local natural or anthropogenic disturbances (e.g. Molinari et al. 2005; Bjune et al. 2009) appear to have facilitated the spread of Picea. A change in fire frequency; from a fire prone pine-dominated forest to a fire free spruce-dominated environment is concomitant with the spread of Picea (Ohlson et al. 2011; Clear et al. 2014). This reduced fire frequency creates a positive feedback that, through lack of natural disturbance, further encourages the dominance of Picea (Clear et al. 2013; Kuosmanen et al. 2014). This in turn causes a decline in deciduous species and subsequent loss of floristic and faunal diversity (Lindbladh et al. 2003) and creates a Picea-dominant forest that becomes more susceptible to natural disturbance e.g. wind throw and spruce bark beetle (Ips typographus) disturbance (e.g. Kärvemo and Schroeder 2010; Griess and Knoke 2011). As a consequence prescribed burning is advocated to induce disturbance-related regeneration to enhance floristic and faunal diversity (Hyvärinen et al. 2009; Halme et al. 2013).
The observed decline in temperate deciduous species with a consequent decrease of floristic diversity during the mid- to late-Holocene in Fennoscandia is commonly attributed to an interaction between climate (e.g. Greisman and Gaillard 2009), species competition, in particularly the spread of Picea (e.g. Seppä et al. 2009b), and increased or intensified anthropogenic disturbance (e.g. Lindbladh et al. 2000; Molinari et al. 2005).
Observing Holocene vegetation and disturbance dynamics using palaeo-proxies (i.e. pollen and charcoal) allows for the reconstruction of forest succession and development (including changing keystone species and stand diversity) and fire history (e.g. Bradshaw and Hannon 1992; Giesecke 2005; Alenius et al. 2008; Ohlson et al. 2011), as well as the identification of the potential causes and drivers behind the observed vegetational change.
Stand-scale palynology (Jacobson and Bradshaw 1981) or the analysis of proxies from sediment deposited in forest hollows, are excellent methods for recording local-scale environmental change and disturbance dynamics. Pollen source area is limited, with half of the pollen derived from the local source area (up to 100 m) and the remaining pollen originating from the regional pollen sources (Andersen 1970; Sugita 1994). Further, macroscopic charcoal fragments (>300 µm) can reconstruct fire activity with a high spatial precision (Ohlson and Tryterud 2000; Ohlson et al. 2006).
The aim of this paper is to explore the expansion of Picea abies and the mid-Holocene decline in deciduous species and floristic diversity within a forest hollow in southern Finland and in particular, establish their relationships to local fire regime. We hypothesise that changes in Holocene fire regime are the major drivers of stand dynamics.
Materials and methods
Study area and sites
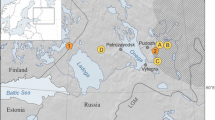
Sudenpesä (61°11′N; 25°09′E), a stand-scale site approximately 15 m2 in area, was selected for its forest hollow characteristics (Jacobson and Bradshaw 1981; Overballe-Petersen and Bradshaw 2011). Sudenpesä is situated in the southern boreal vegetation zone (Ahti et al. 1968) of southern Finland (Fig. 1) in the Evo Research Forest, within the old-growth forest of Sudenpesänkangas. The mean annual temperature is 4.2 °C with a July mean of 16.6 °C and a February mean temperature of −7.1 °C (Pirinen et al. 2012). The mean annual precipitation is 645 mm. The regional bedrock consists of orogenic granitoids covered with till (Palviainen et al. 2009) and the site is composed of nutrient-rich drained peatland (Kaila et al. 2012). The regional vegetation type is dominated by boreal Picea and Pinus mixed with Betula, while at present, Sudenpesä forest is dominated by Picea.

Location map of Sudenpesä forest hollow in southern Finland
Field and laboratory methods
A sediment core was extracted from the centre of the Sudenpesä forest hollow in May 2008 using a 50 cm long, 5 cm diameter Russian corer (Jowsey 1966). Sediment extraction comprised three core sections taken from adjacent holes from the surface down with core drives alternated between two core holes to enable a 10 cm overlap. A total of 145 cm of sediment was extracted from the Sudenpesä forest hollow. Cores were secured in drain piping before being transported to the University of Liverpool and stored below 5 °C prior to sub-sampling. The Sudenpesä sediment was sub-sampled at 0.5 cm intervals between 145 and 60 cm and at 1 cm intervals between 60 and 35 cm. Above 35 cm the sediment was poorly compacted, consisting of loose Sphagnum, and attempts to retain sediment for analysis failed.
Pollen and macroscopic charcoal analysis
Pollen and macroscopic charcoal were analysed from continuous sub-samples. Pollen preparation followed standard procedures (Moore et al. 1991) with a minimum of 500 terrestrial pollen grains counted per sample and, where possible, identified to species level following Moore et al. (1991) and Bennett (1995–2007). The percentage pollen diagram (Fig. 3) was created using TILIA and TILIA.GRAPH 2011 version 1.5.12 (Grimm 2011). A stratigraphically-constrained cluster analysis was applied in CONISS on square root transformed data for selected abundant terrestrial pollen taxa (maximum frequency >1 %, Grimm 1987).
