
Abstract
Changes in distribution of semi-evergreen forest in the Yucatán peninsula and variation of precipitation during the late Holocene were inferred using a geographical information system. Deterministic methods for spatial interpolation using fossil pollen data from seven cores elucidated environmental changes. The changes in the distribution of the semi-evergreen forest allowed to us infer variations in precipitation in the area and to distinguish whether changes of forest cover are a consequence of climate and/or of human activities. The reconstruction of the Preclassic period (at 550 and 50 b.c.) indicates higher precipitation than at present and suggests a more closed vegetation cover. Around a.d. 450 the vegetation acquired an open character indicating a reduction in precipitation. During this time the decrease in forest was not homogeneous in the Yucatán peninsula, indicating human impacts in certain areas. A reduction in forest cover but not a complete deforestation of the region is assessed during the Classic period (a.d. 450). The reconstruction of a.d. 950 shows the recovery of the forest and is related to the Medieval warm period. Geographical information systems are useful tools to reconstruct the spatial history of the vegetation of the Yucatán peninsula during the late Holocene.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Palaeoecological studies have been focussed on the reconstruction of vegetation distribution on different scales to understand the dynamics between vegetation and their main modifying agents: climate and human activities. To include a spatial analysis, first, knowledge is required about the current vegetation and its pollen signals that allow the interpretation of fossil pollen spectra. Secondly, gathering data from diverse locations is essential. Thirdly, the development of techniques and specialized tools for the analysis of data is needed.
From the Yucatán peninsula several palaeoecological studies are available that use fossil pollen (Carrillo-Bastos et al. 2010; Islebe and Sánchez-Sánchez 2002; Leyden et al. 1996, 1998; Torrescano-Valle and Islebe 2006; Torrescano-Valle 2007). Pollen rain studies (Correa-Metrio et al. 2011; Islebe et al. 2001; Torrescano-Valle 2007) as well as multiproxy studies allow the deciphering of climatic history through fossil pollen. However, techniques and specialized tools that allow us to understand data in a spatial context have not been applied.
Spatial reconstructions of other areas have used diverse methods. Some have followed Prentice (1985) modified by Sugita (1993), which allow identification of the source area where pollen originates and later is deposited in a water body. Most focussed on reconstructions generated by isopollen maps (Lenz and Riegel 2001; Paez et al. 2001; Ralska-Jasiewiczowa 1983) and isochrone maps (Birks 1989; Bernabo and Webb 1977). Others have mapped pollen percentages (Brubaker et al. 2005).
Recently, specialized software has become available to create geographical information systems (GIS) that allow the management of pollen data by means of spatial analysis and correlate the distribution of the archaeobotanical data with other spatial parameters (Gaudin et al. 2008). These methods work well to observe the change in distribution of types of vegetation (Flantua et al. 2007) and distribution of taxa (Giesecke and Bennett 2004; Wang et al. 2011). Although this technique is at an exploratory stage, it is quite useful and attractive for analyzing fossil pollen data.
The utility of GIS is to facilitate the interpretation of databases that are mostly large and complex, and which include data from different regions. Additionally, the information in different databases can be compared to carry out an interpretation of a complete area and not only of one location (Flantua et al. 2007; Wang et al. 2011) mainly as some research hypothesis can only be solved with a spatial focus. Such is the case of the determination of changes in the distribution of the vegetation that allow in an indirect way the reconstruction of the history of the climate of an area, as well as the detection of the impact of human activities.
The Yucatán peninsula was an important area of occupation by the ancient Maya civilization and supported a dense population during the Classic period (a.d. 300–900). The Mayan culture developed a vast agricultural system as a livelihood and was a major force in altering the environment and regional flora (Beach et al. 2006; Carozza et al. 2007; Coe 1993; Islebe et al. 1996; Leyden et al. 1998; Leyden 2002). The Mayan modified water bodies as reservoirs, transformed the local landscape by clearing large tracts of forest for construction and agriculture and increased the sedimentation rate in lakes (Anselmetti et al. 2007; Islebe et al. 1996; Leyden et al. 1998). The amount of deforestation is still unknown but the analysis of pollen records using GIS could help to decipher it. This may be possible because GIS has the ability to process multiple characteristics such as vegetation types through different geostatistical interpolation techniques. Having a number of records, these techniques allow the estimation of values in places or points that have not been measured and thereby form continuous thematic maps. In the case of palaeoecological studies it can be fossil pollen percentages that, prior to the calibration of current pollen rain, would represent an analogue of vegetation cover.
