
Abstract
The purpose of this paper is to discuss recent methodological advances in Argentinean archaeobotany that incorporate the use of ethnobotany as an ethnoarchaeological tool for interpreting ancient food systems in South America. This is an integrative paper that takes into account both published and unpublished results. The role of ethnobotany is examined with reference to ethnobotanical and experimental food processing studies on Prosopis, a wild food plant, and two cultivated ones Chenopodium quinoa and Cucurbita spp., followed by laboratory examinations with microscopy to identify diagnostic changes in plant morphology and anatomy. Experimental materials are then compared with archaeological specimens to identify different types of ancient food processing, and to make inferences about prehistoric post-harvest systems. We demonstrated that: (a) it was critical for our food processing studies to achieve the best taxonomical identification resolution that the plant remains allow; (b) a multi-proxy approach was highly advantageous; (c) ethnobotanical data were crucial to identify food processing pathways of individual plants and combinations of them; (d) the understanding of commensality in the wider sense of the term allows us to determine food patterns both in domestic and funerary contexts. These investigations, the first ones of this type in Argentina, constitute a qualitative step in the methodology for this country because they expand our abilities to interpret the nature of routine plant processing from archaeobotanical assemblages, and they are also a substantial contribution to the development of our discipline in general because the taxa discussed in this paper are distributed throughout South America, as well as in other parts of the world.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The purpose of this paper is to discuss recent advances in Argentinean archaeobotany that incorporate ethnobotany used as an ethnoarchaeological tool for interpreting ancient food systems in South America. Ethnobotany is understood here as the study of the dynamic interrelationship(s) between human societies and plant communities within particular socio-cultural and environmental contexts (Alcorn 1995) and its ethnoarchaeological applications as the study of the living culture from archaeological perspectives (David and Kramer 2001; Kuznar 2001). By means of data obtained from these disciplines, and by developing experimental methods, we were able to find diagnostic criteria for the identification of food pathways of Prosopis spp., Cucurbita spp. and Chenopodium spp. within diverse archaeobotanical assemblages from Argentina. These case studies are described here and discussed from an integrative perspective. They include mostly information already published, although some recently completed unpublished reports contribute with original data.
Because food has social implications that go beyond nutrition and energy, we consider both its role in social practices as well as in routine activities related to subsistence (Twiss 2007 and references therein). Most of the ethnobotanical and archaeobotanical literature on food provisioning, which includes, for example, collecting, allocating/sharing, cooking, eating and clearing up, can be, with more or less variation, represented by the phases of procurement, storage, preparation, consumption and disposal; all of these are embedded in a range of cultural factors such as economics, politics, social or belief systems (Goody 1982; Twiss 2007, 2012). Each of these phases, as Samuel (1996) stated, have different material correlates that allow us to study the archaeology of food. Following Samuel (1996), we use the term “food” in its wider sense, to include raw ingredients through intermediate and final stages of preparation up to the point at which the food product is ready for consumption; we also include by-products and discarded elements associated with food-related sequences of techniques.
Plant resources procurement can also be arranged into the spheres of harvest, post-harvest and consumption (Wollstonecroft 2007). For this paper we are particularly interested in post-harvest systems (understood in the sense of Wollstonecroft 2007) and their potential to stimulate social change (Van der Veen 2003, 2007; Wollstonecroft 2007). Post-harvest systems are comprised of sequences of techniques and technologies for the purpose of promoting and/or retaining plant food quality, quantity and to creating safer and more stable foods (Wollstonecroft 2011; Wollstonecroft et al. 2008). Post-harvest practices are fundamentally linked to consumption patterns but, unlike the act of eating, post-harvest activities leave most of the types of archaeological evidence that can be studied (Wollstonecroft 2007).
As Sherratt (1991) stated, people do not eat species; they eat meals, thus the aim of our research is to move beyond a list of identified species to reconstruct the actual meal or food that could have been consumed. In recognition of the fact that meals may be prepared from a single plant or combinations of plants, we consider both types of meal preparation in this paper. Meal preparation may involve several different methods of food processing, for example mechanical, microbial or thermal activities (Valamoti 2011), each of which may produce different evidence in hearths, ovens, tools (such as mortars, querns, flint and metal blades) and vessels, as well as the deposition of particular food residues and remains at each different stage of processing (Samuel 1996). Therefore, we recognize food processing as an activity that provides an essential framework for archaeobotanical interpretation, since practices of processing lie between the ancient acquisition of plants and the preserved remains of archaeology (Capparelli et al. 2011 and references therein). In the cases presented here we discuss critical information for identifying evidence of plant processing from macro and/or microremains of the three taxonomic genera already mentioned, which come from different types and periods of past Argentine societies (see Materials, below). It is worth mentioning that despite the fact that the history of food processing studies in Old World archaeobotany dates back almost 40 years (Capparelli et al. 2011), in Argentina archaeobotanical interpretations have rarely gone beyond general suggestions of plant use, and processing in particular has scarcely been considered, apart from Babot’s (2009, among others) experimental research into evidence of processing upon starch grains. Food processing sequences are seen here, as proposed by Wollstonecroft (2011, p. 144), as “more than the study of “chaines operatoires”, because it requires an understanding of plants as living, biological entities that are subject to their own physiological processes”, so this type of study allows us to link functional properties of the plants consumed by people in the past with the technology they employed to prepare them, and its implications about their routine activities, dietary choices, health and disease prevention, and dietary change (Wollstonecroft 2011, p. 114).