Macroscopic charcoal preparation involved a known volume of sediment treated with dilute NaOH to assist with sediment disaggregation before being sieved at 300 µm. The sediment residue retained was added to 80 ml of double distilled water. 20 ml of this suspension was transferred to a petri dish for analysis. Black brittle crystalline particles with angular broken ends were classified as charcoal (Swain 1973, 1978). Charcoal concentrations (particles cm−3) and charcoal influx (particles cm−2 year−1) were calculated and graphically displayed using SigmaPlot version 12.3. Fire frequency estimates were derived from the charcoal record by assuming that absence of charcoal over a period separated individual fire events. As deposition of charcoal is often distributed over several decades (Higuera et al. 2005), assuming consistent depositional characteristics, the second sample to record consecutive charcoal was taken as the fire year to calculate fire frequency as described in Clear et al. (2013). The absence of charcoal from samples in a forest hollow is not uncommon and is a reflection of the local dispersion and deposition characteristics, Because many of the samples are free from charcoal fragments, it is assumed that the background charcoal signal remains low during times of high charcoal influx and that these peaks in charcoal record primarily local fire events.
Radiocarbon dating
A total of 9 sub-samples were selected for AMS 14C dating (Table 1). The sediment exterior was scraped to remove possible contaminants and the remaining fraction of sediment between 0.5 and 2 g was dated. 14C measurements were calibrated and the age-depth calibration curve was calculated using the Bayesian accumulation model, Bacon (Blaauw and Christen 2011; Fig. 2).

Bayesian accumulation age-depth model (BACON) output for Sudenpesä based on 9 AMS 14C dates. The BACON model can identify date inversions and exclude the outlier from the age-depth model
Statistical analysis
Compositional turnover (Birks 2007; Bjune et al. 2009) was estimated using detrended canonical correspondence analysis (DCCA) in CANOCO 4.5 (Ter Braak and Šmilauer 2002). Pollen data (species) were square-root transformed to stabilise variance and age (cal bp) was applied as the sole constraining variable. Palynological richness (Birks and Line 1992) was estimated using rarefaction analysis in R (R Development Core Team 2010). As the total pollen count varies between samples, estimates of the number of expected taxa [E(Tn)] were calculated using the lowest total pollen count as the baseline for comparison of palynological richness amongst samples. Pollen assemblages do not directly reflect vegetation diversity as there is a nonlinear relationship between pollen-taxa and plant richness (Odgaard 1999). Palynological richness is used here as an estimation of floristic diversity and not as an accurate representation of vegetation communities (e.g. Goring et al. 2013; Meltsov et al. 2013).
Results
Chronology
The Bayesian derived age-depth curve for Sudenpesä indicates that the sediment core extends back over 9,500 cal year bp. Initial sediment accumulation is slow before entering an extended phase of relatively constant sedimentation (Fig. 2) that continues until the present day. Radiocarbon analysis returned results of a modern age from 35 to 36 cm depth. The top 35 cm of Sudenpesä consisted of unconsolidated, wet Sphagnum with little sediment available and yielded a poor record of pollen and no evidence of charcoal. There is uncertainty associated with the radiocarbon result recorded at 110–111 cm with a 14C date of 1,575 ± 30 bp (Table 1), which appears too young to fit linearly with the age-depth curve.
Vegetation and disturbance history
The CONISS analysis identified four pollen accumulation zones (PAZ) defined as SUD 1–4. PAZ SUD-1 (9,600–7,600 cal bp) is representative of the early Holocene and is dominated by a Pinus–Betula forest stand with a high abundance of mixed-deciduous trees, shrubs and herbs. PAZ SUD-2 (7,600–3,100 cal bp) represents the mid-Holocene and is characterised by a co-dominant Pinus–Betula forest stand with an increased abundance of Alnus and a high abundance of mixed-deciduous trees, shrubs and herbs. This zone is further defined by the onset of the almost continuous curve of Picea and the mid-Holocene rise in Picea abundance. PAZ SUD-3 (3,100–1,700 cal bp) extends from the mid- to late-Holocene and is defined by a mixed coniferous (Picea and Pinus), deciduous (Alnus and Betula) forest stand with declining abundance in vegetation diversity, PAZ SUD-4 (1,700-present day) is representative of the late Holocene with a variable abundance of mixed coniferous (Picea and Pinus) and deciduous (Alnus and Betula) species with an increase in Poaceae.
PAZ SUD-1 (9,600–7,600 cal bp) Early Holocene Pinus–Betula forest stand
Early Holocene vegetation development at Sudenpesä consists of a co-dominant Pinus–Betula forest stand (Fig. 3). An initial high presence of Betula coincides with the occurrence of Corylus, Populus, Ericaceae and Cyperaceae. A subsequent rise in Pinus occurs with an increase in Poaceae. Compositional turnover (Fig. 4a) is initially high and declines once Pinus abundance increases. No charcoal is recorded between 9,600 and 8,700 cal bp indicating an initial absence of fire. Between 8,700 and 7,800 cal bp charcoal presence indicates a regular fire occurrence with an average fire return interval of approximately 230 years (Fig. 5), associated with an increase in both Betula and Alnus and a decline in Pinus as well in compositional turnover and palynological richness (Fig. 4a, b).


