
Abstract
It is known from previous studies that qualitative and quantitative morphological attributes of food products and residues may permit the identification of different food processing activities in desiccated algarrobo (Prosopis chilensis and Prosopis flexuosa) remains. Experimental approaches are used here in order to (1) evaluate if those diagnostic features persist even after charring and (2) to use these results as a means of interpreting types of processing from archaeobotanical contexts of the Inka site of El Shincal. Experimentation was made on the basis of traditional practices which were registered during previous ethnobotanical work in the area. Diagnostic patterns of Prosopis flours and patay (bread), as well as of añapa/aloja (fresh and alcoholic beverages, respectively) and arrope (syrup) residues were stipulated for charred remains. In the specific case of El Shincal, specimens derived from flour manufacture (unrefined and refined) and añapa/aloja residues were recognized from different buildings. This must have implied a special organization of the production and the people working on that, in the sense of post-harvest intensification.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
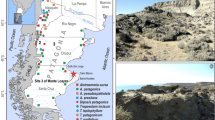
This paper’s main aim is to devise a necessary tool of interpretation of the archaeobotanical record of El Shincal (ESH) archaeological site (Raffino 2004) (Fig. 1). El Shincal is an Inka Administrative Centre located 6 km from the modern village of Londres de Quimivil, Catamarca Province, Northwest of Argentina (27°41′14″ S and 67°10′31″ W). It is situated in the northwestern extreme of the Pipanaco basin at an elevation of 1,300 m above sea level. In Inka times, El Shincal played an important role in the exchange of goods between settlements in the region (Raffino 2004; Williams and D’Altroy 1998). El Shincal main buildings, shown in Fig. 1, have basically only one occupational component (the Inka), however, archaeological evidence recovered from two samples, belonging to both domestic (building K1) and ceremonial (ushno) structures, included the remains of Old World species (cattle, plants such as wheat and barley grains, and peach endocarps) and other archaeological findings, which suggest that ESH continued in this role during the subsequent Hispanic-Indigenous period (Raffino 2004; Capparelli et al. 2005).

Location and map of El Shincal Inka site (modified from Capparelli 2009)
Based on the types of the Inka features present, ESH was regarded as a “New Cusco” by Farrington (1999). The site is made up of numerous types of structures (Fig. 1) including the aukaipata or plaza, a center of political, ceremonial and socio-economic activities and a space of communication between the different buildings of the site—kallankas (i.e. K1, K3)—large structures that were used for a number of different purposes—sinchiwasi (K7)—a large structure with several rooms that may have served as lodgings for labourers—and walled household compounds (RPC; i.e. 5cIII)—rectangular domestic units with patios and roofed areas. The ushno is in the middle of the aukaipata. Food processing areas, mainly spaces containing grinding tools (mortars), are distributed around the central square (see for example “Ruinas” mortars in Fig. 1). A large storage area (i.e. S20E2) is located on top of, as well as behind, a hill found in the northeastern sector of the site (Snead 1992; Raffino et al. 2004a, b, c). This area is comprised of qollqas structures. Qollqas are storehouses which might be similar to those recorded historically by Sánchez Oviedo (see Fig. 6g, h in Capparelli and Lema 2011), but with a one-row round stone basement (see Fig. 1).
Bearing in mind the evident importance of ESH during Inka times, questions arose about the types of economic activities that formerly supported this community. Excavation and recovery techniques at El Shincal were previously described in several papers (see for example Raffino 2004). Archaeobotanical sampling was implemented with systematic flotation. A total of 33 column samples (see grey points in Fig. 1) were taken from 23 architecturally and functionally different structures (see below for more details in recovery techniques, also see Capparelli 1997; Capparelli et al. 2004, 2005). From this analysis, 2,482 non-woody botanical remains were recovered from the Inka floor, and 24 taxa identified (Fig. 2a, b) (see Capparelli 2009). It can be said that El Shincal shows a distinctive pattern of plant use with respect to that of other Inka sites. Wild plants appear to have been as important in the food economy as domesticated species: cultivated foods resources (Zea mays, Phaseolus and Cucurbita) represent 22.4%Footnote 1 of the total plant remains recovered, while wild foods resources (Prosopis, Geoffroea, Zizyphus mistol and Rhamnaceae/Capparidaceae) represent 34.8% (Fig. 2b). Prosopis, together with cultivated maize, were the most ubiquitous taxa of El Shincal food remains (Fig. 2a). Both, Prosopis and maize, were associated to ritual and domestic units and registered the highest percentages of relative counts and densities (Capparelli 2009). But, how can we meaningfully interpret the cultural relevance of one of them with respect to the other from archaeobotanical assemblages? Ethnobotanical and experimental analysis of Prosopis post-harvest practices (can lead to the deposition of different plant parts in different proportions and with different morphological aspects depending on the final product elaborated (Capparelli et al. 2010). Some of these practices can be recognised from Prosopis pod parts deposited in a site, in particular flour remains as well as añapa, aloja and añapa–aloja residues (see. Capparelli and Lema 2011). On the basis of the ethnobotanical and experimental analysis previously mentioned, the objective of the present paper is to evaluate whether diagnostic features previously recognised to be potentially useful for distinguishing ancient Prosopis post-harvest practices from desiccated remains (Capparelli and Lema 2011) persist and are diagnostic even after charring. Finally, these results are used as a means of interpreting types of processing of the Prosopis charred remains recovered from several contexts of the Inka site of El Shincal, as well as to calculate a quantative index of Prosopis use versus that of maize. Data presented here are also part of a major project to evaluate different post-harvest Prosopis trends over time through the analysis of archaeological contexts of different chronological periods of the Argentinian Northwest, before and after agriculture started to be practised (see also Capparelli and Lema 2011).

El Sincal plant taxa. a Ubiquity% (number of samples in which each taxon was present). b Density% (number of charred fragments per 10 l of sediment) (taken from Capparelli 2009)
From ethnohistorical documents, as well as our own ethnobotanical work (see Capparelli 2007, 2008; Capparelli and Lema 2011), we know that Prosopis (species analysed here are Prosopis chilensis and Prosopis flexuosa, known in Spanish as white and black algarrobo, respectively) was an important resource within the Argentinian Northwest, even after the Spanish conquest and up to the present day. The part used is the fleshy and sweet mesocarp of the pod. Pods are dried in the sun and may be consumed raw or parched or may be pounded in stone or wooden mortars into a flour. This may be used to make ulpo, which is a kind of lye, resulting from the dissolution (in water) of algarrobo and toasted maize flour, and patay, which is a bread made from fine, sieved, compacted and dried flour. Flour may be also used to make añapa, a non-alcoholic beverage, for which the flour is soaked in cold water and then manually squeezed to extract the juice, and aloja, which is an alcoholic beverage that is processed in a similar way to añapa, except that the soaked flour is subsequently fermented into a drink similar to beer. A few written sources on Prosopis processing mention also that hot water-soaked pods might be added to the aloja preparation and that chewed pods might be also incorporated to accelerate fermentation (Dasso 2000). A syrup called arrope can be made by boiling the pods, mashing and sieving them through a burlap fabric to obtain juice, which is then boiled until it thickens (for more detail of each type of processing see Capparelli and Lema 2011). These different types of processing may deposit different kinds and quantities of Prosopis parts on the ground where the different activities were carried out. Today, the resultant residues are usually thrown away on the patio floor. Sometimes, they serve to feed domestic animals such as hens.