The distribution of vegetation on the Yucatán peninsula is governed mainly by precipitation levels (Sánchez-Sánchez and Islebe 2002). A clear example is given by the semi-evergreen forest taxa, which grow in areas with a precipitation between 1,000 and 1,500 mm/year (Sánchez-Sánchez and Islebe 2002) and occupy an area of ~62,027 km2 in the Yucatán peninsula. These requirements of precipitation make these taxa excellent indicators of climatic variations.
From the Yucatán peninsula it is known that the mosaic of current vegetation was formed during the early Holocene, which implies that the modern pattern of isohyets was also established then (Leyden 2002). However, any variations that isohyets have experienced in the past are unknown. Therefore, the present work has the objective to incorporate GIS into the analysis of fossil pollen, in order to analyze changes in the precipitation gradient of the Yucatán peninsula, by means of the reconstruction of the distribution of the semi-evergreen forest.
Study area
The Yucatán peninsula is a platform of calcareous rocks, with elevations of <400 m (Vidal-Zepeda 2005). Due to its intertropical position, low elevation, the strong maritime influence and high and uniform insolation along the whole year, the climate on the Yucatán peninsula is warm with slight fluctuations in the annual average (24–26°C) (Orellana et al. 1999). The interannual climate variability there is defined by differences in precipitation rather than temperature (Brenner et al. 2001). Rainfall in the area is highly variable spatially and temporally (Hodell et al. 2005a). As a result of prevailing trade winds, an east-west and south-north-gradient of descending precipitation exists (500–2,000 mm/year) (Hodell et al. 2005a; Miranda 1978; Orellana et al. 1999). The rainy season lasts from May to November, when the Intertropical Convergence Zone (ITCZ, usually located near the equator) and the Azores-Bermuda high-pressure system (centred in the mid-latitude North Atlantic) move northward (Hastenrath 1976) and the sea surface temperature is warm in the tropical/subtropical North Atlantic and Caribbean providing high moisture for precipitation (Brenner et al. 2001). In summer, the Yucatán peninsula frequently experiences tropical storms and hurricanes with much precipitation within a short time period (Brenner et al. 2001). Dry conditions prevail from December to April when the ITCZ and the Azores-Bermuda high-pressure zone migrate southward (Hastenrath 1976). During this time of relatively low sea surface temperature, a high-pressure gradient from the Azores to Bermuda and strong temperature inversion associated with enhanced trade winds produce low precipitation on the Yucatán peninsula (Brenner et al. 2001).
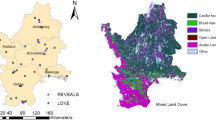
The peninsula has semi-evergreen forest, tropical subdeciduous forest, tropical deciduous forest and several wetland types (Fig. 1; Miranda 1958). Characteristic species of semi-evergreen forest are Manilkara zapota, Vitex gaumeri, Myrcianthes fragrans, Talisia olivaeformis, Swartzia cubensis, Sabal japa, Brosimum aliscastrum, Sideroxylon foetidissimum, Pouteria campechiana, Swietenia macrophylla, Gymnanthes lucida, Coccoloba diversifolia, Drypetes lateriflora, Thrinax radiata and Coccothrinax readii (Sánchez-Sánchez 1987; Duran and Olmsted 1999). Characteristic species of tropical subdeciduous and deciduous forest are Acacia gaumeri, Acacia pennatula, Mimosa bahamensis, Havardia albicans, Gymnopodium floribundum, Metopium brownei and Bursera simaruba. For wetland some species are Cladium jamaicensis, Typha dominguensis or Thalia geniculata, Haematoxylum campechianum, Bucida buceras, Dalbergia glabra, Rhizophora mangle, Avicennia germinans, Laguncularia racemosa and Conocarpus erectus (Duran and Olmsted 1999). The semi-evergreen forest, which is used in this study as an indicative vegetation type, is the most dominant vegetation type of the Yucatán peninsula (Sánchez-Sánchez and Islebe 2002). 25–50% of the taxa lose their leaves during the dry season (Durán and Olmsted 1999) and trees have canopy heights between 15 and 25 m (Sánchez-Sánchez 1987).