Pollen and charcoal diagram for Sudenpesä with pollen expressed as a percentage of total pollen count and macroscopic charcoal fragments (>300 µm) expressed as concentration values (particles cm−3)

a Compositional turnover (standard deviation units), b palynological richness (estimated number of taxa) and c macroscopic charcoal (particles cm−3) for Sudenpesä forest hollow

Charcoal influx (macroscopic particles cm−2 year−1) and selected pollen taxa with fire frequency estimation
PAZ SUD-2 (7,600–3,100 cal bp) Mid-Holocene increase in Picea abies
PAZ SUD-2 is defined by a diverse mixed coniferous-deciduous forest stand co-dominated by Betula, Pinus and Alnus with an underlying presence of Corylus, Juniperus, Populus, Salix, Tilia, Ulmus, Cyperaceae and Poaceae (Fig. 3). The onset of the almost continuous curve of Picea begins around 7,600 cal bp. The period between 7,600 and 700 cal bp is characterised by the absence of charcoal, suggesting a period without fire. Between 7,000 and 5,100 cal bp regular charcoal occurrence indicates a fire return interval of approximately 330 years (Fig. 5) and coincides with high floristic diversity (Fig. 3), steady compositional turnover (Fig. 4a) and variable palynological richness driven by peaks in charcoal abundance (Fig. 4b).
The expansion of the continuous curve of Picea and subsequent increase in Poaceae begin at approximately 5,200 cal bp (Fig. 3). The rise in Picea corresponds to a period of frequent charcoal occurrence with a fire frequency calculation of approximately 150 year intervals between 5,100 and 4,500 cal bp (Fig. 5). This suggests an increase in fire activity associated with the local expansion of Picea. Between 4,500 and 3,500 cal bp charcoal abundance declines, with a fire frequency of approximately 510 year intervals which is lower than any previous background level of fire activity. During this period rarefaction results record variable levels of palynological diversity (Fig. 4a) and relatively high levels of compositional turnover (Fig. 4b) driven by fluctuating pollen abundance (Fig. 3). Species diversity remains high until approximately 3,500 cal bp when charcoal abundance indicates a shift in fire frequency to approximately 200 year intervals (Fig. 5).
PAZ SUD-3 (3,100–1,700 cal bp) mid- to late Holocene decline in floristic diversity
The mid- to late Holocene PAZ SUD-3 is defined by a decrease in floristic diversity, underlined by a decline in the Quercus, Salix, Tilia and Ulmus pollen curves (Fig. 3) and recorded as a declining trend in the palynological richness (Fig. 4b). Compositional turnover remains relatively high (Fig. 4a) and corresponds to peaks in charcoal abundance (Fig. 4c) which indicate a regular fire frequency of approximately 200 year intervals (Fig. 5).
PAZ SUD-4 (1,700 cal bp to present day) late-Holocene boreal forest ecosystem
The late-Holocene PAZ SUD-4 is dominated by a mixed coniferous (Picea and Pinus) and deciduous (Alnus and Betula) forest stand with a high abundance of Poaceae (Fig. 3) which appears to drive compositional turnover (Fig. 4a). Palynological richness (Fig. 4b) is relatively low. Charcoal abundance indicates a fire frequency of at approximately 200 year intervals until 400 cal bp while later the absence of charcoal suggests a lack of recent fire (Fig. 5).
Discussion
Climate-driven natural forest succession
During the early Holocene, the retreating margin of the Weichselian ice sheet reached southern Finland approximately 13,000 cal bp (Lunkka et al. 2004). The drop in the Baltic Ice Lake level and land isostatic adjustment meant that the region was terrestrial by approximately 11,000 cal bp (Johansson et al. 2011). This enabled a succession of post-glacial vegetation development, initially with scattered Betula stands and then with Pinus–Betula forests (Björck and Möller 1987). There is no evidence of fire documented in Sudenpesä prior to 8,800 cal bp. Natural fire requires both availability of fuel (biomass) and favourable climate conditions and, although rapid climate warming during the early-Holocene led to the deglaciation of the Weichselian ice sheet (Lundqvist 1986), climate reconstructions indicate low annual mean temperatures (Heikkilä and Seppä 2003) and high precipitation levels (Korhola 1995). It is only once the mixed coniferous-deciduous forest becomes fully established that fuel availability is substantial enough for the forest to burn. Between 8,800 and 7,700 cal bp there is a rise in fire activity with a fire frequency of approximately 230 year intervals, most probably associated with an increase in fuel load and prevailing cool, dry climate conditions (Seppä et al. 2009a; Zillén and Snowball 2009). The increase in Betula and the decline in Pinus observed during this period in Sudenpesä is also registered in other palaeoecological records at the local (Bjune et al. 2009) and the regional scale (Giesecke 2005; Alenius et al. 2013), suggesting climate as the controlling factor of vegetation changes in the early Holocene (Huntley 1990).
