
Abstract
Monitoring the effectiveness of management activities within reserves is always a complicated task. When the focus of management activities is mammals, it is difficult to monitor their populations in a way that is rapid, effective, and inexpensive. We report on a mammal survey of a reserve in southwest China using remote-trip cameras. We surveyed 329 locations over 2 field seasons in 2002 and 2003. Sixteen species of mammals were detected with these cameras, with four species documented for the first time. After accounting for variation due to slope, aspect, elevation, and habitat type, the distribution of six species was positively associated with the location of conservation stations and/or patrolling routes. Species of medium-sized mammals are excellent candidates for monitoring programs based on these cameras, due to their relative abundance, sufficient size to be detected by the camera units, and sensitivity to human activity. The distribution of mammals relative to management efforts is a relatively rapid means to assess reserve effectiveness. The repeat use of the cameras as part of a monitoring plan should provide a quantifiable measure of reserve effectiveness.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wildlife reserves are under pressure to demonstrate effectiveness as conservation tools (McNeely 1989; Parrish and others 2003; Schwartman and others 2000). International organizations encourage reserves to engage in conservation planning that includes quantifiable goals and measures of success (Hockings 2003; Margoluis and Salafsky 1998; Parrish and others 2003). The push for quantitative measures of success coincides with an increased emphasis on global monitoring of biodiversity by standardized protocols (Green and others 2005). Within a reserve’s budget, however, monitoring must compete for limited conservation funds with other worthwhile programs, such as incentive programs in local communities (Fu and others 2004; Lindsey and others 2005), remote sensing technology (Nagendra and others 2004), and reserve staff infrastructure and training (Rao and others 2002). In a resource-limited environment, reserve management must demonstrate the effectiveness of their staff in conserving wildlife within the reserve (Atauri and others 2005; Parrish and others 2003).
One direct measure of reserve success is the abundance of critical species. Assessing the status of wildlife populations is a necessary component for indexes of sustainability for hunted species (Carrillo and others 2000; Milner-Gulland and Akcakaya 2001), biodiversity measures (Caro 1999), and the effectiveness of conservation plans (Atauri 2005; Hockings 2003; Mace and others 2001; Possingham and others 2001). The utility of these measures is contrasted by the difficulty in assessing some groups of species, such as large and medium-sized mammals in forested habitat (Caro 1999). The cryptic and nocturnal habits of most mammals often result in population estimates based on indirect measures, such as tracks and fecal dropping (Carrillo and others 2000; Gaines 2001; Grigione and others 1999; Schauster and others 2002; Weckerly and Ricca 2000; White 1994). The disadvantage of sign surveys is the inability to discriminate between species that overlap in body size and food selection (Wemmer and others 1996). Direct measures of populations through either sightings or captures can prove to be expensive and time-consuming (Caro 1999; Wemmer and others 1996).
Population estimates obtained over a number of years form the basis of most monitoring programs for mammals (Carrillo and others 2000; Gibbs 2000). Hockings (2003) found that monitoring programs are a major component of over half of the conservation plans associated with reserve management. Monitoring activities, however, often have shortfalls of unclear objectives, inadequate sampling, inappropriate timescales, and primitive analysis (Possingham and others 2001). With mammals, one major issue is whether to base a monitoring program on direct or indirect population measures. Errors in sign detection and the inherent variability of wild populations makes monitoring animal abundance via indirect measures advisable only over long time periods (Gibbs 2000; Wemmer and others 1996). This is especially true for medium-sized mammals whose population variability is four times that of large mammals (Gibbs 2000). This limitation precludes any use of mammals as indicator species for a short-term assessment of reserve effectiveness (Ervin 2003). This is unfortunate because large and medium-sized mammals are often the focus of poaching activities within reserves (Carrillo and others 2000; Robinson and Bodmer 1999). Although quantitative measures should not be the sole focus of monitoring programs (Hockings 2003; Stem and others 2005), improvements in mammal surveys that allow more rapid and quantifiable results would make adaptive management practices more feasible.