Finally, and no less importantly, we take into account that meals have a fundamental element which is commensality, that derives from the Latin com = together with, and mensa = table (Pollock 2012, p. 2). Commensality is far more than just the physical act of eating and drinking together, but also comprises myriad social and political elements entailed in each occasion of “co-presence”, each one of which has the act of sharing at the heart of the commensal act (Pollock 2012). In this paper the information presented is examined from an approach that considers commensality within its wider cultural meaning(s). That is, not only for the living people but also for the dead, and other components of the commensal act, such as the surrounding environment that, under local cosmovisions (for further details within local cosmovisions see for example Allen 2002), might need to be “fed” for renewing the power of nature, deities and governors (Lema et al. 2012a). To investigate food in this wide sense, we are of the view that a multi-proxy study (macro + microremains from associated instruments; ethnobotany + experimentation + archaeobotany) is the best approach.
Materials and methods
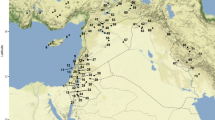
The case studies discussed here were recovered from 13 archaeological sites representing various types of past societies, chronological periods and socio-economic organizations (Table 1). They are located in three main archaeological areas, from north to south (Fig. 1): Northwest, where agriculture replaced hunting and gathering at 3,000–2,000 bp; Midwest, where crop consumption was incorporated into hunting and gathering subsistence by 2,000 bp, and Patagonia, where hunting and gathering continued as the main subsistence pattern throughout the Holocene. All of these sites are distributed along the Argentinean Arid Diagonal, which spans a region from across the northwest of the country to its southeast extremity. This region is distinct in that it has had stable climatic conditions for the last 12,000 years (Abraham et al. 2000), thus we can assume that, in terms of environmental factors, archaeological sites have been subject to similar post-depositional processes throughout this period. Other kinds of post-depositional processes which may have affected archaeobotanical remains were considered in each study case in particular, such as post-depositional fragmentation at Angostura I (see below) and rodent marks at Puente del Diablo (Capparelli and Lema 2011). It is within the Arid Diagonal that the majority of archaeobotanical macroremains have been recovered in Argentina. The most frequent taxa recovered (in terms of ubiquity) are Prosopis, Zea, Cucurbitaceae, Phaseolus, Chenopodium and Capsicum. Few taxonomic identifications were made beyond genus, and in the case of polytypic genera, only rarely was there an emphasis on distinguishing between species.

Location of the Argentinean Arid Diagonal, the archaeological areas comprised in it, and the archaeological sites mentioned in this paper (for reference numbers, see Table 1)
This paper is organised firstly by taxa (Prosopis spp., Chenopodium spp. and Cucurbita spp.). Secondly, results are presented and discussed by research stages: archaeobotanical recognition of relevant features, registration of ethnobotanical information used, and analysis of ethnoarchaeological samples/design of experimental protocols addressing recognition of past food processing in archaeological material. As previously mentioned, this paper’s methodological discussion is made mostly on the basis of published results (Capparelli 2011; Capparelli and Lema 2011; Capparelli and Prates 2015; Lema 2011; Lema et al. 2012b; Lòpez et al. 2011, 2012; Llano and Andreoni 2012; Llano et al. 2012; Ratto et al. 2014). However it also includes original food processing information from the archaeological sites of Las Champas, Finispatriae (Northwest area), Gruta del Indio (Midwest area), Médano I and P26SJ (Patagonian area), some of which were previously described only in internal reports.
Recovery techniques of archaeobotanical macroremains for each study case are described in Table 1. Ethnobotanical investigation used here deals with the recording of local knowledge. Here it was approached mainly by means of direct field research, however, as the past is involved, documents and first-hand information from historical ethnobotany records were used (Medeiros 2010). Interviews, observation and documents, have been used in the same way. Interviews were of an open-ended and semi-structured type, and have been used to explore the procurement, preparation and consumption of food along with observations of daily activities of its preparation. Documents were particularly useful in cases where traditional uses of the studied taxa are not found nowadays, but only historically (Lema 2011; Ciampagna and Capparelli 2012). Field work has been performed in Catamarca, Argentina, and Lipez, Bolivia, where 43 and 10 people have been interviewed respectively (Capparelli and Lema 2011; López et al. 2011, 2012). Modern voucher specimens of plants were gathered during ethnobotanical research, as well as ethnoarchaeological plant material from each processing stage, where possible from Chenopodium quinoa var. quinoa, Prosopis flexuosa and P. chilensis. In most cases, traditional food processing practices were reproduced in the laboratory as experimental archaeology, as detailed below. In the cases of taxa used in similar ways in the past and present (P. flexuosa, P. chilensis in the Northwest area) those experiments allowed us to control several variables that could not be measured in the field, such as characteristics of soaked pod parts during drinks preparation. But experimental archaeology was also useful in the cases of taxa used in the past and present, but in different ways (Prosopis spp. in the Midwest area), taxa used in the past but not used at present (P. denudans, P. alpataco), and combinations of plants that are not used nowadays but which were identified archaeologically (C. quinoa var. melanospermum/Cucurbita maxima). Experimental archaeology allowed us to control carbonization effects in the case of charred archaeobotanical remains (Prosopis spp. and C. quinoa). Archaeological, ethnobotanical and experimental samples were described by means of macro and microscopical observation using SEM and optical and stereoscopic microscopy.
As stated above, we used a multiproxy approach with an emphasis on macroremains, but microremains were considered when necessary. Microremains recovery was done by obtaining three kinds of samples from the active surfaces of stone mortars and pestles that were first cleaned superficially with a synthetic cleaning cloth (Lema et al. 2012b and references therein): First by gentle brushing of the surface with a disposable brush, secondly by scratching with a sterilized metallic instrument, and thirdly by washing with distilled water. In addition, three control samples recovered in the same ways were collected from the non-active surfaces of the artefacts.
Results and discussion
Prosopis spp.