Study area that includes the location of sites of fossil pollen analysis used to reconstruct spatial and vegetation types of the Yucatán peninsula
Methods
The reconstruction of the distribution of the semi-evergreen forest was carried out with ArcGis 9.3 software. Four thematic maps for the last 2,500 years were elaborated which present the percentages of this forest type, at intervals of 500 years. The reconstructed interval corresponds to most of the palaeoecological records of the peninsula. The thematic maps were elaborated using interpolation tools and extrapolation, starting from a database that contained the information from seven cores based on previous works in the Yucatán peninsula (Fig. 1; Table 1): Cenote San José Chulchacá (Leyden et al. 1996), Lake Cobá (Leyden et al. 1998), Puerto Morelos mangroves (Islebe and Sánchez-Sánchez 2002), Puerto Morelos mangroves II (Torrescano-Valle 2007), Lake Tzib I (Torrescano-Valle 2007), Lake Tzib II (Carrillo-Bastos et al. 2010) and Silvituc (Torrescano-Valle 2007). These seven sites were selected because their spatial distribution represents the different regions of the Yucatán peninsula and allows reconstruction of the vegetation in most of this area.
Chronology
To obtain the chronology, linear regressions were performed using the original calibrated ages presented by the authors of each record. Prior to the calculation of the regressions, all dates were recalibrated in order to determine if there were differences with the original calibrations of the authors. However, the differences were minimal and considering the time resolution of the records (Table 1), it was decided to use the calibrated ages provided by the authors. Because the temporal resolution of the data varies between 100 and 400 years, an interval of 500 years for the preparation of maps was established. The oldest map corresponds to the date of about 550 b.c. because the youngest record of one core is 600 b.c. (Table 1). From this age (600 b.c.) a 500 year interval was applied and the ages of the three remaining maps were established.
The number of radiocarbon samples in the seven records is variable. Three of the sites, Lake Coba, Cenote San Jose Chulchaca and Silvituc have six, five and six dates respectively, which allows a reliable age modelling. However, obtaining dating from Lake Tzib (cores I and cores II) only with two dates, has been problematic, given the nature of the sediments, which have a low organic matter content (Carrillo-Bastos et al. 2010). However, the correlation of this record with others obtained from the Yucatán peninsula (Leyden et al. 1998; Hodell et al. 1995, 2007), Guatemala (Islebe et al. 1996; Leyden et al. 1993; Mueller et al. 2010) and elsewhere (Haug et al. 2001), have allowed us to validate the estimated ages for this record.
Setting up the database
The database was built selecting indicator taxa of the semi-evergreen forest according to pollen rain studies developed in the region (Islebe et al. 2001; Torrescano-Valle 2007). An indicator taxon is consistently recorded in the pollen signal of a certain type of vegetation, in this case semi-evergreen forest. Such taxa are: Anacardiaceae, Borreria verticillata, Brosimum alicastrum, Bursera sp., Burseraceae, Euphorbiaceae, Ficus sp., Guettarda combsii, Fabaceae, Moraceae, Myrtaceae, Pouteria sp., Rubiaceae, Sapium and Sapotaceae. Once these taxa were identified and the abundance of each taxon in each of the intervals calculated (Fig. 2), maps were elaborated for ~550 b.c., ~50 b.c., ~a.d. 450, and ~a.d. 950. Later, these abundances were added to obtain the total value from each place at the different dates. Since the program requires at least ten points to carry out the analysis, a similar strategy was used by Flantua et al. (2007) and three points were added to the database. These points were placed near to Lake Cobá, Cenote San José Chulchacá and Lake Silvituc, and they contained the same percentage values as the nearby sites. In the location of these additional sites the environmental conditions were the same. Therefore no environmental stochasticity was added.

Summary diagrams of semi-evergreen forest pollen taxa from five sites. Horizontal lines indicate the ages for which maps are constructed
Development of vegetation layers
The mapping of the vegetation was established with the local polynomial interpolation technique (ILP), provided by the extension geostatistical analyst within ArcGis 9.3. The ILP is a deterministic method that, in the same way as other ILPs, uses values measured at different points inside an area to create a continuous surface (Wadsworth and Treweek 1999). In this study, the local interpolation for polynomials was chosen because it does not imply properties of the measured data, such as normal distribution, and gave smaller prediction errors than the rest of the deterministic interpolators. After several attempts, the prediction was carried out with a power function of (p) = 2 and under the option of search for the ideal influence for each distance (IDD). These parameters are those that produced surfaces with a smaller prediction error. In IDD, the program carries out a cross-validation of the measured values (ESRI 2001). As the name indicates, the technique predicts values inside the polygon that is formed from the measured points. After the interpolation it was necessary to carry out an extrapolation for the rest of the peninsula.