Surveys of mammals in tropical and temperate habitats increasingly rely on remote-trip cameras to detect cryptic mammals (Karanth and Nichols 1998; Moruzzi and others 2002; Numata and others 2005; Sanderson and Trolle 2005). These surveys can be specific for a single species or a measure of species richness. The cameras work best when individual animals can be recognized by natural or man-made markings (Karanth and Nichols 1998, Carbone and others 2001, Trolle and Kery 2003). When individuals cannot be recognized, cameras still perform better than indirect measures for detecting species of small carnivores (Foresman and Pearson 1998). The photographs have been used to create an abundance index using either photographs/camera night or number of days until a species is first detected (Carbone and others 2001; Moruzzi and other 2002). The recent development of detection probability theory (MacKensie 2005; Royle and Nichols 2003) would strengthen the index by generating a “capture history” of a species when there are repeat surveys of a site. We believe that there is a valuable role for remote-trip cameras as a monitoring tool for reserve management.
We report on a general survey of mammals in a reserve in northeastern Sichuan Province in the People’s Republic of China. These survey data were examined with respect to conservation activities using logistic regression. We isolated the impact of human activity (i.e., conservation stations, patrolling routes, roads, and local villages) from landscape variables such as slope, aspect, elevation, and habitat type. Our hypothesis was that there was a discernable benefit to wildlife from patrolling activities and stationing of reserve staff within the reserve. This benefit can be assessed relatively rapidly due to the remote-trip camera and using the distribution of mammals as a metric of a reserve’s success.
Study Area
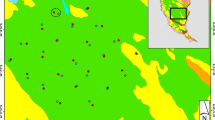
The Tangjiahe Nature Reserve is an approximately 300-km2 reserve in northern Sichuan Province (104°36′–104°53′ latitude and 32°32′–32°41′ longitude) that was set aside for giant panda (Ailuropoda melanoleuca) and takin (Budorcas taxicolor) conservation in 1978 (Tangjiahe NRAD 2000). The terrain comprises steep river valleys with an elevation range from 1150 to 3800 m. The reserve is predominately (> 80%) forested with subalpine meadows at higher elevations and early succession patches along river valleys. All local people were relocated outside of the reserve at its inception, most to just outside the reserve entrance (Fig. 1). A conservation station was situated near the entrance of the reserve and a second station was located in the center of the reserve. We used two surrogate measures of conservation activity. First, there were four patrolling teams that used set routes to patrol the reserve every 2 weeks; our first conservation measure was proximity to these patrolling routes. Second, the patrolling staff were housed at two conservation stations and inclement weather could cause patrols to be abandoned or shortened, so that patrolling activity, and probably all staff activity, was more common nearer the conservation stations (Li Ming Fu, personal communication). There were two roads within the reserve that followed the main rivers; one provided transit to the central conservation station (which was also the reserve headquarters) and the other continued approximately halfway along the other river valley. Due to the steep terrain along the northern and western boundaries, the main entrance for poachers was probably along the Tangjiahe River.

Extent of the Tangjiahe Nature Reserve in Sichuan Province, China. Conservation stations, patrolling routes, and villages used in the analysis are indicated.
Methods
The general schematic of a survey camera is for an infrared sensor to be attached to a camera with built-in flash, auto-advance and auto-focus features (http://www.camtrakker.com; Swann and others 2004). The sensor detects the heat profile of a cone-shaped area in front of the unit and is triggered by abrupt changes in temperature across that cone. A limitation is the sensitivity of the sensor, which can vary depending on structural interference (i.e., vegetation), the temperature difference between the animal and the environment, battery strength, and manufacturer (Swann and others 2004). This difference in sensitivity results in longer detection ranges in open habitat and larger animals being detected at longer distances when the habitats are similar. We compensated for this by limiting the range of the sensor to within 3 m of the unit through careful orientation toward the landscape and vegetation features. The 3-m limit is within recommended guidelines [2–5 m by Swann and others (2004)]. The 3-m limitation is not to allow comparison between species, but to eliminate any bias due to differences in temperature, or vegetation density, between the sampling points.