Archaeobotanical remains
Prosopis was present in archaeobotanical assemblages from more than thirty sites in Argentina, including those from societies with a range of different subsistence patterns, and the remains occurred throughout entire occupation chronologies (Giovannetti et al. 2008). In this paper we discuss Prosopis food pathways recognized by examining charred and desiccated pod remains obtained from domestic, funerary, floor and ritual contexts, as well as microremains recovered from stone tool residues (Table 1). The finds correspond to nine archaeological sites in different areas, from Huachichocana, Puente del Diablo and El Shincal in the Northwest, La Olla in the Midwest, Angostura I, P26SJ, Médano I, Aquihuecó and Michacheo in Patagonia, and various periods in the early, middle and late Holocene. The diagnostic features of these archaeobotanical remains are summarized in Table 1 and comprise a variety of Prosopis pod parts with distinctive characteristics, as well as starch grains and resin bodies from the mesocarp of the legume.
Ethnobotanical information and experimental protocols for recognition of past food processing
Ethnobotanical research on traditional uses of Prosopis fruits carried out in northwest Argentina has shown that modern inhabitants use a variety of techniques to prepare various algarrobo products: the pods may be roughly pounded and reduced in boiling water to make arrope jam, or they can be finely pounded to extract the sweet floury mesocarp from which can be prepared patay bread of sun-dried very compacted flour, and other drinks, such as ulpo, a mixture of toasted maize and algarrobo flour in cold water, añapa non-fermented juice or aloja, a kind of beer. In the information from historical chronicles and other ethnohistorical documents we see that most of these practices have a strong historical root deep in time, not only in the Northwest of Argentina (Capparelli and Lema 2011), but also in the Midwest (Llano et al. 2012) and Patagonia (Ciampagna and Capparelli 2012), which is in accordance with the distribution of Prosopis spp. along the Arid Diagonal. Each of the Prosopis products are prepared by using traditional techniques in their own distinctive sequence, but complexity of post-harvest processing of Prosopis pods is evident because the preparation of foods such as patay leaves residues that may be used for the making of drinks such as añapa. Using all this information, we designed experimental procedures for interpreting Prosopis remains from different parts of the Arid Diagonal.
Firstly, we reproduced Northwestern traditional activities in order to evaluate the characteristics of each stage of each type of P. flexuosa and P. chilensis processing (commonly called algarrobo negro and blanco) as well as the final products and residues (Capparelli and Lema 2011). This information, which was also applicable to P. nigra and P. alba respectively, allowed us to distinguish between soaked (drinks) and non-soaked (foods) in desiccated archaeological specimens of algarrobo from the sites Huachichocana and Puente del Diablo. We could identify a predominance in the preparation of aloja or añapa in Huachichocana from the Early Formative (3,000–1,470 bp) to the Inka and Hispanic aboriginal periods (414 bp), while the preparation of algarrobo flour appears to have been more important at Puente del Diablo from the Archaic (10,000 bp) to the Early Formative period (Table 1). Diversification of processing techniques was observed in the latter contexts of both sites, suggesting, especially at Huachichocana, an increasing importance of algarrobo drinks for consumption and probably exchange, between both living and dead people, for Inka society in northwest Argentina (Capparelli and Lema 2011).
We subsequently evaluated the qualitative effect of carbonization on each stage of the experimental food processing procedure as well as the differential preservation of each Prosopis pod part (Capparelli 2011). This allowed us to recognize, at the Inka site of El Shincal, preparation of various food and drink products (Table 1), and a distinctive distribution pattern of pod production within different buildings (Capparelli 2011)..We further identified the grinding of flour, supported by the recovery of Prosopis starch grains from a mortar (Giovannetti et al. 2008).This indicates specialized Prosopis production and suggests post-harvest intensification (sensu Wollstonecroft 2007). Moreover, the ritual significance of Prosopis is suggested by the fact that the largest number of seeds was recovered from the usno ritual hearth, some of which present evidence of añapa or aloja making (Capparelli 2011; Capparelli et al. 2005).
From the Midwest archaeological area site of La Olla (Fig. 1), a residential multiple activity site of hunter gatherers, came the only effective evidence of desiccated patay from Argentina. It was found occasionally by Lagiglia (1956) associated with a human burial context. Recent excavations near the burial allowed the recovery of more Prosopis remains, charred and in association with a domestic hearth (Table 1; Llano and Andreoni 2012). Curiously, algarrobo endocarps from La Olla showed longitudinal and perpendicular marks on their surfaces (Fig. 2a) that were not seen in the material experimentally ground with a mortar, as done in the Northwest. Therefore, new experiments on grinding P. flexuosa were made by Llano et al. (2012) involving the comparison between the results from a stone mill with a more or less level basal stone and a pestle which is moved in a circular motion (Fig. 2b) and those from a mortar with a basal stone with a deep hole and a pestle which is moved vertically, artefacts that were recovered from La Olla (Fig. 2c). They observed that it was milling which left marks on the endocarp surfaces similar to those of the La Olla macroremains and concluded that mills had been used there to make the Prosopis flour. They also observed that, even when the flour produced in a mortar and a mill had the same nutritional values, mortar grinding had lower processing costs than mill grinding; however mortars would have been more costly to manufacture and transport.