The local establishment of Picea abies
The onset of Picea in Sudenpesä begins at approximately 7,600 cal bp. This coincides with an absence of charcoal suggesting a period without fire between 7,600 and 7,000 cal bp. However, prior to the low-level establishment of Picea, fires occurred approximately every 230 years. The stand-scale establishment of Picea has frequently been linked to local disturbance, most commonly in south-western Scandinavia where the increase of this species often coincides with an increase in anthropogenic disturbance (e.g. Molinari et al. 2005; Bjune et al. 2009). The absence of fire, once Picea becomes established, is also evident elsewhere throughout Fennoscandia where Picea has the ability to alter the fire regime from a fire prone to a predominantly fire free environment (e.g. Tallantire 1972; Ohlson et al. 2011; Clear et al. 2014). However, distinguishing between whether or not the absence or low frequency of fire is a driver or consequence of an increased abundance of Picea remains challenging. In Sudenpesä during its early establishment, the abundance of Picea is negligible suggesting that this could not have been the single cause for the altered local fire regime. The observed reduction in fire may be a causal factor of Picea establishment rather than the result of its increased abundance, indicating that altered climate is the most likely cause for the decreasing local fire frequency (e.g. Carcaillet et al. 2007; Drobyshev et al. 2012) during the early to mid-Holocene in Sudenpesä.
The early continuous curve or tail associated with Picea establishment from approximately 7,600 cal bp is not uncommon and is recorded at local and regional sites throughout Fennoscandia (see Giesecke and Bennett (2004) and references therein). However, the question remains; why does Picea not thrive immediately after becoming locally established? The lack of fire at Sudenpesä between 7,600 and 7,000 cal bp could have reduced the potential for Picea to regenerate during the early-Holocene. However when fire does return to Sudenpesä, at approximately 7,000 cal bp. Picea still does not increase its population size suggesting that factors other than local scale disturbance act as a control over the rise to dominance of Picea at the stand-scale in southern Finland. Picea is generally a continental tree species with high summer temperature requirements and maximum mean coldest month temperature requirements of −1.5 °C (Giesecke 2004). Precipitation has a strong effect on distribution at low latitudes and temperature is important at higher latitudes and altitudes (Giesecke 2004). The increase in Picea abundance in Sudenpesä between approximately 5,500 and 5,000 cal bp is observed in other southern Finland pollen records (e.g. Sarmaja-Korjonen 1998; Jauhiainen et al. 2004; Clear et al. 2013). Then the species spreads in a wave-like distribution across Fennoscandia (Giesecke and Bennett 2004). An independent climate reconstruction using δ18O from lake Saarikko (Heikkilä et al. 2010) suggests summer temperatures reached a maximum between 6,000 and 5,000 cal bp indicating that climate was the limiting factor for Picea establishment in the early to mid-Holocene. This is further justified by the low abundance of Picea recorded from approximately 7,600 cal bp. This low presence of Picea during the early Holocene is also observed in other pollen records from southern and northern Finland (e.g. Vasari 1962; Tolonen 1967, 1983; Tolonen 1980). The theory that continentality controls Picea abundance also fits with the Picea macrofossils found in the Swedish Scandes Mountains (Kullman 2000, 2001) where outposts of Picea population were present during the early post-glacial period approximately 10,000 cal bp. Further, their ability to thrive locally in the Scandes Mountains between 6,000 and 5,000 cal bp was controlled by a changing climate, most notably increased continentality (Giesecke and Bennett 2004). It is during this period that climatic conditions became optimal to drive the spread of Picea at a maximum rate of 250 m year−1 (Bialozyt et al. 2012) in an east–west trajectory across Fennoscandia (Tallantire 1972; Giesecke and Bennett 2004; Seppä et al. 2009b). Picea may still be spreading in southern Sweden and Norway where it is near its ecological limits (Bradshaw 2007), however the spread of Picea in southern Sweden is partly anthropogenic driven, where it is currently planted beyond its climatic range limits (Bradshaw et al. 2000).
In Sudenpesä the local rise in the abundance of Picea between 5,100–4,500 cal bp is associated with increased fire activity with a fire frequency of approximately 150 years. This association is often interpreted as fire disturbance facilitating the local establishment of Picea (e.g. Molinari et al. 2005). However, in Sudenpesä the increase of fire occurs after the initial increase in Picea with fire frequency declining once Picea becomes fully established. Thus, this is associated with a change in fuel type i.e. an increase in spruce saplings and their high ability to burn (Tanskanen et al. 2006). It is interesting to note that once Picea becomes established the fire frequency shifts to the lowest value recorded in Sudenpesä (approximately 510 years). It is well documented that Picea has the ability to alter the local microclimate reducing fire ignition potential (Tanskanen et al. 2005) and thus diversifying the natural fire regime (Ohlson et al. 2011). This is observed in western Scandinavia where the spread of Picea coincided with heightened anthropogenic activity around 2,000 cal bp (Bradshaw and Hannon 1992; Lindbladh et al. 2000). However, in southern Finland the fire frequency prior to Picea establishment was already naturally low, suggesting that fire was not a primary factor in the local establishment of Picea.
Floristic diversity and disturbance
The mid-Holocene deciduous abundance and diversity observed in pollen records throughout Fennoscandia (e.g. Tolonen 1980; Giesecke 2005; Molinari et al. 2005) are concomitant with the mid-Holocene Thermal Maximum (HTM), when temperatures were between 1.5 and 3 °C warmer than today (Seppä and Birks 2001; Heikkilä and Seppä 2003; Bjune et al. 2004; Antonsson and Seppä 2007; Bjune and Birks 2008). It is frequently noted that the decline in deciduous abundance is mainly a product of the mid-Holocene climate cooling and that modern pollen-climate calibration data sets can be combined with fossil pollen data to produce Holocene climate reconstructions through pollen–climate transfer functions (e.g. Seppä et al. 2009a; Bjune et al. 2010). One further explanation for the mid-Holocene decline in deciduous trees (Tilia in particular) is species competition driven by the late expansion of Picea (Seppä et al. 2009a).