Archaeobotanists in the Old World have successfully developed interpretative models of crop processing based on ethnographic and experimental observation (e.g. Hillman 1984; Jones 1984). In this paper, experimental analysis carried out on the basis of the modern traditional practices described above, served to characterize the qualitative and quantitative morphological attributes of various charred Prosopis food products and residues, while previous works made it possible to recognize them from desiccated archaeobotanical remains (see Capparelli 2008; Capparelli and Lema 2011). Experimental work on black (P. flexuosa) and white (P. chilensis) desiccated algarroba showed that unrefined flour remains from white algarrobo were mainly seeds with intact testa, entire endocarps—some with mesocarp traces—and halves of endocarps, while black algarrobo showed mostly closed and fissured endocarps—some with mesocarp traces—but almost no seeds. On the other hand, refined flour remains can be recognized in P. chilensis and P. flexuosa by the proportion of seeds with fissured testa, fragmented seeds, fissured endocarps or halves of endocarps, and absence of traces of mesocarp in endocarps. In the case of cold water-soaked añapa and aloja residues, they can be recognized because the testa of the seeds becomes folded/rolled and loosening in some areas. Cotyledons are clearly visible in several seeds. These alterations are most common within those seeds in which grinding produce fissures in the testa. On the other hand, when grinding did not damage all the testa tissues but only seed epidermis, then, just the epidermis may fold when dried. As regards to the añapa-aloja endocarp residues, in those with epi-mesocarp patches, parts of the epicarp become directly attach to the endocarp surface. Epicarp may also curl in the edges. Clean endocarps present the same features that refined flour, but some of them have a slight black mesocarp patina. If fine and coarse flour fractions are used to prepare the añapa or aloja, the small epicarp residues of the fine fraction stuck to the endocarp surfaces when dried. By contrast, hot water aloja residues showed a thick dark patina on endocarps. Chewed pods showed several fiber bundles (from the external mesocarp of the pod) twisted together. Arrope residues are identified by the presence of closed endocarps with a thin dark patina and large pieces of epicarp sometimes twisted. These are the characters which will be evaluated in this paper to know if they are diagnostic for charred remains.
Modern Prosopis post-harvest food practices registered at the Hualfin valley showed very few instances of possible contact, of these algarrobo products and residues, with fire. As these residues are not commonly used for fuel—instead, they are usually thrown away to the patio floor or serve to feed domestic animals—we can suppose that the charred Prosopis remains from domestic-floor and hearth-contexts of El Shincal are derived from the fraction of pod parts that came into contact with fire accidentally. On the contrary, other kind of archaeological contexts, such as the ritual ones, may reflect purposely charred material (this aspect will be discussed below). Different Prosopis assemblages from domestic and ritual contexts will be analysed later in this paper on the basis of the experimental work developed below.
Due to the widespread distribution of Prosopis species as part of many arid ecosystems and also as plants closely related to humans (Beresford-Jones 2005) the subjects treated here may be of interest to researchers in different regions. For example, ecosystems of the Pipanaco basin (the study area of the present paper) were recognized by Simpson and Solbrig (1977) and Orians and Solbrig (1977) as analogous of those of the Silver Bell, Arizona, USA. Beresford-Jones (2005:177), in his Ph.D. thesis establishes a limited analogy between the ethnobotany of the Sonoran desert and the south coast of Peru, which is justified because of biological and ecological similarities of both areas. Therefore, it is clear from these studies that the south west of North America, the south coast of Peru and the southern extreme of the Hualfin valley are areas more or less related in the way Prosopis–people relationships were practised (Capparelli and Lema 2011).
Methodology
We recognize three main factors that may affect charred representation of archaeobotanical Prosopis macroremains differentially with respect to other plant resources: (1) post-harvest processing of Prosopis pods; (2) differential resistance of each different pod part to charring; (3) differential fragility of the remains after charring and their relation to potential fragmentation derived from pos-depositional processes. The response of Prosopis plant parts to these three factors will be analysed in this paper through different experimental designs.
Post-harvest processing of Prosopis pods
Sets of Prosopis chilensis and P. flexuosa unrefined and refined flour, añapa, aloja and arrope were made following traditional techniques previously recorded by the author along the Hualfín Valley, Catamarca Province, Argentina. The protocol was the same of that described in Capparelli and Lema (2011). Sixteen sets of pod parts were charred in different events. Each of the charring sample consisted of the different pod parts of P. chilensis and P. flexuosa (i.e. flours—from which the coarse fraction can be either intermediate product of añapa–aloja preparation or a residue of patay–ulpo production—portions or final products, in the case of patay; and residues, in the case of patay–ulpo, aloja–añapa, aloja and arrope) derived from each food preparation as follows: 1, 2—P. chilensis and P. flexuosa unrefined flour residues, respectively; 3, 4—P. chilensis and P. flexuosa refined flour residues, respectively; 5, 6—P. chilensis and P. flexuosa añapa–aloja residues (refined flour and cold water), respectively, charred wet; 7, 8—P. chilensis and P. flexuosa añapa–aloja residues (refined flour and cold water), respectively, charred dried; 9, 10—P. chilensis and P. flexuosa añapa–aloja residues (unrefined flour and cold water), respectively; 11, 12—P. chilensis and P. flexuosa añapa–aloja residues (fragmented pods and hot water); 13, 14—P. chilensis and P. flexuosa aloja chewed pod residues; 15—patay portion; 16—P. chilensis and P. flexuosa arrope residues, respectively. Each of these set of pod parts was charred in an ash-filled metal bowl that was indirectly heated. Results of experimental carbonization are comparable between them because all the different charring events were charred under a uniform temperature regime. In this sense, temperatures of each charring event ranged from 170°C at the beginning of the process and to 350°C at the end. To prevent specimens’ loss higher temperatures were avoided during this experiment. Temperatures of each event were measured throughout the whole carbonization period with a thermocoupler and thermometer. Duration of the charring process for each individual pod part varied from 5 to 260 min (= 4 h 20 min) depending on the size, hardness and percent of humidity (añapa, aloja and arrope residues were charred both wet and dried) of the vegetal tissue. In another example, in the case of flour residues, seeds of P. chilensis and P. flexuosa charred at approx. 8 min, endocarps at 25 min and entire joints at 65 min. Similar times were observed for añapa–aloja residues charred dried. However, while añapa–aloja residues charred wet took from 70 to 175 min for seeds, endocarps and entire joints of P. chilensis, they took from 185 to 260 min in the case of P. flexuosa, probably due to a higher absorption of water during soaking in the latter. All specimens were analysed and photographed under Stereoscopic microscope, and were photographed with both a digital camera and scanning electron microscope; measurements were taken with callipers, and weighted on a digital balance before and after charring. Entire pods of P. flexuosa and P. chilensis were charred as control samples of non-processed algarrobo, as well as to analyse qualitative features of pod parts after post-depositional fragmentation in order to differentiate them from those derived from flour production.
Differential resistance of each Prosopis pod part to charring
A second charring experiment was carried out for the purpose of observing which of the different Prosopis pod parts were the most resistant to calcination. Joints, epicarp fragments, endocarps, and seeds were charred all together in an ash-filled metal bowl that was directly heated. Temperatures here reached 400–450°C and charring lasted 15 min. Maize grains were also charred and used as a comparative sample.
Differential fragility of the remains after charring and their relation to potential fragmentation derived from pos-depositional processes
After charring, two of each different Prosopis pod part (joints, pieces of epicarp, entire endocarps, halves of P. chilensis endocarps, and seeds) were softly smashed all together with a pestle in a mortar to evaluate relative fragility.
Results
Post-harvest processing of Prosopis pods
The effect of carbonisation on pod fragments and seeds was similar for both species analyzed and is shown in Figs. 3 and 4.

