
Abstract
Arctic Cod (Boreogadus saida) occur throughout the circumpolar north; however, their distributions at localized scales are not well understood. The seasonal habitat associations and diet preferences across life-history stages of this keystone species are also poorly known, thereby impeding effective regulatory efforts in support of conservation objectives. The distribution of Arctic Cod in the Canadian Beaufort Sea was assessed using bottom trawling in shelf and slope habitats between 20 and 1000 m depths. Highest catch biomasses occurred at 350 and 500 m depth slope stations, coinciding with >0 °C temperatures in the Pacific–Atlantic thermohalocline and Atlantic water mass. Calanus glacialis, Calanus hyperboreus, Themisto libellula, and Themisto abyssorum were identified as key prey species in the diet of Arctic Cod, comprising approximately 86 % of total biomass in guts. Hierarchical cluster analysis with a SIMPROF test identified five statistically significant (p < 0.05) diet groups among gut samples. Arctic Cod shifted from a primarily Calanus diet at shelf stations (<200 m depth) to a Themisto diet in slope habitats (>200 m depth) coinciding with an associated increase in fish standard length with depth. Smaller Arctic Cod fed primarily on Calanus copepods and larger Arctic Cod fed primarily on the larger Themisto species. The habitat and diet associations presented here will inform knowledge of structural and functional relationships in Arctic marine ecosystems, aid in mitigation and conservation efforts, and will enhance our ability to predict the effects of climate change on the local spatial and depth associations of this pivotal marine fish.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Arctic (or Polar) Cod (Boreogadus saida), as a keystone species occupying a central trophic position in Arctic marine ecosystems, is considered to be a significant consumer of zooplankton and is a key prey for marine mammals and birds (Bradstreet et al. 1986; Welch et al. 1993). Arctic Cod occurs throughout the circumpolar north occupying a diverse suite of habitats throughout their geographic range from nearshore estuarine areas to deep-water marine environments (Andriyashev et al. 1980; Craig et al. 1982; Moulton and Tarbox 1987; Gradinger and Bluhm 2004; Majewski et al. 2013; Drost et al. 2014). Distributions at localized scales, however, are poorly understood. Habitat associations and diet preferences across life-history stages are also poorly known. Given the significance of the species in Arctic ecosystems, these knowledge gaps impede oceans management objectives.
Past research on the life-history and habitat associations of Arctic Cod in the Canadian Beaufort Sea has primarily focused on the timing of cohort hatching and the distribution of young-of-year in relation to the break-up of sea ice and water temperature (Fortier et al. 2006; Bouchard and Fortier 2008) and on winter under-ice habitat use in Franklin Bay (Amundsen Gulf) (Benoit et al. 2008, 2010, 2014; Geoffroy et al. 2011). While often considered an ice-associated species (Lønne and Gulliksen 1989; Gradinger and Bluhm 2004; Fortier et al. 2006), hydroacoustic surveys have observed Arctic Cod in near-bottom aggregations (Benoit et al. 2008; Geoffroy et al. in press), indicating that at least some portion of the population lives a benthopelagic lifestyle. Crawford et al. (2012) observed high biomass of Arctic Cod in the Atlantic water mass, generally between 250 and 350 m depth, along the continental slope in the Chukchi and Beaufort seas and near bottom in Barrow Canyon and Mackenzie Trough. Recent bottom trawling surveys indicate that Arctic Cod is numerically dominant in bottom habitats throughout the Beaufort Sea, representing 92 % of fish captured during a multispecies survey of fishes and habitats in shelf and slope waters ranging from 40 to 470 m depths in the western US Beaufort Sea (Logerwell et al. 2011; Rand and Logerwell 2011; Rand et al. 2013). Similarly, in the Canadian Beaufort Sea, Arctic Cod was the most abundant species in beam trawl catches in nearshore shelf and coastal areas (Lowdon et al. 2011; Majewski et al. 2009a, b, 2011). Rand et al. (2013) reported that the diets of demersal Arctic Cod differed from those captured in pelagic nets, indicating differences in habitat usage and feeding strategies within the same geographic area. The generality of differential habitat usage within an area, however, is poorly understood.
Within the Canadian Beaufort Sea, the diversity, distributions, and ecological roles of offshore marine fishes have not been studied comprehensively. The Beaufort Regional Environmental Assessment (BREA) Offshore Marine Fishes Project (2011–2015) was designed to address gaps in baseline ecological knowledge regarding key marine fishes to support regulatory processes regarding hydrocarbon development and conservation efforts in the offshore Canadian Beaufort Sea. Because of their central role in Arctic marine food webs, impacts to Arctic Cod resulting from industrial development and climate change could have both upward and downward cascading trophic effects. As such, it is imperative to understand the habitat and diet associations of Arctic Cod in order to effectively predict and mitigate potential impacts at both local and regional scales. During August 2012, the distribution of Arctic Cod was assessed using bottom trawling in shelf and slope habitats between 20 and 1000 m depths. Herein, we examine (1) the relative biomass of Arctic Cod within and across depth strata, (2) associations among catch biomasses of Arctic Cod with shelf and slope bottom habitats and water masses, and (3) the diet of Arctic Cod within and across bottom and water mass habitats. Results presented here will inform knowledge of structural and functional relationships in the Beaufort Sea ecosystem, enhance our ability to predict the effects of industrial development and climate change, and aid in mitigation and conservation efforts for this pivotal marine fish species.