The entire reserve was divided into 71 4-km2 blocks for sampling. We attempted to survey all blocks, but steep terrain resulted in only 56 blocks being surveyed. All data were collected from March to December 2002 and from March to October 2003, with no data collected during winter months. The elevation range for the sample locations was 1100–3200 m. Camtrakker® infrared triggered cameras (n = 5) were placed in each block for a 2-week period. Within the survey block, cameras were placed in likely animal-use areas, as determined by reserve staff, and placed a minimum of 150 m apart. We cannot assume independence between sampling points, especially for the largest mammals, whose potential home range could encompass the entire reserve. Using standard terminology (MacKensie 2005), we are assuming “occupancy” for the reserve if a species is detected across the entire survey, but we measure a species “use” at a specific location across a range of potential locations within the reserve. We selected the minimal distance of 150 m between sample points based on our knowledge of the movements of medium-sized mammals and the logistics needed to setup or retrieve multiple cameras on the same day. Cameras were placed and retrieved as part of patrolling duties of the reserve staff. Because cameras were placed by different patrolling teams over a 19-month period, subsequent examination of sample points found some points less than 150 m apart. For these nearby points, we included the data in the species list of occupancy for the reserve, but we eliminated some points prior to statistical analysis of use. For each set of nearby points, we retained the point with the most detections and in the rarer habitat type. To increase the probability of detecting carnivores with large home ranges, a dollop of carnivore scent lure, obtained from a commercial trapping supplier (Montgomery Fur Co., Ogden, Utah), was placed within the detection range of each camera (Moruzzi and others 2002). A preliminary survey of captive ungulates species did not show a significant response to lures placed within their enclosure (D. Powell, Wildlife Conservation Society, unpublished communication). Cameras were checked at the end of the 14-day period, and if the camera was not operating, the date of the last picture was considered the last date of sampling (average sampling period =13.7 days).
For all analyses, we used detection/nondetection at a location, rather than the number of pictures. The reason for this is that abundance has a strong seasonal component and it is logistically impossible to sample the entire reserve simultaneously; for example, our sampling covered from spring through autumn over 2 years. Ecoregional monitoring programs in North America are based on presence/absence samples (Manley and others 2005). For the smaller species ( < 300 g), a definitive species identification was not always possible without a specimen, but we assigned a putative species identification based on relative size, coloration, and known distribution of the species in the region (Ma and others 2001; Tangjiahe NRAD 2000). When species identification was not possible based on incomplete picture or poor focus, we discarded the detection.
Variables calculated for each camera location included slope, aspect, elevation, habitat type, distance to nearest conservation station, distance to nearest patrolling route, and distance to nearest village. Habitat type was determined from on-site examination of camera location, with 5 habitats types sampled (Mixed Broadleaf, n = 128; Conifer And Broadleaf, n = 88; Conifer, n = 11; Evergreen and Broadleaf, n = 79; and Alpine, n = 20). Using ARCView 3.3 (ESRI Inc., Redlands, CA), slope, aspect, and elevation were derived from the digital elevation model of the reserve (Fig. 2). Distance values were determined within ARCView using Euclidean distances between attributes.

The distribution of 329 camera locations within the Tangjiahe Nature Reserve, with respect to elevation. Camera locations with solid symbols were eliminated prior to analysis due to close proximity to other points, but were included in the list of species detected
Multiple logistic regressions was used for analysis of animal distributions relative to human activity. A stepwise backward progression algorithm within SYSTAT (Vers. 11) was used with P = 0.10 for variable removal or entry. Aspect and habitat were coded as binary variables. The aspect was divided between “Cold” (i.e., NE, N, NW, W) and “Warm” (i.e., SW, S, SE, E). Habitat associations were examined a priori for each species and the most obvious preferred habitat was compared to all other types in the logistic analysis. Prior to analysis, each continuous variable was examined for Normal distribution (no transformations were necessary). We examined all variables for significant collinearity and found distance to conservation station correlated with distance to village (r = 0.73); we included an interaction term for this set of variables in all analyses. Even though the conservation stations and villages were located along the main river drainage, the extreme terrain resulted in no significant correlation between elevation and either distance to conservation station (r = 0.08) or distance to people (r = 0.06).