Once the palaeovegetation maps were obtained for the four ages, a thematic layer that shows the contour of the current distribution of the semi-evergreen forest was incorporated. This line was added in order to detect changes in distribution. Later, with the objective of detecting changes of precipitation, a third layer was added that presents the modern distribution of the isohyets.
Results
As a result of the interpolation and extrapolation of the percentages of the seven sites (Table 1), four maps were obtained (Fig. 3), which correspond to the calibrated ages of ~550 b.c., ~50 b.c., ~a.d. 450 and ~a.d. 950. These maps show the prediction of the pollen percentages of taxa of semi-evergreen forest, the contour of the current distribution of medium forest, and the modern distribution of the isohyets. The percentages on the maps indicate the cover of arboreal vegetation. The high values, in blue, indicate a greater amount of cover and therefore a more closed arboreal vegetation type. In accordance with the values reported from modern pollen rain studies (Islebe et al. 2001; Torrescano-Valle 2007), the minimum value that indicates the presence of semi-evergreen forest is 30%. Below this threshold, the vegetation is of open character.

Maps corresponding to the ages of A 550 cal. b.c., B 50 cal. b.c., C a.d. 450 and D a.d. 950 as a result of interpolation and extrapolation
In the reconstruction corresponding to ~550 b.c. (Fig. 3A) the highest percentages (85–92%) are observed in the south and central peninsula, while in the northern part the lowest percentages are found (2.4–8%). A tendency to more reduced percentages, although the values are not lower than 30%, is observed in an east–west direction. In the western part the highest percentages correspond to values between 45 and 55%.
In the map of ~50 b.c. (Fig. 3B), the areas of high percentages are more broadly extended. The limit of semi-evergreen forest extends as much in the north as in the south of the peninsula. The area of low percentage (2.4–8%) decreases. The areas of 45–51% and of 51–56%, that were previously restricted to the centre of the peninsula, extend to the western coast.
In the prediction of ~a.d. 450 (Fig. 3C) a clear reduction of the percentages of elements of semi-evergreen forest is observed. The highest values range from 56 to 62%, and are restricted to the east coast of the peninsula. Although the limit of the semi-evergreen forest does not reduce, the area of 45–51% earlier distributed in the central part decreases and is replaced with an area of 40–45%, which indicates the presence of a more open forest type.
The map corresponding to ~a.d. 950 (Fig. 3D) reflects the return of high percentages to the east portion (85–92%) and the central part of the peninsula (51–62%). The limits of the forest (<30%) in the north as well as in the south retreat to a similar distribution as during the ~550 b.c. time frame. In the central part, the distribution pattern is similar to that of ~550 b.c., but with lower values (34–40%).
Discussion
Interpolation tools and spatial extrapolation, used for the analysis of palaeovegetation, allow the mapping of continuous areas of particular percentages of fossil pollen, in areas where no data from cores are available. Although these predictions do not show the exact past distribution of the semi-evergreen forest like the present-day pattern, they exhibit tendencies that allow us to recognize the limits of the past distribution. This is a function of the minimum percentage that the sum of elements obtained in the current pollen rain. The percentages also provide an estimate of the vegetation cover and the amount of arboreal vegetation, and therefore tree cover.
Based on these arguments, it is possible to detect the changes in the distribution of the vegetation and to infer precipitation variations in the Yucatán peninsula for the reconstructed ages. In the four maps the tendency of pollen percentages to increase is seen going from east to west. This is similar to the modern distribution of the isohyets where the precipitation increases towards the eastern coast. This precipitation gradient, as mentioned above, corresponds to the trade winds, which blow from east to the west (Hodell et al. 2005a). These winds carry humidity and they discharge precipitation over the Yucatán peninsula, the largest quantity of which falls near the eastern coast. Due to this gradient, at present, the arrangement of the vegetation is distributed in a similar way; on the eastern fringe it is semi-evergreen forest, which requires higher precipitation (>1,100 mm/year), in the central fringe tropical subdeciduous (>800 mm/year) forest and in the western part tropical deciduous forest (500–1,200 mm/year).
