
Abstract
Habitat selection has consequences for an animal’s fitness, especially for sit-and-wait predators with limited mobility, and which cannot always correct earlier suboptimal choices. Environmental change may nevertheless lead individuals to relocate to another site, although such relocations can be energetically costly or risky. Temperature and illumination are two important factors that undergo change in seasonal and daily cycles that may impact habitat quality. Animals must therefore either acclimate to the new conditions or relocate. Wormlions are sit-and-wait, trap-building predators whose success in foraging is highly dependent on their surroundings. Here, we manipulated temperature (high, low, and moderate) and let the wormlions choose between lit and shaded conditions. We found that the typical wormlion preference for shaded microhabitats decreased with increasing temperature. We then followed wormlion behavior under a full-factorial design of two constant illumination conditions (light vs. shade) and three temperatures. Although both constant light and high temperature reduced foraging performance, expressed in pit construction tendency and pit area, the two conditions had a non-additive effect. Acclimation to extreme thermal conditions moderated the negative effects of such temperatures, expressed in a higher tendency to construct a pit, and equalized performance across temperatures. Finally, the high temperature reduced behavioral consistency while acclimation increased it, suggesting that consistency is impaired by unfavorable environmental change. To conclude, while an environmental change usually affects several environmental factors simultaneously, the induced behavioral change is neither synergic nor additive and can even differ from the response to each unfavorable environmental factor in isolation.
Significance statement
Choosing a suitable habitat is essential for survival and reproduction, especially for sedentary organisms, and requires the consideration of various environmental conditions. Acclimating to suboptimal conditions, however, might conduce to improving performance in a less suitable habitat. Testing the effect of several environmental conditions on habitat choice and foraging behavior, before and after acclimation, has rarely been carried out. Here, we tested the combined effects of temperature, illumination, and their interaction, on habitat choice and foraging performance of a sit-and-wait predator, the wormlion. Wormlions usually prefer shade, but their preference for light increases with decreasing temperature. Both temperature and illumination affect behavior but their joint effect is not additive. Acclimation, which took place for temperature but not for illumination, improved certain foraging behaviors. Our findings highlight the importance of evaluating several environmental conditions and behaviors when studying habitat choice and foraging behavior.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Habitat selection, the behavioral process of selecting among various habitats (Gilliam and Fraser 1987; Morris 2003; Mohammadi 2010), is an important ecological process that influences both an individual animal’s fitness and population level phenomena (Bolnick et al. 2003; Bowler and Benton 2005; Wolf and Weissing 2012). For example, selecting the preferred nest sites relative to the non-preferred ones in respect to vegetation cover has been shown to increase nest success in birds (Martin 1998; Clark and Shutler 1999). However, spatial and temporal changes in the environment, such as temperature and nutrient availability, may force individuals to accommodate their responses to fit current conditions (Mettke-Hofmann et al. 2005; Kronfeld-Schor et al. 2013; Klokočovnik et al. 2016). Alternatively, response to environmental changes might lead to a reconsideration of habitat choice (Brown and Moyle 1991; Kotler et al. 1993; Ziv et al. 1993; Mettke-Hofmann et al. 2005; Yu et al. 2008). For example, over several years, nest site choice of birds and sea turtles shifts in response to changes in precipitation and artificial light, respectively (Martin 2001; Salmon 2003).
Temperature and illumination change in seasonal and daily cycles. Many species respond quickly to such changes. For example, the tempo of ovarian recrudescence in catfish is higher at moderate relative to unfavorably low or high temperatures (Sundararaj and Vasal 1976). Furthermore, between two species of stream fish, in which one preys on the other, light intensity affects predation risk: under bright light, predators seek refuge while the prey species is free to aggregate and be active (Cerri 1983). Temperature and illumination clearly affect activity and other diel and seasonal patterns (Brodie and Russell 1999; Longcore and Rich 2004; Biro et al. 2010), which are responsible for foraging success and ultimately, reproduction and survival (Kavanau 1969; Cerri 1983; Gaston et al. 2013). These two factors thus play a major role in habitat choice across taxa (Herberstein and Fleisch 2003; Scharf et al. 2008; Dubois et al. 2009; Halliday and Blouin-Demers 2014).