Results
We surveyed 392 sample points within the reserve, for a total detection of 509 pictures of 19 species (16 mammal species; Table 1). Although significant sampling effort (4515 camera-days) was devoted to the survey, predator species were rarely detected within the reserve. A previous sign survey (National Giant Panda Survey, unpublished data) detected two large species (giant panda and Asian black bear Ursus thibetanus) that were not detected in the camera survey. The camera units provided the first physical evidence of several species within the reserve: leopard cat (Prionailurus bengalensis), goral (Naemorhedus goral), serow (Capricornis sumatraensis), and musk deer (Moschus moschiferus).
The average number of days before a camera unit records a species can be used to assess if camera monitoring will effectively monitor a specific species (Moruzzi and others 2002). Although cameras were equipped to record the time and date of each picture, this feature often failed on our units. There were, however, six species for which we were able to estimate the average number of days before the species was first detected. For most of these species, the average date of the first picture was approximately 1 week (Tibetan macaque, 6 days; takin and serow, 7 days; reeve’s muntjac, 8 days; and tufted deer, 9 days); for a single species, the average date of first detection was close to the end of the survey period (i.e., goral, 12 days).
For 13 mammal species, there were sufficient detections (more than three locations) to examine their distribution relative to human activity (villages, patrolling routes, and conservation stations). We failed to detect a distribution pattern for two species; one species (i.e., takin) was the most detected species (Table 1) and, essentially, was found throughout the reserve. The second species, rock squirrel (Sciurotamias davidianus), was one of the rarer species detected and the sample size could explain the lack of a discernable pattern. Our survey found only two species, porcupine and hog badger, confined to specific elevation bands (1400–2500 m and 1250–2500 m, respectively). For eight species, distance to conservation station, patrolling route, or village was a significant factor in their detection (Table 2). A graph of mean detection distance from conservation station for each species shows most species closer than the average survey point (Fig. 3). The golden monkey (Rhinopithecus roxellanae) was one of the species distributed negatively with respect to conservation activity—in this case, patrolling routes.

Mean distance to the nearest conservation station for all 271 survey points used in the analysis (crosshatched bar) and the mean distance for detections for each species that was shown to be significantly affected by that variable in a logistic regression (single hatch mark). The mean standard error is shown for each species bar.
For three species for which both conservation station and village distances were significant, there was a significant interaction between the two variables, indicating that animals do not respond similarly to the two correlated variables. For two of those species, white-bellied rat and Himalayan weasel, they were both more abundant closer to human habitation. For the third species, tufted deer (Elaphodus cephalophus), the data show few detections in the area between the villages and the conservation stations, but detections increased once a conservation station was between the village and the sample point. This species was also found closer to patrolling routes than expected by chance (Table 2).
Discussion
The cameras provide demonstrable proof that species exist in reserves. Scat and tracks had been attributed to these species by reserve staff, but due to the overlap in sign characteristics among several species, positive identification was not possible until this survey. Previous attempts to show the distribution of species relied on species’ geographic ranges and elevation restrictions to indicate probable occurrence (Tangjiahe NRAD 2000). We only found two species to respond significantly to the elevation range sampled: porcupine and hog badger. Although there are 85 potential mammal species in the reserve (Tangjiahe NRAD 2000), the majority has never been seen or captured (Li, personal communication). The camera records provide a suite of species that are abundant enough for monitoring purposes.
This reserve contained several hoofed mammals that overlap in body size: tufted deer, muntjac (Muntiacus reevesi), serow, goral, musk deer, and wild boar (Sus scufa). In most instances, examination of tracks would not enable the staff to differentiate among these species because the juveniles of the larger-bodied species produced tracks similar to adults of the smaller-bodied species (Ma and others 2001). Fecal droppings for several species also overlap in size and characteristics (Ma and others 2001). The remote-trip cameras allowed us to differentiate among these species (Table 1) and to determine the correlates for species-specific distributions within the reserve (Table 2).