Experimental post-charring effects on Prosopis pod parts. a–c Flour residues. d–g Añapa/aloja residues (cold water soaking). h–i Añapa/aloja residues (hot water soaking). a, c, d, e and g Prosopis flexuosa; b, e, g and i Prosopis chilensis

Breakage by the suture of their endocarps in experimental charred and smashed Prosopis chilensis entire pods. Scale = 1 mm
Flour derivates
Epicarp fragments preserve better when they have mesocarp traces, otherwise, they became very fragile after charring. Mesocarp traces showed a compacted texture (Fig. 3a, b). P. chilensis closed endocarps preserved their aspect, but some of them opened freeing their seeds after charring. P. chilensis halves of endocarps have little potentiality to survive because single shells, like the epicarp, became very fragile after charring.
Some seeds of P. chilensis and P. flexuosa can resist carbonization without changing significantly except for shrinkage in length and width. However, it is more common that splits appear in seeds after charring mainly along the fissure line or along the chalazae area (Fig. 3c). Albumen and cotyledons commonly protruded through these new splits. In seeds that already had fissured testa before charring, albumen and cotyledons commonly protruded along those fissures after charring. In all cases, the portions of testa remaining (whether in an intact seed or between the splits of a protruded seed) still had a good preservation state after charring, even the epidermis, which preserved almost complete. Contrary to what occurs for añapa/aloja residues (see below), testa fragments are not curled or folded.
Changes in dimensions of flour specimens after charring are given in Table 1 . P. flexuosa and P. chilensis endocarps presented almost no change in size, except for an increase of thickness in P. flexuosa, probably due to an increase in seed thickness. P. chilensis halves of endocarps presented no or little variations in length and width but reduced their thickness and weight drastically, becoming very fragile. Both P. flexuosa and P. chilensis seeds shrinkage more in length (10% to 15%) than in width (0% to 2%), becoming rounder, and increased to 55% their original thickness.
Entire non-processed pods
The qualitative aspects of the pods remained almost the same as those before charring. Epicarps persisted around the pod and had a smooth and lustrous surface. Quantitative features changed. Pods shrinkage in length and width (from 7% to 18%), increased in thickness (from 19% to 25%) and lost weight strongly (from 46% to 54%) after charring (Table 2). After smashing the pods in a mortar it was observed that pod joints commonly broke by the suture of their endocarps (Fig. 4), and that fragmented pod piece retains all its endocarp, mesocarp and epicarp sections. It is therefore very difficult to obtain endocarps without mesocarp traces or clean entire endocarps from this kind of fracture. Also, in P. flexuosa, most of the seeds get stuck to their respective endocarp, breaking in small pieces when smashing. On the contrary, in P. chilensis, some seeds are released from the endocarps. These seeds have the testa better preserved, and the testa appears more brilliant and smooth than that of those charred separately from the endocarps.
Añapa/aloja residues
P. chilensis and P. flexuosa epicarp, mesocarp and seeds were more deteriorated after charring than those processed for flour (Fig. 3d, i). The mesocarp in cold water añapa/aloja usually presents holes after charring, and a softer aspect than that of the flour (Fig. 3f, g), which is due to its dissolution in water. This can be seen from values of thickness reduction on añapa/aloja endocarps with mesocarp still attached, which ranged from 19% to 36%, with respect to that of flour, which is null (Table 1). Also, reduction in weight in joint residues of añapa/aloja is almost twice (from 61% to 82%) than in flour ones (from 43% to 45%) (Table 1).
Due to the fact that dissolution of mesocarp in hot water-soaked residues is more intense than that of cold water, some features are more evident in the former. For example, the surface may present waves along the pod, epicarp curls in the edges of individual joints, and mesocarp present larger holes (Fig. 3h, i). Also, in hot water-soaked pods epicarp or mesocarp may separate from the complex endocarp seed (Fig. 3i). These charred epicarp or mesocarp fragments are fragile and can easily be fragmented.
Clean endocarp residues of añapa/aloja presented similar features to those of refined flour after charring, except that some P. flexuosa and P. chilensis closed endocarps opened freeing their seeds, and some P. chilensis endocarps separated into two shells. Black patina of the endocarps, which showed to be useful for distinguishing hot water soaking from desiccated remains (see Capparelli and Lema 2011), it is obviously not useful for charring remains.
Endocarp residues of añapa/aloja showed variations in size after charring. P. flexuosa endocarps became rounder both charred wet and charred dry, because length shrinkage is more than that of width. P. chilensis endocarps became rounder after wet charring but enlarged after dry charring. Thickness reduced after soaking in all the cases except for P. flexuosa dry charring, which presented no variations. P. chilensis halves of endocarps reduced their thickness and weight drastically, becoming very fragile, as it happens for flour specimens (Table 1).
P. chilensis and P. flexuosa añapa/aloja seeds were more deteriorated than the flour ones after charring (i.e. more seeds became deformed by splits, protrusion of albumen and cotyledons, and changes in the testa aspect). In some of the seeds the testa has wrinkled, folded, rolled, and/or strongly split, or disappeared by patches or completely (Fig. 3d). In others, only the epidermis changed. It may present several parallel longitudinal or transversal splits (Fig. 3e), or may fold or disappear by patches. P. flexuosa seed epidermis was more deteriorated than that of P. chilensis.
Seed residues of añapa/aloja usually increase in size when soaked. This increment depends on the water absorbed. If seed epidermis (formed by malpighian macroschlereids) is not injured by the grinding, then, the seed does not absorb any water. But, with a minimal injury of that layer after grinding (what generally happens), the seed absorbed water and increase its size. In seeds with minimally injured testa, which were wet and charred, these increases in size still persist after charring from 3% to 6% in length and width, and from 48% to 82% in thickness (Table 1). But, if the seeds are charred once dried, they shrink from 3% to 12% in length and width, increasing their thickness from 33% to 35% (Table 1).
Similar to what was noted for desiccated remains, charred chewed pod residues are easily distinguished by the presence of very fine threads of mesocarp fiber bundles heavily twisted and occasionally attached to terminal endocarps, however these fine threads of fibers became very fragile and easily fragmented after charring.
Conglomerates, coming from the desiccation of fine flour añapa/aloja residues (see Capparelli and Lema 2011), separated into individual pieces after charring. It is not possible to distinguish what the grade of refinement of the flour used from charred remains was. On the contrary, however, those conglomerates coming from arrope residues that were charred when wet can be recognized (see below).
Patay
The patay bread become more compacted and hard when charred.
Arrope residues
Endocarps having a slight black patina are characteristic of desiccated arrope residues, but this is not preserved after charring. Pieces of twisted epicarp were even more curled after charring and became very fragile. Wet and charred residues (endocarps and pieces of epicarp) became stuck together, but not those dried and charred. Dimensions could not be measured because of the fragmentation and destruction of pod joints.
Other considerations
With respect to comparing specimens that were wet and charred with those dried and charred, the former had greater amounts of ash adhering to their surface, and sometimes showed patches of a brownish colour instead a deep black one. If wet añapa/aloja residues are taken to the calcination point, then spheres of grey and brownish ashes replace each pod joint.