Food processing related to study cases of Prosopis. a, grinding marks (arrow) on Prosopis flexuosa endocarps from La Olla archaeological site; experimental grinding on P. flexuosa pods, b, by mill; c, by mortar; D, Prosopis denudans half joints (arrow) from Angostura 1 archaeological site; e–f, normal and polarized view Prosopis starch grain from a mortar recovered at P26SJ location; G–L, microbotanical finds ascribable to Prosopis sp. from a mortar recovered at Medano 1 archaeological site; g, resin body; h–j, tector hairs
Finally, the first Prosopis remains from Patagonia were recovered recently at Angostura I (Fig. 1; Capparelli and Prates 2010). Its interpretation also required new types of experiments for several reasons. First, the Prosopis species identified, P. alpataco and P. denudans, were different from those recovered at the previous sites. Secondly, the recovered charred pods had a particular and more or less homogeneous fragmentation pattern with predominant half joints (Table 1; Fig. 2d). The presence of meso- and epicarp in the majority of the remains allowed us to suppose that these pods were not a residue from grinding for flour. New exercises in experimental grinding of sun-dried pods with a stone pestle and mortar demonstrate that this method does not leave half joints, but clean endocarps and large fragments of epicarp and external mesocarp (Capparelli and Prates 2015). Epicarp and mesocarp tissues of Patagonian Prosopis species are more fibrous than those of the Northwestern ones. From a vast compilation of written chronicles and documents about plants used specifically in Patagonia (Ciampagna and Capparelli 2012), we discovered that the pods were usually toasted before grinding. We reproduced these practices experimentally by embedding pods in hot sand at 400 °C for 2 min and by grinding them in the same way as previously mentioned. We observed that for P. alpataco, P. denudans and P. caldenia, the epicarp and mesocarps are reduced to flour easily if the pods have been toasted. Also, that the fragmentation pattern of toasted pods was similar to those of the charred pods found at Angostura I. Therefore, we pulverized experimentally, with a stone pestle, artificially charred pods to simulate patterns of post-depositional fragmentation and obtained as a result half joints similar to those from Angostura I. We thus concluded that Prosopis pods from the Angostura 1 site had been charred accidentally while they were being toasted (Table 1) and that the fragmentation patterns here correspond to post-depositional factors rather than to grinding for flour (Capparelli and Prates 2015).
Apart from Angostura1, seed remains in Patagonia are scarce, so pestles and mortars or mills may provide secondary evidence of ancient plant processing (Ciampagna and Capparelli 2012). Thirteen archaeological stone artefacts have been analyzed so far, from four different sites of the late Holocene, Aquihuecó and Michacheo with funerary contexts (Lema et al. 2012b) and Médano I and P26SJ that correspond to domestic activity areas (Ciampagna 2012). In six of the analysed artefacts there were Prosopis starch grains. Identifications of Prosopis starch grains are based on previous work of some of the authors, (Fig. 2e–f; Table 1; Giovannetti et al. 2008), together with other plant microremains possibly also assignable to Prosopis (Fig. 2g–j) and to medicinal plants (Prosopidastrum sp.). All of these finds were on active faces of the tools and they were absent on inactive faces, confirming the practice of Prosopis grinding in nearly the 46 % of the analysed artefacts.
Chenopodium spp.
Archaeobotanical remains
Chenopodium quinoa (quinoa) is one of the main subsistence resources for many Andean societies. Its remains have been found at archaeological sites in different parts of South America, in a range of contexts, including hearths, burials, storage structures, human digestive tracts and coprolites (Bruno 2008; López et al. 2011, 2012). Chenopodium from three Argentinean sites (from the Northwest to the Midwest archaeological area) are analyzed here (Table 1). Isolated seeds of C. quinoa var. quinoa (quinoa) were recovered from Finispatriae, while isolated entire fruits of C. quinoa var. melanospermum (ajara) came from Gruta del Indio (Fig. 3a). In contrast, the Las Champas site (Northwest area) presented a culinary combination of C. quinoa var. melanospermum with Cucurbita seeds (Fig. 3b–d; Table 1).

Food processing related to study cases of Cucurbita and Chenopodium. a, unprocessed C. quinoa var. melanospermun (ajara) seed grains from Gruta del Indio archaeological site; boiled ajara seed grains from Las Champas archaeological site; b, general view of ajara stuck to a C. maxima ssp. maxima seed; c, detail of ajara wrinkled testa; d, probable Cucurbita epidermis cells stuck to ajara grain surface; e, experimental boiling of ajara seed grains; f, C. maxima ssp. maxima, thin pericarp, from Pampa Grande archaeological site; C. moschata seed, reference material; g, general view of the seed; h, after experimental boiling, cross section of the seed showing lamination (arrow) in sclereid cells; i, archaeological C. maxima seed from Las Champas site, cross section. B–D and G–T taken from Ratto et al. (2014)
Ethnobotanical information and experiments on recognition of past food processing
Traditional processing of quinoa is very rare in Argentina today and ethnohistorical data scarce; consequently we carried out ethnobotanical field research in Lípez, Bolivia (López et al. 2011). As mentioned above, during the fieldwork we collected quinoa samples and associated knowledge from local people. Specimens of quinoa from long term storage to each post-harvest stage of “enhancement” and to short term storage were collected. By enhancement we refer to a series of detoxification stages that reduce saponins to a non-toxic level (López et al. 2011).The type of grain enhancement varies for each culinary use of quinoa, whether it is to be consumed as whole grains, in soups, or to be reduced to flour known as pitu, and it also varies for each local landrace, and involves a sequence of various techniques including parching, treading, rinsing and rubbing, pounding and soaking. We found that 65–82 % of the processed quinoa grain, after detoxification, included just small parts of the pericarp or none. Quinoa enhanced for pitu preparation takes different forms than those consumed as a boiled grain or in soups because, even when perisperm is preserved well, soaking makes the seed testa wrinkled, and more intense parching leaves a major covering of a brownish colour (López et al. 2011). Once enhanced, the grains can be stored for a short time, mainly in cooking areas. Boiling is the main way of preparing grains that are consumed entire or in soups. As we could not obtain direct samples of ethnoarchaeological boiled grains, we experimented boiling them in water over a hearth heated to 350–400 °C, similar conditions to the traditional ones (López et al. 2012). Enhanced grains of quinoa for consumption entire and in soups and ajara grains (ajara grows nowadays as a weed in quinoa plots) were boiled for 15 min until cooked. After experimental boiling individual specimens of both taxa had: a, the small portions of the remnant pericarp separated from the testa; b, testa wrinkled and heavily folded, and c, perisperm deteriorated and gelatinized (Fig. 3e). However, we could distinguish that while quinoa grains have a fragile aspect, ajara grains had a more robust appearance, possibly due to their thicker testa. Apart from that, we also found that foods prepared by boiling quinoa, enhanced to be consumed as whole grains, could be distinguished from those prepared by boiling quinoa enhanced for soups. Assemblages of the former had a brownish coloured testa, absence of the annular embryo, presence of pits on the perisperm, and a low value (≅ 1.6) for the ratio of whole grain to separated parts of embryo. On the contrary, those of the latter contained several grains with an annular embryo, exhibited pits on the perisperm, deformation of the perisperm and episperm, and a high value (≅ 6) for the ratio of whole grain to separated parts of embryo.