Two species increased with proximity to village. The white-bellied rat is a granivore (Sheng and others 1999) and might be attracted to agricultural crops planted near villages. The Himalayan weasel, a small predator of rodents, was also found closer to villages. For the tufted deer, the relationship between distribution and management activity is complex. Detections were farther away from villages and conservation stations, but closer to patrolling routes. Little natural history information is available for this species (Wemmer 1998), but if villages are the primary source of poachers and poaching is heavy on this species, then our data indicate that the location of conservation stations near villages does not compensate for the negative impact of villages. Only conservation activity far from villages (i.e., patrolling routes) coincides with the increased presence of this species.
For golden monkeys, patrolling activity was negatively associated with detections. At Tangjiahe, this species is very sensitive to human disturbance, and upon sighting humans, individuals produce alarm calls that cause an entire troop to flee (Ge 2000). Being arboreal, golden monkeys are hunted with guns, not snares as with most other mammal species. Our data indicate that they do not remain in areas with human activity, regardless of its nature.
Overall detections of mammals within the reserve were low, with a marked absence of many predator species. This result would not be due to lack of available habitat, as this reserve is adjacent to two other reserves (Baishuijing and Dongyanggou nature reserves), and, together, these three reserves encompass a 3680-km2 forest block. We should consider whether limitations of the survey protocol or the cameras produced these low detection rates. Our schedule for camera relocation (i.e., 14 days) was set by the patrolling schedule of the reserve staff. It is possible that the relatively rapid shifting of cameras reduced our ability to evaluate any specific location for species with large home ranges. However, the measure of any indirect survey is “effort” (Gibbs 2000; Wemmer and others 1996); in our case, this was the number of days multiplied by the number of cameras active each day. Our effort (> 4500 camera days) was beyond 18 of 20 studies designed to detect tigers (Pan tigris), a usually scarce predator, in other Asian countries (Carbone and others 2001). There is also no indication of undersampling for the few species for which we able to calculate the mean number of days until first photograph. We know of two large mammal species that occur in the reserve and were not detected during the survey: giant panda and Asiatic black bear. We have detected these species using the cameras at other reserves (unpublished data) and the black bear was detected by a remote-trip camera at Tangjiahe shortly after this survey ended. Densities of these animals within the reserve must be below that for detection at our sampling effort. Schaller and others (2000) estimated both species at less than 20 individuals within the reserve in the 1980s. The low predator density could also be a legacy of the reserve’s recent past, having been only established from a logging concession in the late 1970s.
The reliance of the camera unit on detecting heat profiles of approaching animals compromises our ability to compare species that differ in body size. For example, the most abundant species photographed was also the largest mammal (takin) and the least photographed included two of the smallest mammals [white-bellied rat (Niviventer andersoni) and Himalayan weasel (Martes sibirica)]. Beause one of the primary functions of the reserve was to preserve the takin, the abundance of photographs at multiple locations is an indication that the takin population is abundant and widespread. However, we would not infer from these data that the takin population is more abundant than the population of white-bellied rats because of differences in detection distance (Swann and others 2004).
Although the utility of comparisons between species is limited, there is validity in comparing “capture” locations within a species. Mammal species are relatively mobile and the distribution of animals across a landscape or reserve should indicate the location of favorable and unfavorable locations for species (Garshelis 2000). Locations are favorable due to both environmental factors (i.e., temperature, moisture, and forest types) and management activities (i.e., lack of poaching activity or disturbance and habitat quality). Changes in effectiveness of conservation stations and patrolling routes should be reflected by changes in species distributions, taking into account differences in environmental factors. For 6 of 13 species tested, our surrogate measure of conservation activity (i.e., distance to a conservation station or patrolling route) was positively associated with the species detection, indicating that patrolling by the reserve staff does have a conservation value. For two species, animals were found farther from villages. Fu and others (2004) discussed the pressures on villagers in China to derive income from nearby nature reserves. We do not know the mechanism by which the conservation stations enhance mammal detections; either the patrolling activities itself or the presence of staff within specific valleys or drainages might deter poaching. For most species, the results indicate that conservation stations are of value, and, possibly, additional stations should be established at higher elevations to enhance conservation within those habitats. The addition of conservation stations or patrolling routes should expand the range of several species, and this change would be rapidly detected with the remote-trip cameras. For the golden monkey, for which the response to human activity was negative, it might need focused management attention that cannot be provided by standard patrolling or station establishment.