Differential resistance of each Prosopis pod parts to charring
As regards the results of the second charring experiment, it is worth mentioning that except for individual joints, that were 100% recovered as well as maize kernels, the loss of Prosopis pod parts ranged from 20% to 85% after charring (Table 3). This loss was numerically different between each category (such as endocarps/seeds/epicarp–mesocarp pieces). The recovery of epicarp fragments and seeds reached 80% of the initial number. However, endocarps were the most susceptible to conversion into ashes, even more than seeds. In the case of P. flexuosa, despite the production of abundant entire endocarps with grinding, these fissured or were broken with charring. Also they have a much higher likelihood of being destroyed by charring than P. chilensis closed endocarps. Endocarp tissue are composed mostly by fibers and schlereids, but in the case of P. flexuosa the fibers are shorter and are positioned in a looser and less compacted way than in P. chilensis. The ligneous constitution probably makes them calcinate before seeds do. In the case of P. chilensis, tissue of each shell is very thin, fibers are longer and more compacted than in P. flexuosa. In the experiment developed here from 20 original charred endocarps, just three and a half endocarps and three seeds remained after charring. It is clear that calcinations of the seed may take place with more intensive charring.
Differential fragility of the remains after charring
The smashing of the different Prosopis pod parts affected most to epicarp fragments and halves of P. chilensis endocarps. The pounding of these specimens showed that epicarp fragments and single shells of P. chilensis endocarps, despite having a high recovery rate after charring, are the most fragile pod parts compared with individual joints, seeds and those entire endocarps that resist charring.
Finally, some features useful either for desiccated or charred remains, share limitations. For example, those characters related to changes in testa seed of añapa/aloja residues, which are highly diagnostic for post-harvest practices, affect diagnostic features (fissure line, among others) that are taxonomically important for genus or species recognition. Therefore, taxonomic identification might be possible just for a few of the remains. Other limitations may appear if comparisons are going to be made between this experimental results and remains that might have been charred at more intense regimens. We do not exactly know at which temperatures were El Shincal remains charred, information that will be object of future works. By now, we assume that charring regimes were similar in experimental and archaeological cases. In this sense, even when it is more difficult to recognize post-harvest practices in charred specimens than in desiccated ones, a conjunction of qualitative characteristics of pod parts present in each assemblage (some features related with changes in dimensions after charring might be also useful), their relative quantities, and their contextual associations with other archaeobotanical and archaeological remains can help to recognise charred Prosopis specimens derived from different processing practices. A summary of the most important diagnostic features for the recognition of potential post-harvest processing of Prosopis pods for food, which considers the results of the three different charring experiments presented here, and are described on the basis of the previous data obtained from desiccated remains are shown in Table 4. Similarly to what was observed in the case of the quinoa (Chenopodium quinoa) (see López et al. 2011) these different kind of preparations are more probably to be recognizable from contexts where final stages of processing were carried out, also from those where the preparations are disposed for consumption, and finally—in the case of unrefined or refined flour—from storage contexts. As a complement of Table 4, a general dichotomy key was elaborated in order to potentially distinguish each type of processing from Prosopis charred archaeobotanical samples, as follows.
Dichotomy key to potentially distinguish each type of processing from Prosopis charred archaeobotanical samples:
-
A—resinous masses of mesocarp flour strongly compacted with little epicarp fragments spread within the mass—fine mesocarp flour- patay
-
AA—Other remains—B
-
B—Closed and clean endocarps as the predominant fraction of the sample, together with large pieces of epicarp sometimes twisted, which became very fragile after charring and have low possibilities to survive—arrope residues Footnote 2
-
BB—Seeds, endocarps (with or without mesocarp), and sometimes joints, constituted the assemblage—C
-
C—Seed testa normal; neither rolled nor folded, with good preservation state, even the epidermis, which can be almost complete. Splits may appear in seeds after charring. Albumen and cotyledons commonly protruded through these splits. Epicarp fragments, when present, not rolled neither folded on the edges. Mesocarp, when present, with compacted texture—D
-
CC—Seed testa mainly rolled, folded, or with parallel splits. Testa may disappear by patches or completely, leaving cotyledons visible in some cases. Also, folded pieces of testa separated from the seed may appear. Some seeds charred wet may increase their size. Epicarp fragments, when present, may appear also rolled and folded on the edges, and directly attached to the endocarp surface because of the dissolution of the mesocarp. Mesocarp, when present, usually with holes, and a softer aspect than that of the flour. Low archaeological visibility in general, because tissues remnant after soaking are highly deteriorate—E
-
D—Presence of mesocarp tissue in some pod parts, whether as part of individual joints or in patches over the endocarp surface. Individual joints may be fragmented by the endocarp suture line as a consequence of post-charring fragmentation—residues of unrefined flour
-
DD—Mesocarp tissue, if present, is very few. Endocarps, when present, mostly with clean surface. But in fact, whole endocarps presented a lower resistance to charring with respect to seeds, and halves of endocarps of P. chilensis have little potentiality to survive because single shells, like epicarp, became very fragile after charring. Seeds (mostly in the case of P. chilensis) are predominant—residues of refined flour
-
E—All features described in CC present—residues of añapa–aloja
-
EE—All features described in CC plus very fine threads of mesocarp fiber bundles heavily twisted and occasionally attached to terminal endocarps (these fine threads of fibers became very fragile and have little potentiality to survive because they are easily fragmented after charring—residues of aloja
-
-
-
-
Application of the experimental results to the case of El Shincal Inka site
Previous researches on the site and the archaeobotanical analysis
This site was widely studied during nearly 20 years. Raffino (2004) resumes archaeological, archaeobotanical and ethnohistorical investigation on the area. Archaeobotanical studies started by the author during the 1990s decade and continue up to now (for more detail on recovery methods, identification procedures and interpretation of plant remains see Capparelli 1997, 2009; Capparelli and Raffino 1997; Capparelli et al. 2004, 2005). The most important aspects of these studies that are relevant for this paper are briefly described as follows.
Recovery techniques (for more details, see Capparelli 1997; Capparelli and Raffino 1997; Capparelli et al. 2004, 2005)
A total of 861 l of sediment from 33 column samples (see grey points in Fig. 1) were processed by machine-assisted flotation. The machine utilized had a linen bag to collect light fraction, while a mesh size of approximately 1.5 mm was used to retain heavy fraction (see Capparelli and Raffino 1997).
Each of these column samples, from 20 × 20 cm, covered depths from 0 to 90 cm. They were taken from 23 of the main architecturally and functionally different structures of the site (one to four from each architectural structure depending on the case) (see Fig. 1, grey points). Near a hundred of flotation samples ranged between 6 and 8 l of sediment were processed. In the laboratory, light and heavy floated fractions were sieved with a 2 and 0.4 mm meshes. Fractions >2 mm were sorted by eye, while those between 0.4 and 2 mm were fully sorted under a stereoscopic microscope. Material from screening and manual recovery during archaeological excavations was also examined.
Depending on the post-depositional accumulated sediment, the Inka floor at El Shincal extends at an average of 60–70 cm deep from the modern ground surface. However, we observed high densities of remains in a wider range, between 40 and 90 cm deep, which might be the effect of vertical migration of remains due to root intrusions of the dense modern vegetation cover. Therefore, layers of column samples in between 40 and 90 cm were considered for this paper, similarly to the methodology applied in a previous publication on El Shincal Inka economy (see Capparelli 2009). The Hispanic-aboriginal cultural event mentioned above was represented at El Shincal only in the ushno and the K1 northern sample, while in the rest of the samples just the Inka event was present. In the case of the ushno, both Hispanic-aboriginal and Inka event formed a palimpsest difficult to distinguish one from the other (see below). In the case of K1 northern sample, wood charcoal was the unique plant remains recovered, for what it was not considered in this paper. Less productive deposits were used as controls for evaluating the remains in features and richer deposits.