By comparing the established maps it is possible to observe the contraction and expansion of the forest. During the Preclassic (~550 and ~50 b.c.), the map colours show a greater cover than during the Classic (~a.d. 450) and Postclassic periods (~a.d. 950). These changes coincide with variations in the precipitation that have been reported in other studies, where proxies other than pollen were used (Carrillo-Bastos et al. 2010; Curtis et al. 1996; Haug et al. 2001, 2003; Hodell et al. 2005a, b, 2007).
At ~550 b.c., the position of the area of 40–45% extending along the isohyets from 600 to 800 mm precipitation, probably indicates that more humidity was available then, than at present. This is inferred because at that time the semi-evergreen forest is distributed in areas with a minimum precipitation of 1,000 mm at present.
Hodell et al. (2007) and Carrillo-Bastos et al. (2010) report low values of δ18O, which suggests conditions of abundant precipitation. Hodell et al. (2005b) presented low values of density of the sediments of Lake Chichancanab, suggesting wet conditions. Additionally from the Cariaco basin in Venezuela, Haug et al. (2001) reported high values of titanium, which indicate abundant material of terrigenous origin being washed towards the sea due to high precipitation. Hodell et al. (2007) and Haug et al. (2001) reveal with different proxies that precipitation of ~550 b.c. was higher than at present. This fits with the reconstruction of the semi-evergreen forest, as forests extend to areas with present-day isohyets <800 mm. Curtis et al. (1996) reported low values of δ18O, however these are not lower than present values.
The reconstruction of ~50 b.c. shows expansion of the semi-evergreen forest, and dominance of arboreal vegetation. The isohyets of 600–800 mm coincide with percentages of 56 and 62% of forest elements, indicating that at this time the precipitation along the northeast coast of the of the peninsula was higher compared to ~550 b.c. When observing the values of other proxies, they agree with conditions of abundant precipitation as at Cariaco, where high percentages of titanium were detected around this time (Haug et al. 2003). Hodell et al. (2005b) and Curtis et al. (1996) also reported an increase in precipitation at this date. However, the magnitude of change is not as large as reported by Haug et al. (2001).
The map corresponding to ~a.d. 450 compared with the Preclassic, registers a clear reduction in forest cover. This reduction could be due to the decrease in the precipitation. Haug et al. (2003) detected in this period a reduction in the titanium percentages, indicating less precipitation than ~50 b.c. However, the values registered in the Cariaco basin (Haug et al. 2003), compared with values during the Terminal Classic drought were much lower. Neither was this stronger than the pre-abandonment drought (Haug et al. 2003). Hodell et al. (2005b) do not report a significant reduction in precipitation, as the density values are only slightly higher than those found in the previous age. However, in the core of Punta Laguna, a severe drought is observed (Curtis et al. 1996).
The reduction in vegetation cover may be related to errors in the interpolation, however the interpolation used for the four reconstructions was the same, so that the changes observed between different maps can be comparable.
Another cause of the decrease of the vegetation cover is due to the impact of the activities of the Mayan culture people on the vegetation. Areas like Cobá were already developed: their urbanization began during the Early Classic (Folan 1983). Leyden et al. (1998) suggest that the lake was modified to control the availability of water. This impact can also be explained by the fact that in the eastern part of the peninsula the reduction in tree cover is larger than on the western coast. Although a decrease of precipitation is suggested, the decrease was not of the same magnitude than in the eastern part of the peninsula.
Another important aspect of this reconstruction is related to the use of resources by the people of the Mayan culture. In several publications it is pointed out that this culture made extensive use of natural resources including deforestation (Abrams and Rue 1998; Dunning et al. 2002; Leyden 2002; Turner et al. 2003). However, in this map it is observed that at least in the ~a.d. 450 time frame, most of the peninsula forests remained and a massive deforestation theory is not supported.