We used only the detection/nondetection of a species at an individual survey point in order to minimize detection variability due to camera setup, ambient temperature, or body size. Manley and others (2004, 2005) proposed that a monitoring program employing presence/absence detections for multiple species is the most effective means to monitor biodiversity over a large scale. They proposed a monitoring program for the state of California based on rapid indexes of presence/absence at multiple sample locations. They considered a monitoring protocol to be effective if there are sufficient species’ detections and the ability to sample a large number of locations.
All indirect measures of mammal populations have shortcomings. Sight transects are limited by visibility, such as vegetation density, weather, and ambient light levels (Caro 1999). Care must be taken in sign surveys, as tracks are not consistent for all habitats (Grigione and others 1999) and droppings degrade at different rates depending on moisture, temperature, and the deposition site (Fuller 1991). Cameras have logistic problems of faulty settings, faulty batteries, and detection ranges (Swann and others 2004), but these problems are not influenced by the habitat if properly set up. Species-specific habitat associations are a common use of survey activity in reserves (Garshelis 2001; Gibbs 2001) and camera survey data (Moruzzi and others 2002). Our survey protocol could be improved by incorporating more detection probability theory (MacKensie 2005; Royal and Nichols 2003). The danger of single surveys is that failure to record a species can signify either absence or nondetection due to habitat or sampling attributes. We minimized habitat differences by limiting the range of the infrared sensor. Nevertheless, detection probability theory shows that repeat measures of detection/nondetection at a survey point can be used to estimate the probability of a false record of absence. With repeat surveys, it is possible to estimate how often the survey technique fails to detect a species that is truly present; repeat surveys can either be multiple surveys of the same sample point or a single survey divided into sequential periods. This second scenario would fit best within our protocol, with cameras left in place for 14 days, but our survey cameras did not reliably produce time and date stamps on pictures to allow such an analysis. As opposed to track or sign surveys, it is logistically easier for camera surveys to provide repeat measures because the technique would not depend on frequent returns by staff.
Effective measures of conservation need to be quantifiable, relatively rapid, and relevant to the threat (Ervin 2003; Parrish and others 2003). Some conservationists are advocating moving away from basing effectiveness on ground-based biological surveys because of their costs and slow speed (Green and others 2005; Parrish and others 2003). Population measures are difficult and time-consuming for most mammal species, but recent theoretical advances hold the potential of deriving population estimates from presence/absence data (Royle and Nichols 2003; Tosh and others 2004). Even with the retention of the survey as an indirect measure, the distribution of species relative to conservation infrastructure and activity is a rapid assessment of the necessity of these activities. Over the long term, monitoring does not need to rely on population estimates; it can rely on presence/absence detections (Manley and others 2004). Remote-trip cameras offer an indirect means to detect changes in species presence at sample points distributed throughout a system of reserves. For comparisons between reserves, their use would minimize reserve differences in reserve staff training or expertise, which is the main limitation of sign surveys (Wemmer and others 1996). Remote-trip cameras have limitations in detections and species identification, but these are not severe enough to preclude their use for presence/absence detections of broad suites of species, which is exactly what is needed to establish broad monitoring programs (Manley and others 2004). Effective management of reserves is desperately needed in China (Fu and others 2004) and quantitative measures of success would be one critical step in that direction.