General archaeobotanical results and the recognition of post-harvest practices from El Shincal specific Prosopis findings
Including the wood charcoal, the absolute fragment count of archaeobotanical remains recovered at El Shincal was n = 13,116. Excluding the wood charcoal, the absolute fragment count was n = 2,482. The number of identified taxa was 24 (including seeds, fruit, parenchyma tissue, rhizomes, stems, taproots, but not wood charcoal). In general, the plant remains from ESH are well preserved, possibly due to their being protected by a post-depositional layer of sediment measuring 40–70 cm in depth.
One thousand eleven charred Prosopis macroremains were recovered from the Inka floor occupation (see Table 5), which were analysed in this paper in terms of the recognition of Prosopis post-harvest practices. These remains come from domestic floors and small charred lenses (S20E2, K1, Aukaipata), domestic hearths (Sinchiwasi, 5cIII, K3), as well as from ritual hearths (ushno). They consisted predominantly of seeds, but a small fraction of individual joints fragments and an even lower quantity of endocarps were also recovered (Table 5).
Discussion: ESH remains interpreted on the basis of the experimental results
A distinctive pattern of Prosopis pod parts distribution was recognized at ESH through the analysis of the most diagnostic remains, which is interpreted to be associated with different post-harvest practices at different buildings, as follows.
Aukaipata, K3 and S20E2 are treated together because of similarities in Prosopis remains.
Aukaipata is a center of political, ceremonial and socio-economic Inka activities (Raffino 2004:261; D’Altroy 2003:241) and a space of communication between the different buildings of the site. The most diagnostic remains coming from the aukaipata floor context consisted predominantly of P. chilensis and P. flexuosa fragments of endocarp tissues with their correspondent mesocarp and epicarp, both covering the entire endocarp surface (Fig. 5a, b). Experimental analysis showed that these kinds of specimens are produced as a consequence of a post-charring fragmentation, in this case of individual joints.

Prosopis archaeobotanical remains from ESH: flour remains. a Aukaipata; b S20E2; c–d K3; e–f Sinchiwasi; g 5cIII. Scale = 1 mm
Kallanka K3. Kallankas were used for a number of different purposes by the Inkas: (1) for public feasts, (2) as multiple purpose areas and (3) as occasional lodgings for soldiers (Raffino et al. 2004a; D’Altroy 2003:241). From the domestic hearth in K3, remains consist mostly of endocarp tissues with their correspondent mesocarp and epicarp, like those recovered from the aukaipata, as well as endocarps with traces of mesocarp (Fig. 5c). This mesocarp does not present aspects of having been soaked before charring because the texture is compact. Other plant remains found there include an endocarp of Geoffroea decorticans (chañar) and one maize kernel (see Capparelli et al. 2005).
S20E2
This place is part of a large storage area of the site. Endocarps with mesocarp traces (Fig. 5d) and individual joints of P. flexuosa were the most diagnostic Prosopis remains recovered here together with desiccated rhizomatous fragments of Prosopanche americana, which can be either ancient or recently intrusive.
These three structures are spatially close and share similar kinds of remains which permit us to infer the use of unrefined Prosopis flour. The kind of Prosopis assemblages at these places is similar to those assemblages coming from recent excavations carried out by Giovannetti (2009) on the sediment around two bedrock boulderss which contains numerous mortar holes at El Shincal, the “Ruinas” and “EGP” complexes. The first one is placed near 5cIII buildings, while the second one is located on the cultivation field around the site. From these excavations several Prosopis pod fragments and endocarps with traces of mesocarp were recovered, in a proportion nearly to 1:1 with seed remains (Giovannetti 2009). Other associated plant remains were maize, beans and chañar, among others. Grinding produces a lot of pod parts which spills off the grinding surface during processing. For example, it was reported by Cáceres Freyre (1962) that in the use of the “cimbra” (a particular wood structure with a heavy stone used to grind Prosopis pods in the Pipanaco basin during the first half of the twentieth century, see Capparelli and Lema (2011) for more detail) it was necessary to have two people helping with putting the spilled parts of the pods back on the grinding surface. This habit can be also observed in modern maize grinding (see Fig. 7h in Capparelli and Lema 2011). The spilling out of the Prosopis unprocessed, or partly processed pod parts, during grinding is consistent with the high quantity of individual joints, still with edible mesocarp attached, near the grinding areas found by Giovannetti (2009). The used of El Shincal mortars for Prosopis grinding was confirmed also through the recovery of Prosopis starch grains from the grinding surface of one of the EGP individual mortar holes (Giovannetti et al. 2008). In sum, remains from the aukaipata, S20E2 and K3 are interpreted as specimens of unrefined flour that may had been accidentally charred while being transported from the storage to the further processing/consumption areas.
Sinchiwasi
Rooms of the Sinchiwasi, such as room R10, include two terraces (“beds”) on opposite sides that are each separated from a central hearth area by a partition wall. They may have served as lodgings for labourers (Raffino et al. 2004b). On the opposite of what occurs at the previous structures analyzed, the Ue7 of the R10 central hearth presented a Prosopis assemblage constituted only by seeds. From this context 312 Prosopis seeds were recovered (Fig. 5e, f), together with 56 pieces of highly fragmented G. decorticans (chañar) endocarps and five small pieces of maize (grain and cob) (Capparelli 2009). From the 312 Prosopis specimens, 169 were entire seeds. 97% of them were preliminary identified as Prosopis chilensis, while only 3% were ascribable to P. flexuosa. Footnote 3 The dominance of white algarrobo seeds in this assemblage may be due to the fact that this species releases almost all seeds during fine grinding, which does not occur with P. flexuosa. The testa of these seeds was normal; neither rolled nor folded (Fig. 5f). In most cases, the endosperm had burst out of the seed along the fissure line and/or the edges (Fig. 5e). The large proportion of P. chilensis seeds with neither rolled nor folded testa, but with protrusions of endosperm along the fissure line, together with the absence of joints and endocarps with mesocarp–epicarp traces, permit us to interpret these remains as the result of preparing refined flour. This refined flour could have been used to prepare aloja, patay or ulpo (see Capparelli and Lema 2011). It is interesting to note that this arquitectural structure is interpreted as lodging guests/foreign labourers from the Chaco region (suggested by the ceramics present, Lema et al. 2009). Since Chaco is an area where algarrobo is even more abundant than in El Shincal, it is possible that these people had brought to the site knowledge on how to prepare various Prosopis foods.
5cIII
From this room, only five Prosopis seeds of P. chilensis and P. flexuosa were recovered, together with some seed fragments. A diverse archaeobotanical sample was recovered from this feature. 5cIII appeared to have been a kitchen building, where mostly useful parts of cultivated species were found, together with ordinary ceramics, in a central hearth fired with shrub woods (Capparelli 2009). Surprisingly, Prosopis remains were very scarce here, with respect to cultivated plants such as maize (kernels), squash (seeds) and beans (cotyledons). Although it is clear that the emphasis was not put on Prosopis processing in this building, some activities involved this species. Because seeds are the only part present, we infer that refined Prosopis flour was used here. Seed testa in the 5cIII remains is much deteriorated, with other plant tissues sometimes stuck to the folded testa (Fig. 5g). Unfortunately, up to now we cannot recognize from these seeds diagnostic characters that identify the processes that were applied on them.