With this information we interpreted C. quinoa var. quinoa (quinoa) finds from Finispatriae (Table 1) as already enhanced grains not yet boiled, due to the absence of pericarp and the testa neither wrinkled nor folded. They may have been accidentally charred and disposed of from cooking areas. In contrast, ajara entire grains from the funerary context of Gruta del Indio, which had the entire pericarp, were found to be non-processed ones (Fig. 3a), which are characteristic of long term storage, according to an internal report sent to Dr. Gil, Natural Science Museum of San Rafael, Mendoza (López, personal communication). On the other hand, ajara grains stuck to Cucurbita maxima seeds at Las Champas, which had only traces of pericarp, wrinkled testa (Fig. 3c), absence of perisperm and embryo and patches of possible Cucurbita seed epidermis (Fig. 3d), were interpreted as grains enhanced for consumption as entire grains that, after enhancing, were boiled (Table 1).
Cucurbita maxima
Archaeobotanical remains
As mentioned in the introduction of this paper, Cucurbitaceae are one of the best represented taxa out of the Arid Diagonal macroremains (Lema 2011). However, from all the recovered fruit part samples of the taxa, post-harvest activities, some of them linked to food preparation for consumption, could be inferred just from funerary contexts of the Las Champas site (Ratto et al. 2014) and from funerary and domestic contexts of the Pampa Grande site, both in the Northwest area (Fig. 1). In the former, the ajara seeds previously described were stuck to desiccated C. maxima ssp. maxima seeds (Fig. 3b), while in the latter, diverse desiccated pericarp remains of C. maxima ssp. maxima and ssp. andreana were found (Fig. 3f; Table 1; Lema 2011).
Ethnobotanical information and experiments for recognition of past food processing
Ethnographically and historically reported uses for C. maxima ssp. maxima in Argentinean local communities mention several methods of post-harvest processing: a, dehydration of halved fruits in hot ashes for storage, which are boiled just prior to consumption; b, cooking the halved fruits in hot ashes and consumption once cold; c, cooking the fruits in pit ovens; and d, fruits cut up and boiled (Lema 2011). In the case of fruits with a thick pericarp, the rind may either be discarded or used as a container. On the other hand, C. maxima ssp. andreana fruits, a species which grows nowadays as a weed in squash plots, must be detoxified in hot ashes prior to consumption. Also, Cucurbita spp. seeds may be roasted and pounded for consumption.
It is interesting to note that, even in cases where food processing pathways of the ajara grains from Las Champas could be recognized, the Cucurbita seeds did not exhibit morphological features that allowed us to infer food processing methods from them. However, in cases where ajara grains and Cucurbita seeds were stuck together we thought that they had been subjected to the same food preparation, probably involving some kind of wetting, before they dried on the ground surface. This suggests that boiling was applied not only to ajara but also to Cucurbita seeds. We carried out experimental boiling of C. moschata seeds to observe whether or not diagnostic characteristics were evident at a micromorphological level (Fig. 3g). Experimentation consisted of boiling approximately 100 squash seeds as part of a starchy vegetable stew. The ingredients, mainly potato and Cucurbita seeds, were put in cold water and once the boiling started it was maintained for 20 min until the stew was ready to eat. Finally the preparation was allowed to cool. Ten squash seeds were then separated, washed and dried for a week in an open space, and observed by optical microscopy. We found that experimental boiling produced a tiny lamination in the sclereid walls and a homogenization of the appearance of the epidermis cells in experimental seeds (Fig. 3h). Unfortunately, sclereid lamination is difficult to determine in ancient seeds (Fig. 3i) and the epidermis is such a particularly soft layer that it is rarely preserved.
With respect to the Pampa Grande pericarp remains, we concluded that cucurbits with thicker rinds were used as containers, because they displayed holes and strings which were probably used as part of the support system (Lema 2011). Indeed, fruits with thicker pericarps are usually associated with this kind of use, while thinner pericarped fruits are commonly associated with food consumption; such may be the case of Pampa Grande. However C. maxima fruits with thick pericarps are also used as containers for cooking stews, like a stewpot. In such cases, the mesocarp of the fruit as part of the consumed food is archaeologically poorly visible. We have yet to understand the type(s) of consumption that might explain Cucurbita finds of both rinds and seed remains, even when they may be charred. However, because of the association of Cucurbita seeds with those of boiled ajara at the Las Champas site, we deduced that it is likely that boiling was one type of ancient Cucurbita food processing method (Table 1).