The reconstruction of the ~a.d. 950 map falls in the period of the Medieval climatic optimum and it reflects the recovery of the forest. However, this climatic event has not been corroborated for the entire peninsula, but mainly for those analyses with high temporal resolution (Curtis et al. 1996; Hodell et al. 1995, 2005a; Mueller et al. 2010), and still lacks detailed records of its expression in the region. While analyzing all sites it is possible to detect that, at least for this time frame, the conditions were of more humidity than at ~a.d. 450. This is a period of relative humidity reported by Haug et al. (2003), but during the terminal Classic period a second drought was reported by Hodell et al. (1995). The distribution of the percentages is very similar to that of the ~550 b.c. map. However, in the western portion and central peninsula a slight reduction in tree cover is observed, compared with that of ~550 b.c. When observing titanium values in the Cariaco core, these are high but less than the values reported for ~50 b.c. (Haug et al. 2003). In the Lake Chichancanab record (Hodell et al. 2005b), this date falls into a wet period between two droughts, and the value of density is similar to the ~550 b.c. map. From Punta Laguna the curve shows an increase in precipitation compared to ~550 b.c., while the data of Cytheridella ilosvayi show more arid conditions (Curtis et al. 1996). The latter curve is consistent with the findings of Medina-Elizalde et al. (2010) who reported a drought for this period. However it is not possible to know if the rest of the reconstructions agree or not, because the curve does not go beyond ~a.d. 500.
Also at this time, the Mayan culture people continued inhabiting and modifying the vegetation in some regions of the peninsula, which is reflected in less arboreal vegetation and the presence of agricultural indicators like Zea mays.
Conclusion
With the incorporation of GIS to available palaeoecological information, it was possible to integrate, to process, to project and to analyze the information from diverse sources and to reconstruct the spatial history of the semi-evergreen forest. This reconstruction allowed detection of the variation in precipitation and in qualitative terms to estimate its change. With the spatial perspective it is possible to detect the simultaneous effect of climate and society, through analyzing the change of the different areas of Yucatán peninsula. Changes in vegetation before and during the Preclassic period in the peninsula have been mainly caused by climate. During this period, conditions were wetter than at present. During the Classic the climatic factor is added with activities carried out by the Mayan culture people. At ~a.d. 450 the simultaneous pressure of both factors is observed on the forest cover. In spite of this pressure the vegetation kept an important amount of cover, showing that the use of the resources did not result in total deforestation. Around ~a.d. 950 vegetation recovered related to an increase of precipitation. This increase represents the Medieval warm period in the peninsula.
This is the first approach to integrate GIS in the reconstruction of the palaeovegetation of the peninsula of Yucatán and even in Mexico, and its potential use is revealed. However, the use of this tool is at an early stage and it is necessary to continue in the development of two main research lines: (1) to increase the number of high temporal resolution pollen records (cores) from sites which have not been sampled, that would allow the use of more complex models (with geostatistic bases) to carry out better spatial reconstructions. (2) To improve current models that relate the vegetation with environmental factors, mainly climate, so that they allow quantitative determination of the climatic variations.
References
Abrams EM, Rue DJ (1998) The causes and consequences of deforestation among the prehistoric Maya. Human Ecol 16:377–395
Anselmetti FS, Ariztegui D, Brenner M, Hodell DA, Rosenmeier MF (2007) Quantification of soil erosion rates related to ancient Maya deforestation. Geology 35:915–918
Beach T, Dunning N, Luzzadder-Beach S, Cook D, Lohs J (2006) Impacts of the ancient Maya on soil erosion rates in the central Maya lowlands. Catena 65:166–178