Materials and methods
Description of study area
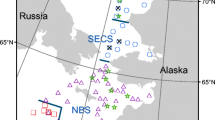
This study took place in the Canadian Beaufort Sea in August 2012 between the Yukon–Alaska border (141.00°W) and the Baillie Islands, Northwest Territories (128.20°W) (Fig. 1). Complex spatial and depth heterogeneity in the Canadian Beaufort Sea results in substantive variability in habitat structure and characteristics. Temperature, salinity, and chemical characteristics can be used to delineate the various water masses (by depth) which provide the structure of habitats available to fishes and other biota. Sea-ice melt, freshwater input from the Mackenzie River, incoming surface flows along the coast from the North Pacific Ocean via Bering Strait, subsurface polar gyres originating from the Atlantic Ocean, and deep Arctic basin water all contribute to the water mass structure of the Canadian Beaufort Sea. Variability in temperature and salinity among these inputs results in buoyancy differences, thereby creating a highly structured system consisting of different water masses as layers by depth. These layers provide distinct habitats within which particular types of fishes and other biota may associate. Herein, we consider the association of Arctic Cod within a simplified representation of the vertical water mass structure of the Canadian Beaufort Sea as generally described in McLaughlin et al. (1996, 2005) and Hodgson et al. (2015). A surface layer of seasonal freshwater inputs from the Mackenzie River and sea-ice melt is subject to wind-driven mixing in the summer to approximately 50 m depth and is herein referred to as the Polar Mixed Layer. Beneath this layer lies a cold halocline of Pacific origin waters (i.e., Pacific water mass) with temperatures ranging between 0 and −1 °C or colder and salinities ranging from 30 to 33.5 PSU. The maximum depth of the Pacific water mass (~200 m) coincides with the outer extent of the continental shelf, where a thermohalocline marks the transition from Pacific waters to warmer but more saline Atlantic water (i.e., Atlantic water mass; up to 1.0 °C, 33.5–34.95 PSU) coinciding with bottom habitats on the upper continental slope. The core Atlantic water mass is found at approximately 400 m depth and extends down the lower slope to roughly 800 m and overlies the colder, highly saline waters of the Arctic water mass (≥900 m depth) with salinity <34.9 PSU and temperatures <0 °C. A representation of the vertical water mass structure at the easternmost and westernmost transects of the study (Fig. 1) is presented in Fig. 2, with labels to indicate the simplified vertical water mass structure described above.

Transects and fishing stations on the Canadian Beaufort Shelf and slope during August 2012

Sampling stations along the TBS and DAL transects in relation to vertical profiles of temperature and salinity. Sampling stations are marked with black triangles. Water mass structure is indicated as generally described in McLaughlin et al. (1996, 2005) and Hodgson et al. (2015), with an upper polar mixed layer reflecting seasonal freshwater inputs, an underlying cold halocline of Pacific origin waters (herein called the Pacific water mass), and a thermohalocline beginning at approximately 200 m depth marking the transition from Pacific waters to warmer more saline Atlantic water (i.e., Atlantic water mass). The core Atlantic water mass is found at approximately 400 m depth and extends to approximately 800 m and overlies colder waters (herein called the Arctic water mass)
Field collections and sample processing
Benthic trawling was conducted from the F/V Frosti along four transects (TBS, GRY, KUG, and DAL; Fig. 1), each with predefined stations at 20–40, 75, 200, 350, 500, 750, and 1000 m depths, using a modified Atlantic Western IIA otter trawl (22.86 m head rope, 21.23 m footrope) with a 1.27-cm (0.5 in) mesh cod-end and intermediate liner. The trawl net was paired with Thyborøn Type II, 2.72-m (107 in) bottom-tending doors. Net deployments were generally for 20-min bottom-time and were standardized to a target speed-over-ground (SOG) of 1.49 m s−1 (2.9 kn; 2.7–3.1 kn acceptable range). Trawling was typically conducted along bathymetric contours unless local currents dictated the need for adjustment to ensure acceptable trawl performance. Scanmar CGM-05/TE40-2 net mensuration equipment was used in conjunction with door spread and trawleye sensors to monitor net performance and bottom-contact. A Furuno GP31 global positioning system (GPS) was used to monitor bottom-contact and lift-off positions, and SOG. Average door spread, speed, and the bottom-time of each net deployment was recorded and used to calculate area swept.
Fish catches were sorted to species onboard the vessel, measured for length, and each fish was given a unique identifier before freezing. Post-cruise, Arctic Cod were thawed, weighed, and gastrointestinal (GI) tracts were removed and fixed in 4 % formaldehyde solution. The selection of Arctic Cod for diet analysis was based on a published age–length relationship for the Canadian Beaufort Sea (Walkusz et al. 2013) to ensure representative coverage across available cohorts. Percent proportion of individual prey items was determined for N = 283 guts using standard laboratory methods and published keys (see Walkusz et al. 2013 for details). Gut contents were identified to the lowest taxonomic level possible. The weight of each taxon was determined based on published resources (see Walkusz et al. 2013 for details) in order to determine relative biomasses of different prey items.