K1
The identification of this structure as a kallanka, a space for multiple activities, was confirmed in K1 by the variety of artifacts found here: mortars, clay, bone instruments, flints, instruments for spinning wool, pucos and globular bowls. Relatively high plant diversity was encountered here: Prosopis maize kernels and cobs, and cotyledons of beans, and Solanum seeds were recovered, among others (Capparelli et al. 2005). However, densities of botanical remains were very low. Several explanations were proposed for this low density of plant remains. Perhaps the floors of the rooms were frequently swept out, or there was only a low level of activities involving fire (see Capparelli 2009 for more detail). This is consistent with the fact that just small lens of charred remains were found here. Prosopis seed findings from K1 are the only ones that present characteristic of añapa/aloja residues (Fig. 6a). They are represented by folded pieces of testa separated from the seed (Fig. 6b), some of them rolled (Fig. 6c, d). The manufacture of añapa/aloja by cold water soaking (not involving fire) is thought to have been carried out here with refined flour already prepared in other places of the site. These results are consistent with early ethnohistorical written documents of the area, such as those about the moving of the city of San Juan de Vera del Valle de Londres (presumably Londres de Pomán, situated on the East of the Pipanaco basin) to the Catamaca valley, and dated between 1679 and 1681 ad (Charcas 23 R7 N71, Archivo General de Indias, Sevilla). This document describes the arboreal spieces of Prosopis in the Pipanaco basin and its intense utilization by aboriginal people. In one paragraph, the Spanish people say that the aborigines are used to get drunk with strong drinks which they made from the algarrobo fruit, which is abundant in that region (“… otros vicios de embriaguez que lo exercitan con brebajes fuertes que hacen de la algarroba que tienen en abundancia en aquella Juridición…”) (Capparelli 1997).

Prosopis archaeobotanical remains from ESH: añapa / aloja residues. a–d K1; e–j, ushno. Scale in (a, c, i and j) = 1 mm
Comparisons between the archaeobotanical assemblage of K1 and those of K3 showed significant differences (Capparelli 2009). For example, samples from the K1 floor and lens contexts were composed of 38.4% of cultivated species and 27.5% of wild food remains (all Prosopis seeds). Samples from the K3 hearth context were composed of only 3.1% of cultivated plants and 72.1% of wild food plants. These differences were interpreted as indicative of different sets of activities carried out in each of the two kallankas. More specifically, a greater emphasis on the processing of cultivated than wild resources, including maize kernel separation from the cobs, was proposed for K1; and an emphasis on the managing of wild resources was proposed for K3 (Capparelli 2009). However, as regards to the type of Prosopis post-harvest practices detected here, such as the manufacture of añapa/aloja, may lead to an underestimate of Prosopis use in this building. This is because of a low archaeological visibility of añapa/aloja remains due to non-use of fire, and also because tissues after the soaking stage are highly deteriorated. Also, the disposal of Prosopis residues could have involved feeding them to domestic animals.
Ushno
The greatest quantities of Prosopis remains of ESH were recovered from the ushno context, together with, chañar, maize, beans, Cucurbita, among other native plants, as well as to European crops, such as wheat, barley, and peaches (see Capparelli et al. 2005; Raffino et al. 2004c). The assemblage was formed by several Prosopis seeds and a few closed endocarps (possibly of black algarrobo); epicarp or mesocarp pieces were absent. The major portion of the ushno seeds consists predominantly in seeds with split testa (Fig. 6e), with folded testa (Fig. 6f), and without testa (Fig. 6g, h). As we see from experimental analysis, the lost of the testa seed is common for aloja/añapa residues. In the ushno, also, seeds are larger than those of the other parts of the site (see Fig. 6h). It can be argued that these seeds are the result of the use of pods larger in size with respect to those of the Sinchiwasi, for example. But the few endocarps recovered showed standard endocarp sizes, taking into account an expected increase in size after soaking.
The high absolute counts and high densities of archaeobotanical remains that were recovered from the ushno may be the result of a persistent use of this ceremonial place during the Hispanic-Indigenous period (Capparelli et al. 2005). The richness of the archaeobotanical assemblage of the ushno may also be partly due to the fact that ritual contexts have greater preservation potential than domestic contexts for several reasons (Capparelli 2009), including: a diversity of burning activities; the selection of plant organs with unusual characteristics, (i.e. larger than the regular ones); as well as the high frequencies of plants deposited in the features as a result of repeated use and/or a deposition of high quantities of material during each activity. Another factor that may have promoted preservation is that the types of fires used in the rituals may have been more suitable for the charring and preservation of plant remains than other activities carried out at the site. In the case of Prosopis remains, the fact that they were purposely charred at the ushno ritual hearth and not accidentally charred (as in the other domestic contexts) may explain the highest quantity of seeds recovered. Añapa/aloja residues commonly decanted from the jar where they were prepared could easily fall into the fire when the liquid fraction of the drink was offered.
The site as a whole
Unrefined flour was recognized from S20E2, Aukaipata, and K3 assemblages, while an advanced grade of Prosopis processing was observed in R10 of Sinchiwasi, K1, ushno and 5cIII. The characteristics of these Prosopis assemblages of macroremains at ESH suggest that edible algarrobo pods may have reached the site after a first stage of processing. As regards the archaeobotanical findings recovered around each excavated stone mortar of ESH (see Giovannetti 2009), as well as starch grains analysis of one of this mortars (Giovannetti et al. 2008), the grinding of Prosopis pods must have taken place at the grinding stones placed around the main buildings of the site. Pods could also have been ground on “cimbra” type mortars similar to those that were registered in use by Cáceres Freyre at the beginning of the twentieth century on the bottom of the Pipanaco basin (see Cáceres Freyre 1962; Capparelli and Lema 2011). Whether grinding was finished in the large stone mortars area around the site or inside some of the main buildings is still unknown.
Modern ethnobotanical data show that pods are dried on the “cañizos”, built on the fields that surround the residential area, and then are ground and stored in bags in storage rooms (see Capparelli and Lema 2011). In sum, structures similar to the “cañizos” might have been built at ESH on the field that surrounds the main construction of the site for drying up Prosopis pods, which might have been coarsely grinding on the mortars around the site previously to take them into the main buildings.
Prosopis vs maize
A gross numerical estimation of Prosopis processing is developed here. Although we recognize that this kind of quantitative comparison has biases, we consider that this estimation can give a better sense of the variables affecting Prosopis remains in archaeological representation.
The weight of Prosopis remains recovered at domestic contexts of El Shincal, which are thought to have been charred accidentally (the ushno was not taken in account because it might have implied purposive charring) was 4.57 g. Supposing arbitrarily that just the 1% of the total volume of residues produced are finally charred (this includes also differential resistance to charring of Prosopis pod parts), and supposing also that all the 4.57 g are residues (in fact, most of them are, because they are seeds), the original quantity of residues prior charring would have been 457 g. If this value is multiplied by the loss of weight during carbonization (50% on average), the remains recovered would represent 914 g of Prosopis residues. We know by experimental analysis that entire recognizable seeds after refined flour production ranged around 12–20% (a median of 16% is used here) of the total original seeds from the pods (Capparelli 2008; Capparelli and Lema 2011). Therefore, these residues would represent a total of 5,712.5 g of residual seeds. Each seed weights an average of 0.04 g, and thus 142,812.5 seeds are represented by this quantity of residues. Each pod has an average of 17 seeds (Capparelli 2008), and then 8,400 pods would have been processed. Each pod weights in average 3 g, thus, at least 2,800 g of pods would have been processed at ESH. This quantity might represent, for several contexts, the result of just the last set of activities carried out by people on the site, if we take in account that activity floors and hearths may have been currently swept or cleaned, respectively.