The decline in deciduous species abundance and subsequent loss in palynological richness observed in Sudenpesä begins at approximately 3,500 cal bp. This does not coincide with an increase in Picea suggesting species competition was an insignificant factor at this site. The observed decline in Sudenpesä does coincide with a shift from a relatively low fire frequency of 510 years to a more frequent fire interval of approximately 200 years. This change in fire frequency could be associated with variation in climate conditions (e.g. Carcaillet et al. 2007; Drobyshev et al. 2012). However, if we compare Sudenpesä to published fire frequency records with established chronologies in southern Finland (e.g. Jauhiainen et al. 2004; Clear et al. 2013), it becomes clear that, independently of climate, increased fire activity, probably driven by anthropogenic activity, contributes to the decline in deciduous species. This is further supported by archaeological evidence in the region where by 3,000 cal bp swidden or slash and burn culture thrived in southern Finland (Sarmela 1987) and focused on the clearance of broadleaf trees (Parviainen 1996). The establishment of permanent agriculture led to population expansion and anthropogenic disturbance became the strongest driver of forest compositional change (Reitalu et al. 2013). This has further been observed elsewhere in Fennoscandia (e.g. Molinari et al. 2005), where an increase in human use of fire appears to drive the mid-Holocene decline in deciduous species and loss of floristic diversity. It can therefore be proposed that the mid-Holocene decline in deciduous species in southern Finland is primarily driven by intensified anthropogenic activity including an increase in the human use of fire and selective slash and burn practices that focus on the clearance of deciduous species. Without this intensified anthropogenic activity, the floristic diversity may have remained until the present day at suitable locations. While there may also be a regional decline in deciduous tree species associated with late Holocene climatic change, anthropogenic disturbance is likely to have been a significant factor. This is also observed in southern Sweden (Björse and Bradshaw 1998).
Conclusion
The shifting drivers of vegetation dynamics are presented for almost 10,000 years of forest compositional change at a local site in southern Finland. Early-Holocene post-glacial vegetation development and compositional change is primarily driven by climate variability. The mid-Holocene expansion of Picea abies occurred independently of local disturbance and did not initiate a decline in deciduous species or alter floristic diversity. The almost continuous curve of low abundance of Picea pollen from approximately 7,600 cal bp suggests a regional presence of Picea with local, independent climate records supporting a shift in continentality that favoured spruce expansion from 5,200 cal bp, leading to its further regional spread. The observed decline in deciduous species from approximately 3,500 cal bp could be climatically driven at Sudenpesä, but is associated with a shift in fire frequency indicative of increased human impact and selective slash and burn cultivation. This suggests that the local decline in deciduous species is at least partly controlled by anthropogenic disturbance. The present day fire frequency observed in southern Finland is low relative to reconstructions of fire frequency in the recent past (i.e. the time of significant human disturbance), but is within the Holocene natural range of variability when observing fire frequencies during the early to mid-Holocene. Our initial hypothesis is partially upheld as changes in fire regime, driven by both climatic and anthropogenic factors, are closely associated with major dynamic episodes such as the loss of deciduous species and the rise to local dominance of Picea. However ecosystem modelling must be combined with palaeoecological observations to further disentangle the major drivers of vegetation change.
References
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in northwestern Europe. Ann Bot Fenn 5:169–211
Alenius T, Mikkola E, Ojala A (2008) History of agriculture in Mikkeli Orijärvi, eastern Finland as reflected by palynological and archaeological data. Veget Hist Archaeobot 17:171–183
Alenius T, Mökkönen T, Lahelma A (2013) Early farming in the northern Boreal zone: reassessing the history of landuse in southeastern Finland through high-resolution pollen analysis. Geoarchaeology 28:1–24
Andersen ST (1970) The relative pollen productivity and pollen representation of north European trees, and correction factors for tree pollen spectra. Dan Geol Unders II 96:1–99
Antonsson K, Seppä H (2007) Holocene temperatures in Bohuslan, southwest Sweden: a quantitative reconstruction from fossil pollen data. Boreas 36:400–410