Bernabo JC, Webb T (1977) Changing patterns in the Holocene pollen record of northeastern North America: a mapped summary. Quat Res 8:64–96
Birks HJB (1989) Holocene isochrone maps and patterns of tree-spreading in the British Isles. J Biogeogr 16:503–540
Brenner M, Hodell DA, Curtis JH, Rosenmeier M, Abbott MB (2001) Abrupt climate change and Pre-Columbian cultural collapse. In: Markgraf V (ed) Interhemispheric climatic linkages. Academic Press, San Diego, pp 87–103
Brubaker LB, Anderson PM, Edwards ME, Lozhkin AV (2005) Beringia as a glacial refugium for boreal trees and shrubs: new perspectives from mapped pollen data. J Biogeogr 32:833–848
Carozza JD, Galop D, Metailie JP, Vanniere B, Bossuet F, Monna JA, Lopez-Saez MC, Arnauld V, Breuil M, Forne M, Lemonnier E (2007) Landuse and soil degradation in the southern Maya lowlands from Pre-Classic to Post-Classic times: the case of La Joyanca (Peten, Guatemala). Geo Acta 20:195–2007
Carrillo-Bastos A, Islebe GA, Torrescano-Valle N, González NE (2010) Holocene vegetation and climate history of central Quintana Roo, Yucatán Península, Mexico. Rev Palaeobot Palynol 160:189–196
Coe MD (1993) The Maya. Thames and Hudson, London
Correa-Metrio A, Bush MB, Pérez L, Schwalb A, Cabrera KR (2011) Pollen distribution along climatic and biogeographic gradients in northern Central America. Holocene 21:687–698
Curtis JH, Hodell DA, Brenner M (1996) Climate variability on the Yucatán Península (Mexico) during the past 3500 years, and implications for Maya cultural evolution. Quat Res 46:37–47
Dunning NP, Luzzadder-Beach S, Beach T, Jones JG, Scarborough V, Culbert TP (2002) Arising from the Bajos: the evolution of a Neotropical landscape and the rise of Maya civilization. Ann Assoc Am Geogr 92:267–283
Duran R, Olmsted I (1999) Vegetación de la península de Yucatán. In: Córdoba J (ed) García de Fuentes A. Atlas de procesos territoriales de Yucatán. UADY, Mérida, pp 163–182
ESRI (2001) Using ArcGis geostatistical analyst. ESRI, Redlands
Flantua S, Van Boxel J, Hooghiemstra H, Van Smaalen J (2007) Application of GIS and logistic regression to fossil pollen data in modelling present and past spatial distribution of the Colombian savanna. Clim Dyn 29:697–712
Folan WJ (1983) Summary and conclusions. In: Folan WJ, Kintz ER, Fletcher LA (eds) Cobá, a classic Maya metropolis. Academic Press, New York, pp 211–217
Gaudin L, Marguerie D, Lanos P (2008) Correlation between spatial distributions of pollen data, archaeological records and physical parameters from north-western France: a GIS and numerical analysis approach. Veget Hist Archaeobot 17:585–595
Giesecke T, Bennett KD (2004) The Holocene spread of Picea abies (L.) Karst in Fennoscandia and adjacent areas. J Biogeogr 31:1,523–1,548
Hastenrath S (1976) Variations in low-latitude circulation an extreme climatic events in the Tropical Americas. J Atmos Sci 33:202–215
Haug GH, Hughen KA, Sigman DM, Peterson LC, Rohl U (2001) Southward migration of the intertropical convergence zone through the holocene. Science 293:1,304–1,308
Haug GH, Gunther D, Peterson LC, Sigman DM, Hughen KA, Aeschlimann B (2003) Climate and the collapse of Maya civilization. Science 299:1,731–1,735
Hodell DA, Curtis JH, Brenner M (1995) Possible role of climate in the collapse of classic Maya civilization. Nature 375:391–394
Hodell DA, Brenner M, Curtis JH (2005a) Climate change on the Yucatán Península during the Little Ice Age. Quat Res 63:109–121
Hodell DA, Brenner M, Curtis JH (2005b) Terminal Classic drought in the northern Maya lowlands inferred from multiple sediment cores in Lake Chichancanab (Mexico). Quat Sci Rev 24:1,413–1,427
Hodell DA, Brenner M, Curtis JH (2007) Climate and cultural history of the Northeastern Yucatan Península, Quintana Roo, Mexico. Clim Chang 83:215–240
Islebe GA, Sánchez-Sánchez O (2002) History of late holocene vegetation at Quintana Roo, Caribbean coast of Mexico. Plant Ecol 160:187–192
Islebe GA, Hooghiemstra H, Brenner M, Curtis JH, Hodell DA (1996) A holocene vegetation history from lowland Guatemala. Holocene 6:265–271
Islebe GA, Villanueva-Gutierrez R, Sánchez-Sánchez O (2001) Relación lluvia de polen-vegetación en selvas de Quintana Roo. Bol Soc Bot México 69:31–38
Lenz O, Riegel W (2001) Isopollen maps as a tool for the reconstruction of a coastal swamp from the Middle Eocene at Helmstedt (Northern Germany). Facies 45:177–194