Analysis of the distribution of Arctic Cod biomass across depth habitats
Biomass catch-per-unit-effort (CPUE) was calculated for each station as grams of Arctic Cod per square meter (g m−2) of sea floor swept. A Shapiro–Wilk procedure was applied to test the normality of CPUE data (α = 0.05). To assess whether Arctic Cod biomass differed significantly across station depths, biomass was grouped for depth stations across transects and a Kruskal–Wallis test was applied to mean values (α = 0.05). A Conover–Inman pairwise comparison was applied using the same depth groupings to determine whether depth stations grouped according to Arctic Cod biomass (α = 0.05). A Holm correction was applied to the p values of the Conover–Inman test to control familywise error rate.
Full oceanographic profiles of the water column were collected at each fishing station using a Seabird Model 25 conductivity, temperature, and depth (CTD) probe. Measurements were taken at 1m intervals throughout the water column. Fishing depth stations were overlaid onto integrated vertical water mass profiles of temperature and salinity (Ocean Data View v4) of the transects (Fig. 2) to assess spatial overlap of Arctic Cod biomass with differential water mass habitats. For comparison of temperature and salinity values relative to demersal fish catches, minimum, maximum, and mean values were calculated using the bottom-most three intervals from each cast.
Determining key prey items
Key prey items were identified based on cumulative percent contribution to total biomass of prey in guts. Starting from the highest contributor, and incrementing the total in descending order, species were added until a total equal to, or greater than, 85 % of the total gut biomass was achieved. A new Bray–Curtis similarity matrix was then constructed for these key prey items and used for follow-on similarity-based tests. The 85 % cutoff was selected to eliminate prey items that did not contribute substantially to total biomass, as well as larger prey items that occurred very infrequently in the diet (e.g., a few small Arctic Cod consumed as prey, see Results).
Detailed diet analyses
Hierarchical cluster analysis (HCA), using group-averaged linking, was performed on the Bray–Curtis similarity matrix to examine potential diet groupings based on key prey. A permutation-based similarity profile test (SIMPROF, PRIMER-E v7) was simultaneously applied to test for statistically significant (α = 0.05) clusters (Clarke and Gorley 2006). Non-metric multidimensional scaling (nMDS) was also performed on gut contents data, and SIMPROF groups from the HCA were labeled by color on the ordination (Fig. 5) to provide a concise visual representation of diet groupings. A similarity percentage (SIMPER, PRIMER-E v7) test was applied across SIMPROF groups to determine which key prey items contributed most to the average within-group similarities, thereby identifying the prey items that typified each group.
A permutation dispersion (PERMDISP, PRIMER-E v7) test was applied to gut content data across transects to determine whether dispersion differed significantly (α = 0.05). An analysis of similarity (ANOSIM, PRIMER-E v7) test was applied to determine whether gut contents differed significantly (α = 0.05) across transects overall.
A Shapiro–Wilk procedure was applied to test the normality of the standard length distribution of Arctic Cod (α = 0.05), followed by a Kruskal–Wallis test (α = 0.05) for difference in mean standard length among SIMPROF groups. A Conover–Inman pairwise comparison identified which groups were significantly different (α = 0.05), and a Holm correction was applied to control familywise error rate.
Results
The distribution of Arctic Cod biomass across depth habitats
At each transect, Arctic Cod biomass CPUE was highest at 350 and 500 m depth stations (Fig. 3). Mean standard length generally increased with station depth along each transect with a major size increase occurring at 200 m depth (Fig. 3). The standard length and biomass CPUE data failed the Shapiro–Wilk tests for normality (p > 0.05), dictating a nonparametric approach for further statistical testing. Mean biomass of Arctic Cod differed significantly (p < 0.01) among depth stations that were grouped across transects (i.e., 20- to 45-, 75-, 200-, 350-, 500-, 750-, 1000-m groups). Arctic Cod occurred in all habitats, though pairwise comparisons of grouped biomass across all station depths indicated that Arctic Cod biomass was significantly higher at the 350- and 500-m stations (Fig. 4).

a Biomass catch-per-unit-effort and b mean standard length of Arctic Cod (Boreogadus saida) across depth stations within transects

Mean Arctic Cod (Boreogadus saida) biomass CPUE and standard deviation pooled for station depths. The embedded table contains p values for pairwise tests of the mean biomass CPUE values for different depth stations (header box, i vs. j). Pairwise comparisons with significant p values (α = 0.05) are listed in the top box and indicated with letter symbols above error bars
The 20–45 m depth stations coincided with the Polar Mixed Layer and the 75 m depth stations coincided with the colder, more saline, Pacific water mass (Fig. 2). The 200 and 350 m stations sat at the upper and lower extremities of the Pacific–Atlantic thermohalocline, as indicated by the pronounced shift in temperature and salinity between these depths (Table 1). The 500 and 750 m depth stations coincided with the relatively warm and saline Atlantic water mass, and the 1000 m stations coincided with the colder Arctic water mass (Fig. 2). Peak Arctic Cod biomass coincided with the lower-thermohalocline (i.e., 350 m) and upper Atlantic water mass (i.e., 500 m) where the warmest mean water temperatures (0.44 and 0.41 °C, respectively) occurred, and salinities ranged between 34.67 and 34.86 PSU (Table 1). The mean temperatures, salinities, and associated standard deviations for grouped depth stations are presented in Table 1, along with minimum and maximum values for each parameter.