Summarizing, consumption habits are principal human selection criteria for the plants used and food processing pathways used. However, actual plant consumption is archaeologically difficult to identify unless remains are found in coprolites or the digestive tracts of human remains (Wollstonecroft 2011). But even with such evidence, the reconstruction of food consumption practices from individual food plants is a difficult task unless an integrated approach is employed, comprised of multiple methods of analysis (Van der Veen et al. 2010). A single plant may constitute a meal, or a drink; such is the case of Prosopis, but these are not the most common cases. A meal often involves a combination of several plants or parts of plants. Thus, the Las Champas archaeobotanical ajara/Cucurbita association provides a significant key to recognizing preparation methods. That find, together with a charred mass of various plant tissues recovered from the ritual context of the ceremonial usno ritual hearth of the El Shincal Inka site (Capparelli et al. 2005), where epidermis of Phaseolus and Capsicum were found as part of a single mass, are the unique evidence of culinary combinations to date from Argentina.
Modelling methodological proposals for the understanding of food in archaeological contexts
Our investigations into food processing in different archaeological areas, chronological periods and socio-economic types of early Argentine societies permit us to put forward some important insights into the identification of food processing from archaeobotanical assemblages.
First of all, it is critical to achieve the best taxonomical identification resolution that the plant remains allow. Different species within the same families and genera can produce diverse plant parts and in different quantities, even when they were processed with the same techniques, as with Prosopis flexuosa and P. chilensis. Different species within the same taxon, or varieties within the same species, often require processing in different ways, depending on their physical and chemical properties, as in the case with Patagonian and Northwestern Prosopis species, and with thicker and thinner C. maxima pericarp. Likewise, varieties of the same species may have different ecological traits, require distinct detoxification methods and/or have diverse cultural significance such as the weed-crop complexes in Cucurbita and Chenopodium. The need for high-level species identifications might be obvious to archaeobotanists working elsewhere, but in Argentina it is a polemic issue as most macroremains identifications continue to be made at genus level.
Secondly, it is highly advantageous to use a multiproxy approach for investigating prehistoric food patterns, taking into consideration Samuel’s statement that there is far more to food than the raw materials (Samuel 1996). This approach allows one to take into account the type and part of the plant involved, the mode of consumption, as well as the processing pathways, discard patterns and consequent archaeological preservation potential of the remains. Within this multiproxy approach, the analysis of microremains is a necessary addition to macroremains interpretations, particularly when the latter are scarce, and it permits assignment of functions to some ancient artefacts. As observed above, linking evidence from macro- and microremains has led to a deeper understanding of the different steps of the processing sequences. In microremain analysis it is usual to find a combination of plant taxa, as at Michacheo, with Prosopis and Zea mays starch gains, however it is difficult to discern if this is the result of different processing events of individual plants or a unique processing event of different plants for one or more culinary purposes. A similar limitation affects coprolite analysis. On the other hand, macroremains have a major potential for defining culinary combinations of plant taxa. In addition, as not all taxa produce clear signals of processing, combinations of plants, when found, can be useful for recognizing food processing patterns in mixed remains, provided that a contextual association can be clearly recognized, as with the ajara seed grains with Cucurbita seeds from Las Champas.
Experimentation, based on ethnobotanical and ethnohistorical records, for the purpose of reproducing of traditional plant practices is a crucial way to explore food pathways, in terms of both macro- and microremains as at the Patagonian sites. The reproduction of post-depositional fragmentation patterns and carbonization effects for the interpretation of charred archaeological remains is also necessary. In the case of Angostura1, post-depositional processes were recognized as the cause of the fragmentation patterns, and experiments in grinding fresh and toasted pods allowed us to recognize innovations in post-harvest practices. For example, the possible toasting of Prosopis pods in the hearth, which preserved the materials and increased their archaeological visibility when resulted accidentally charred. On the other hand, experimentation and the study of evidence of discarded material at Huachichocana allowed identification of changes in plant processing methods during the time of the Inka society. This example shows how the study of food pathways might contribute to recognition of changes in social identities (Twiss 2007).
As mentioned before, commensality is understood here in its wider conception (Lema et al. 2012a). This is reflected by the analysis of the processed ajara and Cucurbita “meal” that was deposited together with the human remains recovered from the cave at Las Champas, while unprocessed ajara was recovered from another funerary context in the Midwest area, at Gruta del Indio. The identification of ajara grains from burial sites may reflect a different use of them, compared with the domestic contexts in which quinoa is usually found, as at the Finispatriae site. Here again, the accurate taxonomic identification allowed us to learn about cultural selection and the cultural significance of individual plants. Although currently Bolivian people consume ajara only as a famine food (Bruno 2008), our knowledge about modern ajara food processing is lacking. From the contexts of the Las Champas site, we can conclude that it was processed in similar ways to quinoa grains, but it appears in a different kind of context, a funerary one. It is interesting to note that some traditional Andean societies use it in ritual activities (Nielsen, personal communication). People from Lipez, Bolivia, for example, recognize ajara as a wild seed that represents their ancestors before the Hispanic conquest, while maize represents their Christian side. Thus their current ritual activities include the consumption of both, ajara and maize (Nielsen, personal communication). We can see from these examples how food can be a powerful symbol of identity, as expressed by Twiss (2007), even when it is not literally incorporated into the human body, as occurs in funerary contexts. In this kind of context, ajara meals might represent a commensality that goes beyond daily subsistence, as well as cultural identity. In the ritual context of the El Shincal usno mentioned before, a Phaseolus-Capsicum mixture exemplifies a commensality pattern in which the earth is fed by processed and non-processed plants being deposited in it (Capparelli et al. 2005). It is an interesting paradox that daily consumption, a dynamic activity of any society, is poorly visible archaeologically because the food effectively disappears when eaten, while the feeding of the dead is highly visible in the finds of culturally significant plant material found in graves. However, funerary contexts might also shed light on daily practices, such as for example at the Michacheo and Aquihuecó sites, where stone mortars and pestles, used in everyday life, were deposited with the dead in the burials (Lema et al. 2012b).