Leyden BW (2002) Pollen evidence for climatic variability and cultural disturbance in the Maya lowlands. Anc Mesoam 13:85–101
Leyden BW, Brenner M, Hodell DA, Curtis JH (1993) Late Pleistocene climate in the Central American lowlands. Geophys Mon Ser 78:165–178
Leyden BW, Brenner M, Whitmore T, Curtis JH, Piperno D, Dahlin BH (1996) A record of long- and short-term climatic variation from northwest Yucatán: Cenote San José Chulchacá. In: Fedick SL (ed) The managed mosaic: ancient Maya agriculture and resource use. University of Utah Press, Salt Lake City, pp 30–50
Leyden BW, Brenner M, Dahlin BH (1998) Cultural and climatic history of Cobá Lowland Maya city in Quintana Roo, Mexico. Quat Res 49:111–122
Medina-Elizalde M, Burns SJ, Lea DW, Asmerom Y, Von Gunten L, Polyak V, Vuille M, Karmalkar A (2010) Stalagmite climate record from the Yucatán Peninsula spanning the Maya terminal classic period. Earth Planet Sci Lett 298:255–262
Miranda F (1958) Estudios acerca de la vegetación. In: Beltran E (ed) Los recursos naturales del sureste y su aprovechamiento. Ediciones del Instituto Mexicano de Recursos Naturales Renovables, México, pp 215–271
Miranda F (1978) Vegetación de la Península de Yucatán: rasgos fisiográficos. Colegio de posgraduados, Chapingo
Mueller AD, Islebe GA, Anselmetti FS, Ariztegui D, Brenner M, Hodell DA, Hajdas I, Hamann Y, Haug GH, Kennett DJ (2010) Recovery of the forest ecosystem in the tropical lowlands of northern Guatemala after disintegration of Classic Maya polities. Geology 38:523–526
Orellana LR, Balam K, Bañuelos RI, García de Miranda E, González-Iturbide AJ, Herrera CF, Vidal J (1999) Evaluación climática. In: Córdoba J (ed) García de Fuentes A. Atlas de procesos territoriales de Yucatán. UADY, Mérida, pp 163–182
Paez MM, Schäbitz F, Stutz S (2001) Modern pollen-vegetation and isopoll maps in southern Argentina. J Biogeogr 28:997–1021
Prentice C (1985) Pollen representation, source area and basin size: toward a unified theory of pollen analysis. Quat Res 23:76–78
Ralska-Jasiewiczowa M (1983) Isopollen maps for Poland: 0–11000 years B.P. N Phytol 94:133–175
Sánchez-Sánchez O (1987) Estructura y composición de la selva mediana superennifolia presente en el Jardín Botánico del CIQRO, Puerto Morelos. Universidad Veracruzana, Xalapa
Sánchez-Sánchez O, Islebe GA (2002) Tropical forest communities in southeastern Mexico. Plant Ecol 158:183–200
Sugita S (1993) A model of pollen source area for an entire lake surface. Quat Res 39:239–244
Torrescano-Valle N (2007) Reconstrucción paleoambiental del Holoceno Medio-Tardío en la parte centro-sur de la península de Yucatán, México. Doctoral Thesis, El Colegio de la Frontera Sur
Torrescano-Valle N, Islebe G (2006) Tropical forest and mangrove history from southeastern Mexico: a 5000 yr pollen record and implications for sea level rise. Veget Hist Archaeobot 15:191–195
Turner BL II, Klepeis P, Schneider LC (2003) Three millennia in the southern Yucatán peninsular region: implications for occupancy, use and “carrying capacity”. In: Gómez-Pompa A, Allen M, Fedick SL, Jiménez-Osornio J (eds) The lowland Maya area: three millennia at the human-wildland interface. Halworth Press, New York, pp 361–387
Vidal-Zepeda R (2005) Las regiones climáticas de México. Instituto de Geografía, UNAM, Mexico
Wadsworth R, Treweek J (1999) Geographical information systems for ecology: an introduction. Longman, London
Wang LC, Wu JT, Lee TQ, Lee PF, Chen SH (2011) Climate changes inferred from integrated multi-site pollen data in northern Taiwan. J Asian Earth Sci 40:1,164–1,170
Acknowledgments
Conacyt is acknowledged for funding the projects “Reconstrucciones paleoclimáticas y paleoecológicas del Holoceno tardío de Quintana Roo” and “Variabilidad climática y paleoecología de los últimos 1500 años del sureste de México”.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by E. C. Grimm.
Rights and permissions
About this article
Cite this article
Carrillo-Bastos, A., Islebe, G.A. & Torrescano-Valle, N. Geospatial analysis of pollen records from the Yucatán peninsula, Mexico. Veget Hist Archaeobot 21, 429–437 (2012). https://doi.org/10.1007/s00334-012-0355-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-012-0355-1