Determining key prey items
Four prey items contributed to a cumulative percent total of 86.10 % of total prey biomass in Arctic Cod guts: Calanus hyperboreus (37.98 %), Calanus glacialis (14.45 %), Themisto libellula (29.16 %), and Themisto abyssorum (4.51 %) (Table 2). Herein, these zooplankters are considered as “key prey items.”
Detailed diet analyses
There were five diet groups of Arctic Cod indicated by HCA with SIMPROF (p < 0.05) (Online Resource 1). The diet groups were generally supported by nMDS, as indicated by SIMPROF group labels on the associated two-dimensional ordination (stress = 0.11; Fig. 5). C. glacialis accounted for 100 % of the average similarity within group 1 indicating that, of the key prey items, this group fed exclusively on the smaller Calanus species (Table 3). Group 2 was typified by the larger copepod, C. hyperboreus, which accounted for 75 % of the average similarity within the group. The larger Themisto species, T. libellula, accounted for 93 % of average similarity within group 3, strongly typifying the diet of that group. C. hyperboreus (40 %) and T. libellula (41 %) contributed equally to the average similarity within group 4, with the smaller prey items C. glacialis and T. abyssorum each contributing approximately 10 %. Group 5 was typified by T. abyssorum, contributing 99 % of the average similarity, indicating that Arctic Cod within that group fed almost exclusively on the smaller Themisto species relative to other key prey (Table 3).

Non-metric multidimensional scaling (nMDS) ordination of Arctic Cod (Boreogadus saida) gut contents from all stations based on Bray–Curtis similarity. SIMPROF diet groups are indicated by symbol and color
Dispersion did not differ significantly (p > 0.05) within gut contents data across the four transects. The gut contents of Arctic Cod did not differ significantly across transects overall (p > 0.05), suggesting that the structure in the gut contents data, as reported in the SIMPROF groupings, was not strongly driven by differences in diet among transects (i.e., east to west).
Group 1 contained Arctic Cod guts from a relatively equal mix of shelf-based stations (i.e., 20–45 and 75 m; n = 26), which coincided with the Polar Mixed Layer and Pacific water mass, and slope-based stations (i.e., ≥200 m; n = 23) which coincided with the Pacific–Atlantic thermohalocline, upper Atlantic and Arctic water masses (Table 4). Group 2 coincided primarily with cod from 20 to 75 m depth stations (63 of 91), while groups 3–5 all coincided generally with cod from stations equal to, or deeper, than 200 m (34 of 35, 30 of 40, and 15 of 16 stations, respectively). Fish standard length differed generally among SIMPROF groups (p < 0.01; Fig. 6), with fish from groups 3 to 5 being significantly larger (p < 0.05) than those in group 1, and fish in groups 3 and 4 being significantly larger (p < 0.05) than fish in group 2.

Mean Arctic Cod (Boreogadus saida) standard length and standard deviation pooled for SIMPROF groups. The embedded table contains p values for pairwise tests of the mean standard length values for the different SIMPROF groups (header box, i vs j). Pairwise comparisons with significant p values (α = 0.05) are listed in the top box and indicated with letter symbols above error bars
In general, these results indicate that larger Arctic Cod occupied deeper slope habitats and fed predominantly upon larger Themisto prey. In particular, larger Arctic Cod associated with upper-slope habitats within the relatively warm lower Pacific–Atlantic thermohalocline and the upper Atlantic water mass. Smaller Arctic Cod primarily occupied shallower shelf-based habitats, within the colder Pacific water mass and overlying Polar mixed layer, and fed predominantly upon smaller Calanus prey. Fish size generally coincided with the size of prey consumed. Fish standard length, primary habitat occupancy, and characteristic prey items are summarized by diet group (i.e., SIMPROF group) in Table 4.
Discussion
This study documented the summer distribution of Arctic Cod in bottom habitats across the continental shelf and slope of the southern Canadian Beaufort Sea. The results indicate that peak Arctic Cod biomass occurs at 350 and 500 m depths in thermohalocline waters of the Pacific and Atlantic water masses and the upper Atlantic water mass. The size of Arctic Cod generally increased with depth, with a pronounced increase in standard length at 200 m which coincides with the outer continental shelf and the upper-extent of the Pacific–Atlantic thermohalocline. Using net-validated hydroacoustics to assess biomass of Arctic Cod in pelagic habitats of the Canadian Beaufort Sea, Geoffroy et al. (in press) observed that the average length and weight of Arctic Cod progressively increased with depth and that adults (age 1+ and older) predominantly associated with mesopelagic depths of 200–400 m throughout the year. Benoit et al. (2010) studied the winter/spring vertical distribution of Arctic Cod, copepods, and ringed seals in Franklin Bay and proposed that large Arctic Cod maintained depths in excess of 180 m in order to minimize predation by deep-diving mature seals. Smaller Arctic Cod also occupied depths below 180 m, but were observed to perform diel vertical migrations synchronized with the light/dark cycle, presumably to avoid foraging interference by larger Arctic Cod and to access food at shallower depths. The authors of these studies suggest that adult Arctic Cod occupy the mesopelagic layer in order to avoid predation by diving seabirds and marine mammals. We speculate that the risk associated with cannibalism in the near-bottom could be a reason why Arctic Cod do not descend to the bottom in early life-stages, but rather grow to a threshold size in the pelagial prior to the descent.