Finally, our archaeological and experimental analysis of plant remains from La Olla has shown that food processing also needs to be evaluated in terms of bioaccessibility, the proportion of a nutrient that is released from a food matrix, and bioavailability, the proportion of the food or nutrient capable of being absorbed into the human body, as well as in terms of processing costs and calorific returns, which lead to a more complete view of the whole post-harvest system (Wollstonecroft et al. 2008) and its human potential implications for social change.
Conclusions
Historically there has been little emphasis on going beyond the presence or absence and the possible economic and social uses of plant taxa in archaeobotanical interpretation in Argentina. This study shows how a combination of ethnobotany and experimental archaeology can substantially contribute to the understanding of plant processing patterns, culinary practices, changes in culinary practices through time and space, and functional applications of artefacts, among others. In this paper we have explained how ethnobotanical approaches, used as an ethnoarchaeological tool, can permit the exploration of food as a complex social factor in addition to its commensality; in other words, food systems feed more than simply the necessary daily subsistence practice. Our methodology involves the integration of macro- with microremains from associated artefacts, with ethnobotany by experimentation, and with archaeobotany for the purpose of identifying, interpreting and comparing food processing patterns from hunter-gatherer and agro-farmer societies. These investigations are the first ones of this type in Argentina and constitute a qualitative step in the methodology for this country. This is mainly because they expand our abilities to interpret the nature of routine plant processing from archaeobotanical assemblages. The taxa discussed in this paper are distributed throughout South America as well as in other parts of the world, therefore, although the details of processing practices may vary, we propose that the techniques involving boiling, toasting and soaking and their effects on closely related plants could be similar. For this reason, we believe that the research described in this paper is a substantial contribution to the development of our discipline in general.
References
Abraham de Vázquez EM, Garleff K, Liebricht H et al. (2000) Geomorphology and palaeoecology of the Arid diagonal in southern South America. In: Miller H, Hervé F (eds) Geoscientific cooperation with Latin America. Zeitsch Angew Geol SH1, Hannover, pp 55–61
Alcorn J (1995) The scope and aims of ethnobotany in a developing world. In: Schultes R, Von Reis S (eds) Ethnobotany: evolution of a discipline. Dioscorides Press, Oregon, pp 23–39
Allen C (2002) The hold life has: coca and cultural identity in an Andean community. Smithsonian Books, Washington
Babot MP (2009) La cocina, el taller y el ritual: explorando las trayectorias del procesamiento vegetal en el noroeste argentino. Darwiniana 47:7–30
Bruno MC (2008) Waranq waranqa: ethnobotanical perspectives on agricultural intensification in the Lake Titicaca Basin (Taraco Peninsula, Bolivia). Unpubl. Doctoral Dissertation, Missouri, USA
Capparelli A (2011) Elucidating post-harvest practices involved in the processing of algarrobo (Prosopis spp.) for food at El Shincal Inka site (Northwest Argentina): an experimental approach based on charred remains. Archaeol Anthropol Sci 3:93–112
Capparelli A, Lema V (2011) Recognition of post-harvest processing of algarrobo (Prosopis spp.) as food from two sites of Northwestern Argentina: an ethnobotanical and experimental approach for desiccated macroremains. Archaeol Anthropol Sci 3:71–92
Capparelli A, Prates L (2010) Identificación específica de frutos de algarrobo (Prosopis spp., Fabaceae) y mistol (Ziziphus mistol Grises., Rhamnaceae) en un sitio arqueológico de Patagonia. In: Pochettino ML, Ladio A, Arenas P (eds) Traditions and transformations in ethnobotany (ICEB 2009). CYTED, San Salvador de Jujuy, pp 13–19
Capparelli A, Prates L (2015) Explotación de frutos de algarrobo (Prosopis sp.) por grupos cazadores recolectores del sur de Sudamérica: el caso de Patagonia. Chungara
Capparelli A, Lema V, Giovannetti M, Rodolfo R (2005) The introduction of Old World crops (wheat, barley and peach) in Andean Argentina during the 16th century A.D.: archaeobotanical and ethnohistorical evidence. Veget Hist Archaeobot 14:472–484
Capparelli A, Valamoti SM, Wollstonecroft M (2011) After the harvest: investigating the role of food processing in past human societies. Archaeol Anthropol Sci 3:1–5
Ciampagna ML (2012) Prácticas poscolecta -molienda- de grupos cazadores recolectores de la costa norte de Santa Cruz (Holoceno tardío). IX Jornadas de Jóvenes Investigadores en Ciencias Antropológicas, Abstracts, Buenos Aires, p 68
Ciampagna ML, Capparelli A (2012) Historia del uso de las plantas por parte de las poblaciones que habitaron la Patagonia Continental argentina. Cazadores recolectores del Cono Sur Revista de Arqueología 6:45–75
David N, Kramer C (2001) Ethnoarchaeology in action. Cambridge University Press, Cambridge
Giovannetti MA, Lema VS, Bartoli CG, Capparelli A (2008) Starch grain characterization of Prosopis chilensis (Mol.) Stuntz and P. flexuosa DC, and the analysis of their archaeological remains in Andean South America. J Archaeol Sci 35:2,973–2,985
Goody J (1982) Cooking, cuisine and class: a study in comparative sociology. Cambridge University Press, Cambridge
Kuznar LA (2001) Ethnoarchaeology of Andean South America: contributions to archaeological method and theory. (Ethnoarchaeological Series 4) International Monographs in Prehistory, Ann Arbor, MI