References
Atauri J. A., J. V. De Lucio, M. A. Munoz. 2005. A framework for designing ecological monitoring programs for protected areas: a case study of Galachos del Ebro Nature Reserve (Spain). Environ Manage 35:20–33
Carbone E. S., K. Christie, T. Conforti, et al. 2001. The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim Conserv 4:75–79
Caro T. M. 1999. Conservation monitoring: Estimating mammal densities in woodland habitats. Anim Conserv 2:305–315
Carrillo E., G. Wong, A. D. Cuaron. 2000. Monitoring mammal populations in Costa Rican protected areas under different hunting restrictions. Conserv Biol 14:1580–1591
Ervin J. 2003. Rapid assessment of protected area management effectiveness in four countries. Bioscience 53:833–841
Foresman K. R., D. E. Pearson. 1998. Comparison of proposed survey procedures for detection of forest carnivores. J Wildl Manage 62:1217–1226
Fu B., K. Wang, Y. Lu, et al. 2004. Entangling the complexity of protected area management: The case of Wolong Biosphere reserve, Southwestern China. Environ Manage 33:788–798
Fuller T. K. 1991. Do pellet counts index white-tailed deer numbers and population change? J Wildl Manage 55:393–396
Gaines W. L. 2001. Large carnivore surveys in North Karakorum Mountains, Pakistan. Nat Areas J 21:168–171
Garshelis D. L. 2000. Delusions of habitat evaluation; measuring use, selection and importance. In L. Boitani, T. K. Fuller (eds), Research techniques in animal ecology; controversies and consequences. Columbia University Press, New York pp 111–164
Ge T. 2000. Preliminary study of the behavioral ecology of wild golden monkeys. In The progress of foundation. Tangjaihe Nature Reserve, Qingchuan County, Sichuan, People’s Republic of China (in Chinese with English abstract) pp 165–170
Gibbs J. P. 2000. Monitoring populations. In Boitani L, Fuller TK (eds), Research techniques in animal ecology; controversies and consequences. Columbia University Press, New York, pp 213–252
Green R. E., A. Balmford, P. R. Crane, et al. 2005. A framework for improved monitoring of biodiversity: Responses to the World Summit on Sustainable Development. Conserv Biol 19:56–65
Grigione M. M., P. Burman, V. C. Bleich, et al. 1999. Identifying individual mountain lions Felis concolor by their tracks: Refinement of an innovative technique. Biol Conserv 88:25–32
Hockings M. 2003. Systems for assessing the effectiveness of management in protected areas. Bioscience 53:823–832
Karanth K. U., J. D. Nichols. 1998. Estimation of tiger densities in India using photographic captures and recaptures. Ecology 79:2852–2862
Lindsey P. A., R. Alexander, J. T. Du Toit, et al. 2005. The cost efficiency of wild dog conservation in South Africa. Conserv Biol 19:205–1214
Ma S., X. Ma, W. Shi. 2001. A guide to mammal tracking in China. China Forestry Publishing House. Beijing
Mace G. M., J. E. M. Baillie, S. R. Beissinger, et al. 2001. Assessment and management of species at risk. In M. E. Soule, G. H. Orians (eds), Conservation biology; research priorities for the next decade. Island Press, Washington, DC pp 11–30
MacKensie D. I. 2005. What are the issues with presence–absence data for wildlife managers? J Wildl Manage 69:849–860
Manley P. N., M. D. Schlesinger, J. R. Roth. 2005. A field-based evaluation of a presence–absence protocol for monitoring ecoregional-scale biodiversity. J Wildl Manage 69:950–966
Manley P. N., W. J. Zielinski, M. D. Schlesinger. 2004. Evaluation of a multiple-species approach to monitoring species at the ecoregional scale. Ecol Appl 14:296–310
Margoluis R., N. Salafsky. 1998. Measures of success: Designing, managing and monitoring conservation and development projects. Island Press, Washington, DC
McNeely J. A. 1989. Protected areas and human ecology: How national parks can contribute to sustaining societies of the twenty-first century. In Western D, Pearl M (eds), Conservation for the twenty-first century. Oxford University Press, New York, pp 150–157
Milner-Gulland E. J., H. Resit Akcakaya. 2001. Sustainability indices for exploited populations. Trends Ecol Evol 16:686–691
Moruzzi T. L., T. K. Fuller, R. M. DeGraaf, et al. 2002. Assessing remotely triggered cameras for surveying carnivore distributions. Wildl Soc Bull 30:380–386