Our study supports previous work in the Canadian Beaufort Sea that indicates an association of Arctic Cod with the relatively warm waters of the upper Atlantic water mass, but also highlights differences in habitat associations of this species compared to adjacent areas. The relatively high biomass of Arctic Cod along the upper continental slope in this study coincided with the highest mean temperatures recorded across depth stations at 0.44 and 0.41 °C. Salinities at these depths were intermediate relative to the overlying Pacific water mass and the more saline underlying lower Atlantic and Arctic water masses. This biomass distribution matches observations by Crawford et al. (2012) who documented high densities of Arctic Cod in Atlantic waters along the continental slope at Barrow and Mackenzie Canyons at depths of 250–350 m and temperatures above 0 °C. The current study extends this distributional observation to the entire upper continental slope of the Canadian Beaufort Sea. The relatively low Arctic Cod biomass in the colder Pacific waters and those of the lower Atlantic and Arctic water masses may reflect a preference of Arctic Cod for warmer water temperatures. This result does not match findings by Logerwell et al. (2011) who found a negative correlation between Arctic Cod biomass and temperature in the western Alaskan Beaufort Sea, where the highest relative biomass was associated with cold (−1.5 °C or colder), highly saline (>33 PSU) waters from the Chukchi Sea, relative to warmer shelf waters. Results from Logerwell et al. (2011) were consistent with earlier research in the Alaskan Beaufort by Craig (1984) who documented an apparent association of high Arctic Cod catches with cold relative temperatures and high relative salinities in Simpson Lagoon. Logerwell et al. (2011) suggested that Arctic Cod may associate with the colder, carbon rich, offshore Chukchi waters due to higher relative secondary productivity compared to waters inshore, and thus, there being potentially more available prey. Primary production in our study area, or specifically the Beaufort shelf/slope, was highest in the upper water column (i.e., <100 m depth) where maximum chlorophyll concentrations and nitrate depleted waters were measured (Eert et al. 2015). The warm Atlantic waters coinciding with the high Arctic Cod biomass were well below the euphotic zone where primary production would be most directly linked with enhanced secondary production. Therefore, the significant cod layers observed in this study were not associated with waters rich in particulate organic carbon. The prey supporting the high Arctic Cod biomass at depth may rely on variable organic carbon fluxes (Forest et al. 2006), potentially receiving only a small portion of the organic matter produced within colder surface water layers (Forest et al. 2013). We suspect that zooplankton may concentrate at the Pacific–Atlantic water mass boundary either due to density differences between the water masses, thereby creating a barrier for planktonic organisms, or due to physiological response to the transition between negative and positive water temperatures. Further studies are required to validate this hypothesis. The documented differences in the habitat associations of Arctic Cod across different regions of the Beaufort Sea indicate complex habitat interactions that likely reflect a combination of factors including predator avoidance, prey availability, and optimal environmental conditions (e.g., thermal) to support physiological functions (e.g., growth).
Gut content analyses revealed that Arctic Cod in the Canadian Beaufort Sea fed on a diverse variety of prey but were highly dependent on Calanus copepods and Themisto amphipods. At least 40 prey taxa were documented in the diet of Arctic Cod during this study. Of these, 38 were zooplankton and two were fish including Arctic Cod and family Cottidae (Table 2). Despite the diversity of the cod diet, which suggests opportunistic feeding habits, C. hyperboreus, C. glacialis, T. libellula, and T. abyssorum accounted for the vast majority of the biomass in guts. These zooplankters are widely found throughout the Arctic and offer significant energy supply due to their high lipids content (Scott et al. 2000; Auel et al. 2002). Because these species contributed the majority of the combined food biomass in our study, and have high energy content, they can be regarded as key species supporting Arctic Cod populations in the Canadian Beaufort Sea. In a 2008 survey in the western US Beaufort Sea, Rand et al. (2013) observed that demersal Arctic Cod fed primarily on fish (>75 % by wet weight), though calanoid copepods were also prominent from a percent count perspective. Fish in pelagic catches had fed primarily on copepods (frequency of occurrence). Fish comprised 5.3 % of total prey biomass in this study, with Arctic Cod accounting for 4.3 %; however, only six Arctic Cod were found within four guts (1.4 % of all guts) reflecting their relatively large size and suggesting that cannibalism was limited during the study. T. abyssorum accounted for a similar proportion of prey biomass (4.5 %) in this study, yet their biomass was comprised of 1200 individual specimens and the species occurred in 25.1 % of guts. Walkusz et al. (2013) examined the diet of bottom-dwelling Arctic Cod on the Canadian Beaufort Shelf in 2006–2009 and found that copepods (Pseudocalanus spp., C. glacialis, C. hyperboreus, Limnocalanus macrurus, and Jaschnovia tolli), amphipods (Apherusa glacialis and T. libellula), and mysids (Mysis oculata) were important diet items, with C. glacialis and C. hyperboreus being prevalent across all age-classes examined. Three of the four key prey items in our study coincide with those identified by Walkusz et al. (2013). T. abyssorum was not considered an important prey species in the Walkusz et al. (2013) study, likely due to its occurrence predominantly in Atlantic-derived waters (Koszteyn et al. 1995), which coincide with deep slope habitats in the Canadian Beaufort Sea and outside of the sampling range of Walkusz et al. (2013). Nahrgang et al. (2014) reported that bottom-caught Arctic Cod associated with the Arctic water mass at Rijpfjorden, Svalbard, fed primarily on T. libellula which occurred in 100 % of guts sampled. The results of the Rand et al. (2013), Walkusz et al. (2013) and Nahrgang et al. (2014) studies suggest both local and regional differences in the feeding habits of Arctic Cod and likely reflect differences in the types and quality of the prey that are available to them in a given area and time (season).