When animals select a habitat, this choice affects their ability to function adequately (Huey 1991), according to their innate thermal preference (Angilletta 2009, ch. 3). Animals, however, may adapt to environmental change following exposure through acclimation, defined as modifications in behavior and physiology that improve individual performance under unfavorable climatic conditions (Bowler 2005; Angilletta 2009, ch. 5). There are numerous examples of acclimation (e.g., Wilson et al. 2000; Geister and Fischer 2007; Gvoždik et al. 2007). Desert lizards, for example, shift their thermal maximum and minimum following acclimation to higher and lower temperatures, respectively (Kaufmann and Bennett 1989).
While animals may respond to changes in temperature by acclimation, response to changes in illumination may be expressed through entrainment. Entrainment of circadian rhythms constitutes the synchronization of the endogenous clock to the day/night cycle as a result of environmental cues, such as illumination and temperature (Pittendrigh 1993; Longcore and Rich 2004). Under natural conditions, changes in temperature match changes in day length, giving rise to seasonality—colder during short days and warmer during long days (Coppack and Pulido 2004; Bradshaw and Holzapfel 2007). Thus, the response to temperature and illumination should be correlated. For example, long days lead to a preference for higher temperature in turtle species (Graham and Hutchison 1979). Illumination can also change interspecific interactions, such as predator-prey interactions. Light pollution and a full moon lead both to a decrease in activity of nocturnal rodent prey and an increase in the activity of their predators (Kotler et al. 1991; Gaston et al. 2013).
Some environmental changes might alter habitat characteristics, which in turn may threaten the organism’s homeostasis. In response to such unfavorable changes, individuals might be forced to relocate from their ecological niche (Pearson and Dawson 2003; Steinberg 2012, ch. 1). Furthermore, if such conditions alter fast or drastically, they may not only affect behavior but may also impact behavioral consistency (Sih et al. 2004; Dingemanse and Wolf 2013). For example, starvation impairs consistency concerning aggression, boldness, and web location choice in spiders (Pruitt et al. 2011a, b; Lichtenstein et al. 2016; but see Stahlschmidt et al. 2014). Similarly, higher phenotypic variance is induced by unfavorable or unpredictable environments (Köhler et al. 2009).
Consistency might conceivably restrict an individual’s ability to cope with environmental change (Pruitt et al. 2011a, b). The effect of unfavorable environmental conditions on behavioral consistency has received little attention and a pattern of change in behavioral consistency across an environmental gradient of unfavorable vs. favorable conditions remains to be determined. Behavioral consistency is a trait in its own right that differs among individuals, similar to many other behavioral traits (Bell et al. 2009; Pruitt et al. 2011a, b). Habitat choice, for example, varies consistently among individuals, with different individuals demonstrating different balance points between conflicting demands, which can result in situations in which some individuals appear to prefer suboptimal habitats (Spencer and Thompson 2003; Katz et al. 2016).
We examined here the effect of illumination, temperature, and thermal acclimation on microhabitat choice and foraging performance of a trap-building predator, the wormlion larva (Diptera: Vermileonidae). Trap-building predators (mostly spiders, antlions, and wormlions) are limited in their mobility and depend on local environmental conditions for the construction of their trap, without which their prey capture drastically diminishes (Heinrich and Heinrich 1984; Arnett and Gotelli 2001; Dor et al. 2014). This leads to a preference for specific microhabitats (e.g., specific substrate and illumination; Adams 2000; Devetak 2008; Pruitt et al. 2011a, b; Devetak and Arentt 2015; Adar et al. 2016). In their natural habitat, wormlions are solely found under sheltered conditions (either vegetation cover or beneath buildings), where they are protected from direct sunlight. Furthermore, when given a choice between illuminated and shaded microhabitats, wormlions strongly and consistently prefer shade (Adar et al. 2016; Katz et al. 2016). Exposure to light and high temperatures are disfavored by wormlions as they entail the risk of desiccation, and lit habitats are therefore probably considered by wormlions as unsuitable for settlement.
We tested here (1) whether temperature affects wormlion preference for shaded microhabitats. We expected that the preference for shade would be higher at high temperatures because these can indicate an ongoing ground heating. This response could be interpreted as an attempt to regulate the later negative effects associated with potential desiccation; (2) whether performance, reflected in pit characteristics (pit construction tendency and area), differs with temperature and illumination. We expected that performance would be maximized under shade and at a moderate temperature; (3) whether acclimation to specific temperatures affects performance, with an expected improvement following acclimation leading to similar levels of performance across treatments; and (4) whether behavioral consistency is affected by illumination, temperature, and thermal acclimation. We expected consistency to be lower under light and at the low and high temperatures tested, leading to an inverse U-shaped pattern of consistency with temperature change. Acclimation was expected to moderate existing differences in consistency levels among treatments, for which, following acclimation, similar levels of behavioral consistency across treatments were expected.