From previous estimations of production of cultivated resources at El Shincal, made on the basis of the extrapolation from the subsistence requirements of one person using fallow and permanent agricultural systems, the cultivated surface of ESH (approximately 726 ha) could sustain from 1,613 to 2,792 people (Capparelli 1997:162). This is many more than the minimum of 586 inhabitants calculated by Raffino (2004:29) to have been living there.Footnote 4 However, even when Inka people were able to produce enough cultivated food resources, such as maize, beans and squash (Capparelli 2009), Prosopis pods may have played an important role as food/drink products in the daily live of ESH inhabitants
These results are consistent with the use of Prosopis by other native North, Central and South American people, mentioned by Beresford-Jones (2005), who noted that Prosopis was very important to “incipient cultivators” in the Tehuacán Valley, Mexico, the south-western USA, the coast of Peru and the coast of Chile (see Beresford-Jones 2005 for a review of that evidence). He also maintained that even the agriculturalists of the Sonoran desert, “such as the Akimel O’odham and their predecessors the Hohokam, depended upon wild foods for up to 50% of their food supply. By far the most important of these were Prosopis beans…” Beresford-Jones et al. (2009:323).
Final considerations
The recognition of different Prosopis post-harvest practices at El Shincal represents an advance in the understanding of Inka societies and its economy. Up to now, the importance of algarrobo’s food products to native South American people and to Spaniards living in these areas during the 16th to the 18th centuries was attested by historical records (see Cobo (1653) and Cieza de León (1553) in Beresford-Jones 2005:229; De las Casas in Yacovleff and Herrera 1934:291; and other records in D’Antoni and Solbrig 1977; Boixados, 2002; Quiroga 1999; Noli 1999). Results from El Shincal analyzed here, and from Huachichocana III (Capparelli and Lema 2011) are the first archaeological sites in which Prosopis findings were analyzed in order to recognize different post-harvest practices, and from where the manufacture of Prosopis flour and drinks by the Inka state can be confirmed.
In the Kollasuyu territory (the southern section of the Inka state, see Raffino 2004), Prosopis remains have been recovered from several sites. Surprisingly, all of them are from Argentina, and six out of seven come from the modern province of Catamarca where El Shincal is located. These Catamarca sites are Potrero-Chaquiago (Williams 1985), Punta de la Peña 3-La Tranca (Rodríguez pers. comm. Intihuatana de Fuerte Quemado (Kriskautzky and Morales 1999); Mishma 7 (Sempé de Gomez Llanes 1986); Batungasta (Ratto et al. 2007); and Los Colorados (Giovannetti 2009). The exception is Huachichocana III, located in the modern Jujuy province (Capparelli and Lema 2011). ESH provided one of the largest finds of Prosopis in Kollasuyu, by comparison to the sparse finds at other sites. Food/drink products prepared at ESH were diverse and took place at different buildings. This implies a special organization of Prosopis production, in the sense of a post-harvest intensification (sensu Wollstonecroft 2007). In Inka times, El Shincal played an important role in the exchange of goods between settlements in the region (Raffino 2004; Williams and D’Altroy 1998), and Prosopis may have been one of those exchanged resources. On the other hand, the sacred associations of the Prosopis tree are ancient in the Andes (see discussion in Beresford-Jones 2005:224) and Prosopis at ESH may have had also a special significance for the people living there, since the largest number of seeds recovered from the ritual hearth of the ushno.
As result of the homologous role of Prosopis to humans in various similar ecosystems (the Southwest USA, Peru, Chile, and elsewhere in Argentina), the experimental analyses presented in this paper might be useful for interpreting ancient Prosopis processing outside the Hualfín valley.
Notes
Values based on density counts (for more details, see Capparelli 2009)
Ancient arrope have to be interpreted with caution because some ethnobotanists, based on ethnohistorical written documents, support that its manufacture for Argentina is post-hispanic (see discussion in Capparelli 2007)
Identification is based on qualitative and quantitative features of seeds and endocarps of diagnostic value that authors are preparing for publication (Capparelli et al. In Prep.).
Raffino based his estimation on the relationship of one inhabitant per 3 m2 of the area corresponding to the buildings potentially roofed
References
Beresford-Jones DG (2005) Pre-hispanic Prosopis human relationships on the South Coast of Peru: riparian forest in the context of environmental and cultural trajectories of the Lower Ica Valley. Ph.D. thesis, Magdalene College, University of Cambridge, Cambridge
Beresford-Jones DG, Arce S, Oliver T, Whaley Q, Chepstow-Lusty A (2009) The role of Prosopis in ecological and landcape change in the Samaca basin, lower Ica Valley, South coast Peru from the Earlier Horizon to the Late Intermediate period. Latin Am Antiq 20(2):303–332
Boixados R (2002) Los pueblos de indios de La Rioja colonial. Tierra, trabajo y tributo en el siglo XVII. In: Farberman J, Gil Montero R (eds) Los pueblos de indios del Tucumán colonial: pervivencia y desestructuración. UNQ-Ediunju, Bernal, pp 15–58
Cáceres Freyre J (1962) Fabricación del patay en los algarrobales del Campo de Palcipas (La Rioja y Catamarca). University of La Plata, La Plata, Argentina, Serie Contribuciones 76:1–22
Capparelli A (1997) Reconstrucción ambiental de la instalación arqueológica Inka El Shincal. Unpublished Ph.D. thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata
Capparelli A (2007) El algarrobo blanco y negro—P. chilensis (Mol.) Stuntz y Prosopis flexuosa DC, Fabaceae—en la vida cotidiana de los habitantes del NOA: subproductos alimenticios. Kurtziana 33:1–19
Capparelli A (2008) Caracterización cuantitativa de productos intermedios y residuos derivados de alimentos del algarrobo (Prosopis flexuosa DC y P. chilensis (Mol.) Stuntz, fabaceae): aproximación experimental aplicada a restos arqueobotánicos desecados. Darwiniana 46(2):175–201
Capparelli A (2009) Intra-site comparison of the archaeoethnobotanical evidence of El Shincal: implicances to the Inka economy. In: La alimentación en la América precolombina y colonial: una aproximación interdisciplinaria, edited by the Department of Arcaheology and Anthropology of the Instituto Milà y Fontanals-Consejo Superior de Investigaciones Científicas (CSIC), España, Treballs d’Etnoarqueologia 7:113–144
Capparelli A, Lema V (2011) Recognition of post-harvest processing of algarrobo (Prosopis spp.) as food from two sites of Northwestern Argentina: an ethnobotanical and experimental approach for desiccated macroremains. Journal of Archaeological and Anthropological Sciences. doi:10.1007/s12520-011-0052-5
Capparelli A, Raffino RA (1997) Arqueoetnobotánica de El Shincal I: tallos finos, frutos y semillas. Tawantinsuyu 3:40–57
Capparelli A, Lema V, Giovannetti M (2004) El poder de las plantas. In: Raffino R (ed) El Shincal de Quimivil, Sarquís Editorial, Catamarca, Argentina. Chapter VII. pp 141–164
Capparelli A, Lema V, Giovannetti M, Raffino R (2005) The introduction of old world crops (wheat, barley and peach) in Andean Argentina during the 16th century A.D.: archaeobotanical and ethnohistorical evidence. Vegetation History and Archaeobotany 14:472–484
Capparelli A, Valamoti S, Wollstonecroft M (2010) Recent research in post-harvest traditions in human prehistory: Old and New World palaeoethnobotanical approaches to linking the archaeology and ethnobotany of plant processing. In: Pochettino ML, Ladio A, Arenas P (eds) Traditions and transformations in Ethnobotany, Cyted, S.S. de Jujuy. pp. 104
Capparelli A, Martínez A, Lema V (In Prep.) Algarrobo blanco y negro: diferenciación taxonómica en base a semillas y endocarpos.