Differences in the prey composition among the SIMPROF diet groups coincided generally with the known habitat associations of the four key prey species, and the size of the prey items that typified the groups correlated positively with the mean standard length of Arctic Cod. The four key prey taxa identified in this study are typical marine zooplankton and the lower salinity and higher temperature/turbidity found on the Canadian Beaufort Shelf (due to the Mackenzie River plume) are, to some extent, sub-optimal conditions for them. C. hyperboreus is considered a true Arctic species that is abundant in the Arctic Ocean (Hirche and Mumm 1992), and thus, its greater prevalence in guts from fish captured at offshore upper-slope stations is not surprising. Both Themisto species are found in Arctic Seas, but only T. libellula is a genuine Arctic species, while T. abyssorum is considered subarctic-boreal (Koszteyn et al. 1995) and thus seems to be better adapted to the warmer water found deeper on the slope. C. glacialis is considered a “shelf” species that spawns in Arctic marginal seas (Falk-Petersen et al. 2009) and prefers shallower locations, although it occurs widely. The five diet-based SIMPROF groups showed a transition from C. glacialis in shelf habitats to the larger C. hyperboreus on the upper slope, then to the Themisto species in offshore slope habitats. This shift in diet with depth was previously observed on the Canadian Beaufort Shelf (Walkusz et al. 2013) and can likely be attributed both to differences in the general distributions of the prey items as described above (i.e., prey availability) and to a positive relationship between predator and prey size (i.e., prey selection; Scharf et al. 2000). In general, the smallest Arctic Cod within either shelf or slope habitats fed primarily on the smallest key prey items likely available. The relative size of the primary diet item(s) that typified the SIMPROF diet groups generally increased with mean Arctic Cod standard length across the groups.
Arctic Cod is a keystone species in Arctic marine ecosystems, understood to be a significant consumer of zooplankton and is a key prey species for marine mammals and birds. While it occurs throughout the circumpolar north, distributions at more localized scales are less understood, including seasonal habitat and diet associations in deep (>200 m) offshore habitats. Knowledge of these factors is a key building block to understanding the role of Arctic Cod in the structure and function of the Beaufort Sea ecosystem. This study attempted to address these issues for demersal Arctic Cod in the southern Canadian Beaufort Sea. We documented the distribution of demersal Arctic Cod biomass across shelf and deep-water slope habitats, demonstrating that Arctic Cod occupies virtually all marine benthic habitats in the region. However, the diet of Arctic Cod differed with size and within shelf and slope habitats. Because of its central role in Arctic marine food webs, Arctic Cod and its habitats and prey need to be considered in mitigation and conservation efforts related to Oceans Management and offshore resource development. Impacts to Arctic Cod, at both regional (i.e., population-level) and local (e.g., within critical habitats that support life-history stages) scales could have wide-ranging and significant impacts on the ecosystem.