Bennett KD (1995–2007) Pollen catalogue of the British Isles. http://chrono.qub.ac.uk/pollen/pc-intro.html. Accessed 2008–2013
Bialozyt R, Bradley LR, Bradshaw RHW (2012) Modelling the spread of Fagus sylvatica and Picea abies in southern Scandinavia during the late Holocene. J Biogeogr 39:665–675
Birks HJB (1996) Contributions of Quaternary palaeoecology to nature conservation. J Veget Sci 7:89–98
Birks HJB (2007) Estimating the amount of compositional change in late-Quaternary pollen-stratigraphical data. Veget Hist Archaeobot 16:197–202
Birks HJB, Line JM (1992) The use of rarefaction analysis for estimating palynological richness from quaternary pollen-analytical data. Holocene 2:1–10
Björck S, Möller P (1987) Late Weichselian environmental history in southeastern Sweden during the deglaciation of the Scandinavian ice sheet. Quat Res 28:1–37
Björse G, Bradshaw R (1998) 2000 years of forest dynamics in southern Sweden: suggestions for forest management. For Ecol Manag 104:15–26
Bjune AE, Birks HJB (2008) Holocene vegetation dynamics and inferred climate changes at Svanåvatnet, Mo i Rana, northern Norway. Boreas 37:146–156
Bjune AE, Birks HJB, Seppä H (2004) Holocene vegetation and climate history on a continental–oceanic transect in northern Fennoscandia based on pollen and plant macrofossils. Boreas 33:211–223
Bjune AE, Ohlson M, Birks HJB, Bradshaw RHW (2009) The development and local stand-scale dynamics of a Picea abies forest in southeastern Norway. Holocene 19:1,073–1,082
Bjune A, Seppä H, Birks H (2010) Quantitative summer-temperature reconstructions for the last 2000 years based on pollen-stratigraphical data from northern Fennoscandia. J Paleolimnol 41:43–56
Blaauw M, Christen JA (2011) Flexible palaeoclimate age-depth models using an autoregressive gamma process. Bayesian Anal 6:457–474
Bradshaw RHW (1993) Tree species dynamics and disturbance in three Swedish boreal forest stands during the last 2000 years. J Veget Sci 4:759–764
Bradshaw RHW (2007) Stand-scale palynology. In: Elias SA (ed) Encyclopaedia of Quaternary Science. Elsevier, Amsterdam, pp 2,535–2,543
Bradshaw RHW, Hannon GE (1992) Climatic change, human influence and disturbance regime in the control of vegetation dynamics within Fiby forest, Sweden. J Ecol 80:625–632
Bradshaw RHW, Holmqvist BH, Cowling SA, Sykes MT (2000) The effects of climate change on the distribution and management of Picea abies in southern Scandinavia. Can J For Res 30:1,992–1,998
Carcaillet C, Bergman I, Delorme S, Hornberg G, Zackrisson O (2007) Long-term fire frequency not linked to prehistoric occupation in Northern Swedish Boreal Forest. Ecol 88:465–477
Clear JL, Seppä H, Kuosmanen N, Bradshaw RHW (2013) Holocene fire frequency variability in Vesijako, Strict Nature Reserve, Finland, and its application to conservation and management. Biol Conserv 166:90–97
Clear JL, Molinari C, Bradshaw RHW (2014) Holocene fire in Fennoscandia and Denmark. Int J Wild Fire 23:781–789
Cowling SA, Sykes MT, Bradshaw RHW (2001) Palaeovegetation-model comparisons, climate change and tree succession in Scandinavia over the past 1500 years. J Ecol 89:227–236
Development Core Team R (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Drobyshev I, Niklasson M, Linderholm HW (2012) Forest fire activity in Sweden: climate controls and geographical patterns in 20th century. Agric Forest Meteorol 154–155:174–186
Flannigan MD, Bergeron Y, Engelmark O, Wotton BM (1998) Future wildfire in Circumboreal forests in relation to global warming. J Veget Sci 9:469–476
Giesecke T (2004) The Holocene Spread of Spruce in Scandinavia. Acta Universitatis Upsaliensis. Comprehensive summaries of Uppsala Dissertations from the Faculty of Science and Technology 1027, Uppsala
Giesecke T (2005) Holocene forest development in the central Scandes Mountains, Sweden. Veget Hist Archaeobot 14:133–147
Giesecke T, Bennett KD (2004) The Holocene spread of Picea abies (L.) Karst. in Fennoscandia and adjacent areas. J Biogeogr 31:1,523–1,548
Giesecke T, Bjune AE, Chiverrell RC, Seppä H, Ojala AEK, Birks HJB (2008) Exploring Holocene continentality changes in Fennoscandia using present and past tree distributions. Quat Sci Rev 27:1,296–1,308
Goring S, Lacourse T, Pellatt MG, Mathewes RW (2013) Pollen assemblage richness does not reflect regional plant species richness: a cautionary tale. J Ecol 101:1,137–1,145
Greisman A, Gaillard M-J (2009) The role of climate variability and fire in early and mid Holocene forest dynamics of southern Sweden. J Quat Sci 24:593–611
Griess VC, Knoke T (2011) Growth performance, windthrow and insects: meta-analyses of parameters influencing performance of mixed-species stands in boreal and northern temperate biomes. Can J For Res 41:1,141–1,159
Grimm EC (1987) CONISS: a FORTRAN 77 Program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput Geosci 13:13–35