Lagiglia HA (1956) La presencia de patay en una tumba indígena de San Rafael (Mza.). (Notas del Museo nº 1) Museo de Historia Natural de San Rafael, San Rafael, Mendoza, Argentina
Lema VS (2011) The possible influence of post-harvest objectives on Cucurbita maxima ssp. maxima and ssp. andreana evolution under cultivation at the Argentinean Northwest: an archaeological example. Archaeol Anthropol Sci 3:113–139
Lema VS, Capparelli A, Martínez A (2012a) Las vías del algarrobo: antiguas preparaciones culinarias en el noroeste argentino. In: Babot MP, Marschoff M, Pazzarelli F (eds) Las manos en la masa: Arqueologías y antropologías de la alimentación en Suramérica, UNC-UNT. Museo de Antropología de Córdoba, Córdoba, pp 639–665
Lema VS, Della Negra C, Bernal V (2012b) Explotación de recursos vegetales silvestres y domesticados en Neuquén: implicancias del hallazgo de restos de maíz y algarrobo en artefactos de molienda del Holoceno tardío. Magallania 40:229–247
Llano C, Andreoni D (2012) Caracterización espacial y temporal en el uso de los recursos vegetales entre los grupos cazadores-recolectores del sur mendocino durante el holoceno. In: Neme GF, Gil A (eds) Paleoecología humana en el sur de Mendoza: perspectivas arqueológicas. Sociedad Argentina de Antropología, Buenos Aires, pp 57–84
Llano C, Ugan A, Guerci A, Otaola C (2012) Arqueología experimental y valoración nutricional del fruto de algarrobo (Prosopis flexuosa): inferencias sobre la presencia de macrorrestos en sitios arqueológicos. Intersecciones en Antropología 13:513–524
López ML, Capparelli A, Nielsen A (2011) Traditional post-harvest processing to make quinoa grains (Chenopodium quinoa var. quinoa) apt for consumption in Northern Lipez (Potosí, Bolivia): ethnoarchaeological and archaeobotanical analyses. Archaeol Anthropol Sci 3:49–70
López ML, Capparelli A, Nielsen A (2012) Procesamiento pre-consumo de granos de quínoa (Chenopodium quinoa– Chenopodiaceae) en momentos prehispánicos tardíos en el Norte de Lípez (Potosí, Bolivia). Darwiniana 50:187–206
Medeiros MFT (2010) Historical ethnobotany: an approach through historical documents and their implications nowadays. In: Albuquerque UP, Hanazaki N (eds) Recent developments and case studies in Ethnobotany. Sociedad Brasileira de Etnobiologia e Etnoecologia. Núcleo de Publicaçóes en Ecología e Ethnobotánica Aplicada, Recife, Brazil pp 127–142
Pollock S (2012) Towards an archaeology of commensal spaces: an introduction. J Anc Stud 2:1–20
Ratto N, Lema VS, Lopez ML (2014) Entierros y ofrendas: Prácticas mortuorias, agrícolas y culinarias en los siglos XIII y XIV en Tinogasta (Catamarca, Argentina). Darwiniana, nueva serie 2:125–143
Samuel D (1996) Approaches to the archaeology of food. Petits Propos Culin 54:12–21
Sherratt AG (1991) Palaeoethnobotany: from crops to cuisine. In: Queiroga F, Dinis AP (eds) Paleoecologia e Arqueologia II. Centro de Estudos Arqueologicos Famalicences, Vila Nova de Famalicão, pp 221–236
Twiss KC (2007) Home is where the hearth is: food and identity in the Neolithic Levant. In: Twiss KC (ed) The archaeology of food and identity, Chapter 3. (Occasional Paper N° 34) Center for Archaeological Investigations, Southern Illinois University Carbondale, pp 50–68
Twiss KC (2012) The complexity of home cooking: public feasts and private meals inside the Catalhöyük house. J Anc Stud 2:53–73
Valamoti SM (2011) Ground cereal food preparations from Greece: the prehistory and modern survival of traditional Mediterranean “fast food”. Archaeol Anthropol Sci 3:19–39
Van der Veen M (2003) When is food a luxury? World Archaeol 34:405–427
Van der Veen M (2007) Food as an instrument of social change: feasting in Iron Age and early Roman southern Britain. In: Twiss KC (ed) The archaeology of food and identity. Center for Archaeological Investigations, Southern Illinois University, Carbondale, pp 112–129
Van der Veen M, Morales J, Cox A (2010) Identifying foodways - new methodological approaches. Terra Nostra 2010/2, Brune-Mettcker, Wilhelmshaven, pp 91
Wollstonecroft M (2007) Post-harvest intensification in late Pleistocene Southwest Asia: plant food processing as a critical variable in Epipalaeolithic subsistence and subsistence change. Doctoral Dissertation, UCL Institute of Archaeology, London
Wollstonecroft M (2011) Investigating the role of food processing in human evolution: a niche constriction approach. Archaeol Anthropol Sci 3:141–150
Wollstonecroft M, Ellis PR, Hillman GC, Fuller DQ (2008) Advancements in plant food processing in the Near Eastern Epipalaeolithic and implications for improved edibility and nutrient bioaccessibility: an experimental assessment of sea club-rush (Bolboschoenus maritimus (L.) Palla). Veget Hist Archaeobot 17(suppl 1):S19–S27
Acknowledgments
The authors are indebted to the Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET) PIP 0459 funding to Aylen Capparelli, and to Facultad Ciencias Naturales y Museo, UNLP, which has provided institutional and financial support for this work. The authors would like to thank Michèle Wollstonecroft, who gave us very interesting comments that helped to improve the first version of the text, to the anonymous referees for their invaluable suggestions and to the editors and J. Greig for helping us to improve the English writing.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. M. Valamoti.
Rights and permissions
About this article

Cite this article
Capparelli, A., Pochettino, M.L., Lema, V. et al. The contribution of ethnobotany and experimental archaeology to interpretation of ancient food processing: methodological proposals based on the discussion of several case studies on Prosopis spp., Chenopodium spp. and Cucurbita spp. from Argentina. Veget Hist Archaeobot 24, 151–163 (2015). https://doi.org/10.1007/s00334-014-0497-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-014-0497-4