References
Andriyashev AP, Mukhomediayaroz BF, Pavshtiks EA (1980) On mass congregations of the cryopelagic cod fishes (Boreogadus saida and Arctogadus glacialis) in circumpolar arctic basins. In: Vinogradov ME, Melnikav LA (eds) Biology of the central Arctic Basin. Shirov Institute of Oceanology, Academy of Sciences, pp 196–211. (National Museums of Canada, translation, 1981; 23 p)
Auel H, Harjes M, Da Rocha R, Stübing D, Hagen W (2002) Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipods Themisto abyssorum and T. libellula. Polar Biol 25:374–383
Benoit D, Simard Y, Fortier L (2008) Hydroacoustic detection of large winter aggregations of Arctic cod (Boreogadus saida) at depth in ice-covered Franklin Bay (Beaufort Sea). J Geophys Res Ocean 113:C06S90
Benoit D, Simard Y, Gagné J, Geoffroy M, Fortier L (2010) From polar night to midnight sun: photoperiod, seal predation, and the diel vertical migrations of polar cod (Boreogadus saida) under landfast ice in the Arctic Ocean. Polar Biol 33:1505–1520
Benoit D, Simard Y, Fortier L (2014) Pre-winter distribution and habitat characteristics of polar cod (Boreogadus saida) in Southeastern Beaufort Sea. Polar Biol 37:149–163
Bouchard C, Fortier L (2008) Effects of polynyas on the hatching season, early growth and survival of polar cod Boreogadus saida in the Laptev Sea. Mar Ecol Prog Ser 355:247–256
Bradstreet MS, Finley KJ, Sekerak AD, Griffiths WB, Evans CR, Fabijan MF, Stallard HE (1986) Aspects of the biology of Arctic cod (Boreogadus saida) and its importance in arctic marine food chains. Can Tech Rep Fish Aquat Sci 1491:viii + 193 p
Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial. PRIMER-E, Plymouth
Craig PC (1984) Fish use of coastal waters of the Alaskan Beaufort Sea: a review. Trans Am Fish Soc 113:265–282
Craig PC, Griffiths WB, Haldorson L, McElderry H (1982) Ecological studies of Arctic cod (Boreogadus saida) in Beaufort Sea coastal waters, Alaska. Can J Fish Aquat Sci 39:395–406
Crawford RE, Vagle S, Carmack EC (2012) Water mass and bathymetric characteristics of polar cod habitat along continental shelf and slope of the Beaufort and Chukchi seas. Polar Biol 35:179–190
Drost HE, Carmack EC, Farrell AP (2014) Upper thermal limits of cardiac function for Arctic cod, Boreogadus saida, a key food web fish in the Arctic Ocean. J Fish Biol 84:1781–1792
Eert J, Meisterhans G, Michel C, Niemi A, Reist J, Williams WJ (2015) Physical, chemical and biological oceanographic data from the Beaufort Regional Environmental Assessment: Marine Fishes Project, August–September 2012. Can Data Rep Hydrogr Ocean Sci 197:vi + 83 p
Falk-Petersen S, Mayzaud P, Kattner G, Sargent JR (2009) Lipids and life strategies of Arctic Calanus. Mar Biol Res 5:18–39
Forest A, Sampei M, Hattori H, Makabe R, Sasaki H, Fukuchi M, Wassmann P, Fortier L (2006) Particulate organic carbon fluxes on the slope of the Mackenzie Shelf (Beaufort Sea): physical and biological forcing of shelf-basin exchanges. J Mar Syst. doi:10.1016/j.jmarsys.2006.10.008
Forest A, Babin M, Stemmann L, Picheral M, Sampei M, Fortier L, Gratton Y, Bélanger S, Devred E, Sahlin J, Doxaran D, Joux F, Ortega-Retuerta E, Martín J, Jeffrey WH, Gasser B, Carlos Miquel J (2013) Ecosystem function and particle flux dynamics across the Mackenzie Shelf (Beaufort Sea, Arctic Ocean): an integrative analysis of spatial variability and biophysical forcings. Biogeosciences 10:2833–2866. doi:10.5194/bg-10-2833-2013
Fortier L, Sirois P, Michaud J, Barber D (2006) Survival of Arctic cod larvae (Boreogadus saida) in relation to sea ice and temperature in the Northeast Water Polynya (Greenland Sea). Can J Fish Aquat Sci 63:1608–1616
Geoffroy M, Dominique R, Gérald D, Fortier L (2011) The aggregation of polar cod (Boreogadus saida) in the deep Atlantic layer of ice-covered Amundsen Gulf (Beaufort Sea) in winter. Polar Biol 34:1959–1971
Geoffroy M, Majewski A, LeBlanc M, Gauthier S, Walkusz W, Reist JD, Fortier L (2015) Vertical segregation of age-0 and age-1 + polar cod (Boreogadus saida) over the annual cycle in the Canadian Beaufort Sea. Polar Biol (in press)
Gradinger RR, Bluhm BA (2004) In-situ observations on the distribution and behavior of amphipods and Arctic cod (Boreogadus saida) under the sea ice of the High Arctic Canada Basin. Polar Biol 27:595–603
Hirche HJ, Mumm N (1992) Distribution of dominant copepods in the Nansen Basin, Arctic Ocean in summer. Deep Sea Res PtII 39:S485–S505
Hodgson R, Martin K, Melling H (2015) Marine protected area network planning in the Western Arctic Bioregion: development and use of a classification system to identify ecological units as required planning components. DFO Can Sci Advis Sec Res Doc 2015/020. v+ 41 p
Koszteyn J, Timofeev S, Weslawski JM, Malinga B (1995) Size structure of Themisto abyssorum (Boeck) and Themisto libellula (Mandt) populations in European Arctic Seas. Polar Biol 15:85–92