D’Altroy T (2003) The Incas. Blackwell, Oxford
D’Antoni HL, Solbrig OT (1977) Algarrobos in South American cultures, past and present. In: Simpson BB (ed) Mesquite: its biology in two desert scrub ecosystems, Dowden. Hutchinson & Ross, Inc., Pennsylvania, pp 189–199
Dasso MC (2000) Celebración de la aloja: la preservación del fundamento mítico entre los Mataco-Wichí del Chaco central. Scripta Ethnologica 22:61–76
Farrington I (1999) El Shincal: un Cusco del Kollasuyu. In: Diez Marín C (ed) Proceedings of the 12th National Congress of Argentinean Archaeology, La Plata, 1997, vol 1. National University of La Plata Editorial, La Plata. pp 53–62
Giovannetti MA (2009) Articulación entre el sistema agrícola, el de irrigación, y las áreas de molienda como medida del grado de ocupación Inka en El Shincal de Quimivil-Los Colorados (Valle de Hualfín, Pcia. De Catamarca). Unpublished Ph.D. thesis, FCNyM, UNLP
Giovannetti MA, Lema V, Bartoli CG, Capparelli A (2008) Starch grains characterization of Prosopis chilensis (Mol.) Stuntz and P. flexuosa DC, and their implication in the analysis of archaeological remains in Andean South America. J Archaeol Sci 35:2973–2985
Hillman G (1984) Interpretation of archaeological plant remains: The application of ethnographic models from Turkey. In: Van Zeist W, Caspary WA (eds) Plants and Ancient Man. Studies in palaeoethnobotany. Biologisch-Archaeologisch Institute, State University, Groningen. AA Balkema, Rotterdam. pp 1–41
Jones G (1984) Interpretation of archaeological plant remains: Ethnographic models from Greece. In: Van Zeist W, Caspary WA (eds) Plants and Ancient Man. Studies in palaeoethnobotany. Biologisch-Archaeologisch Institute, State University, Groningen. AA Balkema, Rotterdam. pp 43–59
Kriskautzky N, Morales F (1999) La vivienda incaica en el sitio “Intihuatana de Yokavil”, Fuerte Quemado, Catamarca. In: Diez Marín C (ed.), Actas del XII Congreso Nacional de Arqueología Argentina, vol. I. pp 233–238.
Lema V, Giovannetti M, Deschamps C, Capparelli A, Raffino R (2009) Análisis de restos faunísticos en el sitio inkaico El Shincal (Catamarca, Argentina). Comparación con información arqueobotánica y análisis cerámico. In: La alimentación en la América precolombina y colonial: una aproximación interdisciplinaria, edited by the Department of Arcaheology and Anthropology of the Instituto Milà y Fontanals-Consejo Superior de Investigaciones Científicas (CSIC), España, Treballs d’Etnoarqueologia 7:97–112
López ML, Capparelli A, Nielsen AE (2011) Traditional post-harvest processing to make quinoa grains (Chenopodium quinoa var. quinoa) apt for consumption in Northern Lípez (Potosí, Bolivia): ethnoarqueological and archaeobotanical analyses. Journal of Archaeological and Anthropological Sciences. doi:10.1007/s12520-011-0060-5
Noli E (1999) La recolección en la economía de subsistencia de las poblaciones indígenas: una aproximación a través de fuentes coloniales (piedemonte y llanura tucumano- santiagueña, gobernacio’n del Tucumán). In: Aschero C, Korstanje A, Vuoto P (eds) En los tres reinos: prácticas de recolección en el cono sur de América. UNT, Tucumán, Argentina, pp 205–216
Orians GH, Solbrig OT (1977) In: Orians GH, Solbrig OT (eds) Convergent Evolution in warm deserts. An examination of strategies and patterns in deserts of Argentina and the United Status. Dowden, Hutchinson & Ross, Inc, Stroudsburg
Quiroga L (1999) Los dueños de los montes, aguadas y algarrobales: Contradicciones y conflictos coloniales en torno a los recursos silvestres. Un planteo del problema. In: Aschero C, Korstanje A, Vuoto P (eds) En los tres reinos: prácticas de recolección en el cono sur de América. UNT, Tucumán, Argentina. pp 217–226
Raffino RA (2004) El Shincal de Quimivil, Raffino RA (ed), Sarquís Editorial, Catamarca
Raffino RA, Iturriza RD, Gobbo D, Capparelli A, Deschamps C (2002) El Sinchiwasi de El Shincal de Quimivil. In: Academia Nacional de la Historia, Col. Investigaciones y Ensayos, Buenos Aires, Argentina, vol 52. pp 39–68
Raffino RA, Iturriza RD, Gobbo D, García Montes V, Capparelli A, Deschamps C (2004a) Kallanka. In: Raffino RA (ed) El Shincal de Quimivil, Sarquí Editorial, Catamarca, Argentina, Chapter IV. pp 91–106
Raffino RA, Iturriza RD, Gobbo D, Capparelli A, Moralejo R (2004b) Sinchiwasi. In: Raffino RA (ed) El Shincal de Quimivil, Sarquí Editorial, Catamarca, Argentina, Chapter V. pp 107–120
Raffino RA, Gobbo D, Iturriza RD, Capparelli A, Deschamps C (2004c). Ushno. In: Raffino RA (ed) El Shincal de Quimivil, Sarquí Editorial, Catamarca, Argentina, Chapter III. pp 69–90
Ratto N, Rodríguez MF, Hershey D (2007) Explotación y uso de recursos vegetales en sitios arqueológicos del área cordillerana y del valle mesotérmico (Dpto. Tinogasta, Catamarca, Argentina). Actas del XVII Congreso Nacional de Arqueología Chilena. Universidad Austral de Chile, Valdivia, Chile
Sempé de Gomez Llanes MC (1986) Mishma n7-Sitio incaico del valle de Abaucandto. Tinogasta-Catamarca. Revista del Museo de La Plata (Nueva Serie) 8:405–438
Simpson BB, Solbrig OT (1977) Introduction. In: Mesquite: its biology in two desert scrub ecosystems. Dowden, Hutchinson & Ross, Inc, Stroudsburg, pp 1–25
Snead J (1992) Imperial infraestructura and the Inka state storage system. In: Levine TY (ed) Inka storage system. University of Oklahoma, Norman, pp 62–106
Williams V (1985) Arqueología incaica en la región Centro-Oeste de Catamarca (República Argentina). Unpublished Ph.D. thesis, FCNyM, UNLP
Williams V, D’Altroy T (1998) El sur del Tawantinsuyu: un dominio selectivamente intensivo. Tawantinsuyu 5:170–178
Wollstonecroft M (2007) Post-harvest intensification in late pleistocene Southwest Asia: Plant food processing as a critical variable in epipalaeolithic subsistence and subsistence change. Unpublished Ph.D. thesis, Institute of Arcaheology, University College London
Yacovleff E, Herrera FL (1934) El mundo vegetal de los antiguos peruanos. Botánica Etnológica, Revista del Museo Nacional 3:243–322
Acknowledgements
The authors thank Soultana Valamoti, Ian Farrington, Dorian Fuller, and the anonymous referees who provided useful suggestions on the first versions of this article. Thanks is also given to R. Raffino, who gave me the possibility to be part of his archaeological team during these last 20 years.
Author information
Authors and Affiliations
Corresponding author
Additional information
Dedicated with affection to Gordon Hillman who introduced me to the world of food processing in archaeobotany more than 10 years ago
Rights and permissions
About this article
Cite this article
Capparelli, A. Elucidating post-harvest practices involved in the processing of algarrobo (Prosopis spp.) for food at El Shincal Inka site (Northwest Argentina): an experimental approach based on charred remains. Archaeol Anthropol Sci 3, 93–112 (2011). https://doi.org/10.1007/s12520-011-0061-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-011-0061-4