Nagendra H., C. Tucker, L. Carlson, J. Southworth, M. Karmacharya, B. Karna. 2004. Monitoring parks by remote sensing: Studies in Nepal and Honduras. Environ Manag 34:748–760
Numata S., T. Okuda, T. Sugimoto, et al. 2005. Camera trapping: a non-invasive approach as an additional tool in the study of mammals in Pasoh Forest Reserve and adjacent fragmented areas in Peninsular Malaysia. Malayan Nature J 57:29–45
Parrish J. D., D. P. Braun, R. S. Unnasch. 2003. Are we conserving what we say we are? Measuring ecological integrity within protected areas. Bioscience 53:851–860
Possingham H. P., S. J. Andelman, B. R. Noon, et al. 2001. Making smart conservation decisions. In M. E. Soule, G. H. Orians (eds), Conservation biology; research priorities for the next decade. Island Press, Washington, DC, pp 225–244
Rao M., Rabinowitz A., S. T. Khaing. 2002. Status review of the protected areas systems in Myanmar, with recommendation for conservation planning. Conserv Biol 16:360–368
Robinson J. G., R. E. Bodmer. 1999. Towards wildlife management in tropical forests. J Wildl Manage 63:1–13
Royle J. A., J. D. Nichols. 2003. Estimating abundance from repeated presence–absence data or point counts. Ecology 84:777–790
Sanderson J. G., M. Trolle. 2005. Monitoring elusive mammals; unattended cameras reveal secret of some of the world’s wildest places. Am Sci 93:148–155
Schaller G. B., Q. Teng, K. G. Johnson, et al. 2000. The feeding ecology of giant pandas and Asiatic black bears in the Tangjaihe Peserve, China. The progress of foundation. Tangjaihe Nature Reserve, Qingchuan County, Sichuan, People’s Republic of China, pp 235–255
Schauster E. R., E. M. Gese, A. M. Kitchen. 2002. An evaluation of survey methods for monitoring swift fox abundance. Wildl Soc Bull 30:464–477
Schwartman S., A. Moreira, D. Nepstad. 2000. Rethinking tropical forest conservation: Perils in parks. Conserv Biol 14:1351–1357
Sheng H., O. Noriyuki, H. Lu. 1999. The mammalian of China. China Forestry Publishing House, Beijing
Stem C., R. Margoluis, N. Salafsky, et al. 2005. Monitoring and evaluation in conservation: A review of trends and approaches. Conserv Biol 19:295–309
Swann D. E., C. A. Haas, D. C. Dalton, et al. 2004. Infrared-triggered cameras for detecting wildlife: An evaluation and review. Wildl Soc Bull 32:357–365
Tangjiahe NRAD (Nature Reserve Administrative Department). 2000. The progress of foundation. Tangjaihe Nature Reserve, Qingchuan County, Sichuan, People’s Republic of China
Tosh C. A., B. Reyers, A. S. van Jaarsveld. 2004. Estimating the abundances of large herbivores in the Kruger National Park using presence–absence data. Anim Conserv 7:55–61
Trolle M., M. Kery. 2003. Estimation of ocelot density in the Pantanal using capture–recapture analysis of camera-trapping data. J Mammal 84:607–614
Weckerly F. W., M. A. Ricca. 2000. Using presence of sign to measure habitats used by Roosevelt elk. Wildl Soc Bull 28:146–153
Wemmer C (ed). 1998. Deer; status survey and conservation action plan. IUCN/SSC Deer Specialist Group. IUCN, Gland, Switzerland
Wemmer C., T. H. Kunz, G. Lundie-Jenkins, et al. 1996. Mammalian Sign. In D. E. Wilson, F. R. Cole, J. D. Nichols, et al. (eds), Measuring and monitoring biological diversity; standard methods for mammals. Smithsonian Institution Press. Washington, DC pp 157–176
White L. J. T. 1994. Biomass of rain forest mammals in the Lope Reserve, Gabon. J Anim Ecol 63:499–512
Acknowledgments
This work was supported by Friend’s of the National Zoo, Peking University, Sichuan Forestry Department, Fuji Film, and China Wildlife Conservation Association. Daniel Guertin performed some of the preliminary analyses. The patrolling staff at Tangjiahe Nature Reserve performed the hard work of moving the cameras. Reviewers substantially improved the quality of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dajun, W., Sheng, L., McShea, W.J. et al. Use of Remote-Trip Cameras for Wildlife Surveys and Evaluating the Effectiveness of Conservation Activities at a Nature Reserve in Sichuan Province, China. Environmental Management 38, 942–951 (2006). https://doi.org/10.1007/s00267-005-0302-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00267-005-0302-3