Logerwell E, Rand K, Weingartner TJ (2011) Oceanographic characteristics of the habitat of benthic fish and invertebrates in the Beaufort Sea. Polar Biol 34:1783–1796
Lønne OJ, Gulliksen B (1989) Size, age and diet of Polar cod, Boreogadus saida (Lepechin 1773), in ice covered waters. Polar Biol 9:187–191
Lowdon MK, Majewski AR, Reist JD (2011) Fish catch data from Herschel Island, Yukon Territory, and other offshore sites in the Canadian Beaufort Sea, July and August 2008, aboard the CCGS Nahidik. Can Data Rep Fish Aquat Sci 1237:vi + 99 p
Majewski AR, Reist JD, Park BJ, Sareault JE, Lowdon MK (2009a) Fish catch data from offshore sites in the Mackenzie River estuary and Beaufort Sea during the open water season, July and August, 2005, aboard the CCGS Nahidik. Can Data Rep Fish Aquat Sci 1204:vii + 53 p
Majewski AR, Reist JD, Park BJ, Lowdon MK (2009b) Fish catch data from offshore sites in the Mackenzie River estuary and Beaufort Sea during the open water season, July and August, 2006, aboard the CCGS Nahidik. Can Data Rep Fish Aquat Sci 1218:vii + 37 p
Majewski AR, Lowdon MK, Reist JD, Park BJ (2011) Fish catch data from Herschel Island, Yukon Territory, and other offshore sites in the Canadian Beaufort Sea, July and August 2007, aboard the CCGS Nahidik. Can Data Rep Fish Aquat Sci 1231:vi + 50 p
Majewski AR, Lynn BR, Lowdon MK, Williams WJ, Reist JD (2013) Community composition of demersal marine fishes on the Canadian Beaufort Shelf and at Herschel Island, Yukon Territory. J Mar Sys 127:55–64
McLaughlin FA, Carmack EC, Macdonald RW, Bishop JKB (1996) Physical and geochemical properties across the Atlantic/Pacific water mass front in the southern Canadian Basin. J Geophys Res 101:1183–1197
McLaughlin F, Shimada K, Carmack E, Itoh M, Nishino S (2005) The hydrography of the southern Canada Basin, 2002. Polar Biol 28:182–189. doi:10.1007/s00300-004-0701-6
Moulton LL, Tarbox KE (1987) Analysis of Arctic Cod movements in the Beaufort Sea nearshore region, 1978–79. Arctic 40:43–49
Nahrgang J, Varpe Ø, Korshunova E, Murzina S, Hallanger IG, Vieweg I, Berge J (2014) Gender specific reproductive strategies of an Arctic key species (Boreogadus saida) and implications of climate change. PLoS One 9(5):e98452. doi:10.1371/journal.pone.0098452
Rand KM, Logerwell EA (2011) The first demersal trawl survey of benthic fish and invertebrates in the Beaufort Sea since the late 1970s. Polar Biol 34:475–488
Rand KM, Whitehouse A, Logerwell EA, Ahgeak E, Hibpshman R, Parker-Stetter S (2013) The diets of polar cod (Boreogadus saida) from August 2008 in the US Beaufort Sea. Polar Biol 36:907–912
Scharf FS, Juanes F, Rountree RA (2000) Predator size-prey size relationships of marine fish predators: interspecific variation and effects of ontogeny and body size on trophic-niche breadth. Mar Ecol Prog Ser 208:229–248
Scott CL, Kwasniewski S, Falk-Petersen S, Sargent JR (2000) Lipids and life strategies of Calanus finmarchicus, Calanus glacialis and Calanus hyperboreus in late autumn, Kongsfjorden, Svalbard. Polar Biol 23:510–516
Walkusz W, Majewski A, Reist JD (2013) Distribution and diet of bottom dwelling Arctic Cod in the Canadian Beaufort Sea. J Mar Syst 127:65–75
Welch HE, Crawford RE, Hop H (1993) Occurrence of Arctic Cod (Boreogadus saida) schools and their vulnerability to predation in the Canadian High Arctic. Arctic 46:331–339
Acknowledgments
We thank the crew of the F/V Frosti for their dedication and professionalism at sea and the staff at Frosti Fishing Ltd for technical and logistical support throughout the program. We are also thankful to Lorena Edenfield (University of Alaska Fairbanks) and the late Charlie Ruben (Paulatuk, NT) for help with onboard sampling. The BREA (Aboriginal Affairs and Northern Development Canada), Environmental Studies Research Fund (Natural Resources Canada), International Governance Strategy (Fisheries and Oceans Canada), Program of Energy Research and Development (Natural Resources Canada), internal Fisheries and Oceans Canada sources, and ArcticNet provided funding for this research. We are grateful to the Inuvialuit Game Council for supporting this project and for providing valuable input to the study design and logistics.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article belongs to the special issue on the “Ecology of Arctic Gadids”, coordinated by Franz Mueter, Jasmine Nahrgang, John Nelson, and Jørgen Berge.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
Hierarchical cluster analysis of Arctic Cod (Boreogadus saida) gut content samples. Solid lines indicate groupings that are statistically different at α = 0.05 (TIFF 145 kb)
Rights and permissions
About this article

Cite this article
Majewski, A.R., Walkusz, W., Lynn, B.R. et al. Distribution and diet of demersal Arctic Cod, Boreogadus saida, in relation to habitat characteristics in the Canadian Beaufort Sea. Polar Biol 39, 1087–1098 (2016). https://doi.org/10.1007/s00300-015-1857-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-015-1857-y