Grimm EC (2011) TILIA and TILIA.GRAPH. Springfield, Illinois State Museum
Halme P, Allen KA, Auniņš A et al (2013) Challenges of ecological restoration: lessons from forests in northern Europe. Biol Conserv 167:248–256
Heikkilä M, Seppä H (2003) A 11000 year palaeotemperature reconstruction from the southern boreal zone in Finland. Quat Sci Rev 22:541–554
Heikkilä M, Edwards TWD, Seppä H, Sonninen E (2010) Sediment isotope tracers from Lake Saarikko, Finland, and implications for Holocene hydroclimatology. Quat Sci Rev 29:2,146–2,160
Higuera PE, Sprugel DG, Brubaker LB (2005) Reconstructing fire regimes with charcoal from small-hollow sediments: a calibration with tree-ring records of fire. Holocene 15:238–251
Huntley B (1990) European post-glacial forests: compositional changes in response to climatic change. J Veget Sci 1:507–518
Hyvärinen E, Kouki J, Martikainen P (2009) Prescribed fires and retention trees help to conserve beetle diversity in managed Boreal forests despite their transient negative effects on some beetle groups. Insect Conserv Divers 2:93–105
Jacobson GL, Bradshaw RHW (1981) The selection of sites for paleovegetational studies. Quat Res 16:80–96
Jauhiainen S, Pitkänen A, Vasander H (2004) Chemostratigraphy and vegetation in two boreal mires during the Holocene. Holocene 14:769–778
Johansson P, Lunkka JP, Sarala P (2011) The glaciation of Finland. In: Ehlers J, Gibbard PL, Hughes PD (eds) Quaternary glaciations: extent and chronology: a closer look (developments in quaternary science 15). Elsevier, Amsterdam, pp 105–116
Jowsey PC (1966) An improved peat sampler. New Phyt 65:245–248
Kaila A, Asam ZUZ, Sarkkola S, Xiao L, Laurén A, Vasander H, Nieminen M (2012) Decomposition of harvest residue needles on peatlands drained for forestry: Implications for nutrient and heavy metal dynamics. For Ecol Manag 277:141–149
Kärvemo S, Schroeder LM (2010) A comparison of outbreak dynamics of the spruce bark beetle in Sweden and the mountain pine beetle in Canada (Curculionidae: Scolytinae). Entomol Tidskr 131:215–224
Kasischke ES, Stocks BJ (2000) Fire, climate change and carbon cycling in the Boreal forest. Ecol Studies 138. Springer, New York
Korhola A (1995) Holocene climatic variations in southern Finland reconstructed from peat-initiation data. Holocene 5:43–57
Kullman L (2000) The geoecological history of Picea abies in northern Sweden and adjacent parts of Norway. A contrarian hypothesis of postglacial tree-immigration patterns. Geo-Öko 21:141–172
Kullman L (2001) Immigration of Picea abies into North-Central Sweden. New evidence of regional expansion and tree-limit evolution. Nord J Bot 21:39–54
Kuosmanen N, Fang K, Bradshaw RHW, Clear JL, Seppä H (2014) Role of forest fires in Holocene stand-scale dynamics in the unmanaged taiga forest of northwestern Russia. Holocene 24:1,503–1,514
Lindbladh M, Bradshaw RHW, Holmqvist BH (2000) Pattern and process in South Swedish forest during the last 3000 years, sensed at stand and regional scale. J Ecol 88:113–128
Lindbladh M, Niklasson M, Nilsson SG (2003) Long-time record of fire and open canopy in a high biodiversity forest in southeast Sweden. Biol Conserv 114:231–243
Lundqvist J (1986) Late Weichselian glaciation and deglaciation in Scandinavia. Quat Sci Rev 5:269–292
Lunkka JP, Johansson P, Saarnisto M, Sallasmaa O (2004) Glaciation of Finland. In: Ehlers J, Gibbard PL (eds) Quaternary glaciations: extent and chronology (developments in quaternary science 2). Elsevier, Amsterdam, pp 93–100
Meltsov V, Poska A, Reitalu T, Sammul M, Kull T (2013) The role of landscape structure in determining palynological and floristic richness. Veget Hist Archaeobot 22:39–49
Molinari C, Bradshaw RHW, Risbøl O, Lie M, Ohlson M (2005) Long-term vegetational history of a Picea abies stand in south-eastern Norway: implications for the conservation of biological values. Bio Conserv 126:155–165
Molinari C, Lehsten V, Bradshaw RHW, Power MJ, Harmand P, Arneth A, Kaplan JO, Vannière Sykes M (2013) Exploring potential drivers of European biomass burning over the Holocene: a data-model analysis. Global Ecol Biogeogr 22:1,248–1,260
Moore PD, Webb JA, Collinson ME (1991) Pollen Analysis. Blackwell, Oxford
Odgaard BV (1999) Fossil pollen as a record of past biodivrestiy. J Biogeogr 26:7–17
Ohlson M, Tryterud E (2000) Interpretation of the charcoal record in forest soils: forest fires and their production and deposition of macroscopic charcoal. Holocene 10:519–525
Ohlson M, Korbøl A, Økland RH (2006) The macroscopic charcoal record in forested boreal peatlands in southeast Norway. Holocene 16:731–741
Ohlson M, Brown KJ, Birks HJB, Grytnes J-A, Hörnberg G, Niklasson M, Seppä H, Bradshaw RHW (2011) Invasion of Norway spruce diversifies the fire regime in boreal European forests. J Ecol 99:395–403
Overballe-Petersen MVAB, Bradshaw RHW (2011) The selection of small forest hollows for pollen analysis in boreal and temperate forest regions. Palynology 35:146–153
Palviainen M, Finér L, Laiho R, Shorohova E, Kapitsa E, Vanha-Majamaa I (2009) Carbon and nitrogen release from tree stumps in boreal clear-cut forests. BOKU, Vienna
Parviainen J (1996) Impact of fire on Finnish forests in the past and today. Silva Fenn 30:353–359
Pirinen P, Henriikka S, Aalto J, Kaukoranto J-P, Karlsson P, Ruuhela R (2012) Climatological statistics of Finland 1981-2010. Finnish Meteorological Institute, Helsinki
Reitalu T, Seppä H, Sugita S, Kangur M, Koff T, Avel E, Kihno K, Vassiljev J, Renssen H, Hammarlund D, Heikkilä M, Saarse L, Poska A, Veski S (2013) Long-term drivers of forest composition in a boreonemoral region: the relative importance of climate and human impact. J Biogeogr 40:1,524–1,534
Sarmaja-Korjonen K (1998) Latitudinal differences in the influx of microscopic charred particles to lake sediments in Finland. Holocene 8:589–597
Sarmela M (1987) Swidden cultivation in Finland as a cultural system. Suomen antropologi 4
Seppä H, Birks HJB (2001) July mean temperature and annual precipitation trends during the Holocene in the Fennoscandian tree-line area: pollen-based climate reconstructions. Holocene 11:527–539
Seppä H, Bjune AE, Telford RJ, Birks HJB, Veski S (2009a) Last 9000 years of temperature variability in Northern Europe. Clim Past 5:523–535
Seppä H, Alenius T, Bradshaw RHW, Giesecke T, Heikkilä M, Muukkonen P (2009b) Invasion of Norway spruce (Picea abies) and the rise of the boreal ecosystem in Fennoscandia. J Ecol 97:629–640
Stocks BJ, Fosberg MA, Lynham TJ, Merns L, Wotton BM, Yang Q, Jin J-Z, Lawrence K, Hartley GR, Mason JA, McKenney DW (1998) Climate change and forest fire potential in Russian and Canadian boreal forest. Clim Change 38:1–13
Sugita S (1994) Pollen representation of vegetation in Quaternary sediments: theory and method in patchy vegetation. J Ecol 82:881–897
Swain AM (1973) A history of fire and vegetation in northeastern Minnesota as recorded in lake sediments. Quat Res 3:383–396
Swain AM (1978) Environmental changes during the past 2000 years in North-central Wisconsin: analysis of pollen, charcoal, and seeds from varved lake sediments. Quat Res 10:55–68
Tallantire PA (1972) Spread of spruce (Picea abies (L.) Karst) in Fennoscandia and possible climatic implications. Nature 236:64–65
Tanskanen H, Venäläinen A, Puttonen P, Granström A (2005) Impact of stand structure on surface fire ignition potential in Picea abies and Pinus sylvestris forests in southern Finland. Can J For Res 35:410–420
Tanskanen H, Granström A, Venäläinen A, Puttonen P (2006) Moisture dynamics of moss-dominated surface fuel in relation to the structure of Picea abies and Pinus sylvestris stands. For Ecol Manag 226:189–198
Ter Braak CJF, Šmilauer P (2002) CANOCO reference manual and CanoDraw for Windows user’s guide: Software for canonical community ordination (version 4.5). Microcomputer Power
Tolonen K (1967) Über die Entwicklung der Moore im finnischen Nordkarelien. Ann Bot Fenn 4:219–416
Tolonen M (1980) Post-glacial pollen stratigraphy of Lake Lamminharvi, S. Finland. Ann Bot Fenn 17:15–25
Tolonen K (1983) The history of Norway spruce, Picea abies, in Finland. Sorbifolia 14:53–59
Vasari Y (1962) A study of the vegetational history of the Kuusamo district (North east Finland) during the late Quaternary period. Ann Bot Soc Zoo Bot Fenn Vana 33:1–138
Wallenius TH (2011) Major decline in fires in confierous forests: reconstructing the phenomenon and seeking for the cause. Silva Fenn 45:139–155
Wallenius TH, Lilja S, Kuuluvainen T (2007) Fire history and tree species composition in managed Picea abies stands in southern Finland: implications for restoration. For Ecol Manag 250:89–95
Whitlock C, Higuera PE, McWethy DB, Briles CE (2010) Palaeoecological perspectives on fire ecology: revisiting the fire-regime concept. Open Ecol J 3:6–23
Zackrisson O (1977) Influence of forest fires on the North Swedish Boreal forest. Oikos 29:22–32
Zillén L, Snowball I (2009) Complexity of the 8 ka climate event in Sweden recorded by varved lake sediments. Boreas 38:493–503
Acknowledgments
The work was funded by the project FIREMAN (NE/G002096/1) in the BIODIVERSA ERAnet and by the EBOR project funded by the Academy of Finland. We further acknowledge support from the Czech University of Life Sciences, Prague (Project CIGA No. 20154309). With special thanks to Lisa Farrell for her assistance with fieldwork in May 2008.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M.-J. Gaillard.
Rights and permissions
About this article

Cite this article
Clear, J.L., Seppä, H., Kuosmanen, N. et al. Holocene stand-scale vegetation dynamics and fire history of an old-growth spruce forest in southern Finland. Veget Hist Archaeobot 24, 731–741 (2015). https://doi.org/10.1007/s00334-015-0533-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-015-0533-z