Material and methods
Model organism
Wormlion larvae (Diptera: Vermileonidae) are trap-building predators that capture small arthropods in the cone-shaped pits that the larvae dig in loose soil (Wheeler 1930; Devetak 2008). They spend most of their life cycle in the larval stage, which usually lasts up to a year. Following pupation, a weak-flying and short-lived adult emerges (Wheeler 1930). Wormlions are found at high densities and are abundant in urban areas featuring thin layers of loose soil and shaded conditions, often near buildings (Dor et al. 2014; Scharf and Dor 2015; Devetak and Arentt 2015). Their strong preference for shaded conditions also appears under laboratory conditions, with most individuals consistently selecting shaded over illuminated conditions (Adar et al. 2016; Katz et al. 2016).
Two experiments were conducted to evaluate how temperature, illumination, and their interaction would influence the wormlion preference for shaded conditions, their performance, reflected in pit construction and pit area, and their behavioral consistency. The first experiment examined how larval microhabitat preference for illumination conditions, pit performance, and behavioral consistency were influenced by different temperatures. The second experiment examined how pit construction tendency, pit area, and behavioral consistency were influenced by thermal acclimation (tested before and after acclimation), temperature (high = 37 °C, moderate = 26 °C, and low = 15 °C), and illumination conditions (light and shade). All national guidelines for the collection, care, and use of insects were followed. Following the experiments, wormlions were released back into their original habitat.
Experiment 1: the effect of temperature on shade preference
Preparations for the experiment
Wormlion larvae (n = 168; body mass = 0.0272 ± 0.0275 g; mean ± 1 SD) were collected from sandy patches at Tel Aviv University in November 2016. The larvae were of different ages and sizes but were evenly distributed across treatments. Furthermore, in wormlions, body mass was previously observed to either have no or only weak correlation with pit performance (Dor et al. 2014). Sex could not be determined at the larval stage. Immediately after collection, larvae were allocated to one of three similar mass groups of 56 individuals. Each larva was kept in an individual cup (4.5-cm diameter) filled with 1.5-cm-deep filtered and homogenized fine sand (particle size <106 mm). To promote habituation to the experimental conditions, larvae were kept undisturbed inside climate chambers (under specific air temperature of 15, 26, or 37 °C, light/dark cycle of 12:12 h), for 5 days prior to the beginning of the experiment. Sand temperature was not measured but was plausibly similar to air temperature due to its low volume and long maintenance in the climate chamber. To standardize the hunger level, each wormlion larva was fed with one similar-sized flour beetle larva (Tribolium castaneum) 3 days after collection.
Experimental set-up
During the experiment, the climate chambers were constantly illuminated and larvae were kept in aluminum trays (30 × 10 cm) filled with 1.5-cm-deep fine sand. Each tray was half-covered with an aluminum lid, creating two similar-sized sections: full shade and illuminated. Larvae were placed in the center of the tray, between the two illumination conditions. Wormlion behavior was followed by photographing individual trays every 24 h for three consecutive days. Photos were taken between 8:00–10:00 a.m. To minimize bias when analyzing the photos, the observer was blind to the treatment.
Following each photograph, larvae were placed back in the center of the tray, allowing them to re-choose either shade or light. For each of the 3 days, four response variables were documented: (1) Light/shade preference, preference for either the illuminated or shaded side of the tray, or no preference. (2) Location relative to shade, the ratio between the larva’s distance from the tray’s fully shaded edge relative to the tray’s length (values smaller than 0.5 represent preference for full shade). (3) Pit construction, whether a pit was built or not. (4) Pit area, the area bounded by the pit edges, only for larvae constructing pits. Behavioral consistency was evaluated by comparing each trait between days and testing for correlation or agreement of choice (“level of agreement” was defined as choosing the same illumination conditions and as constructing pits on consecutive days or not constructing a pit on both days). Location relative to shade and pit area were measured using ImageJ (Abràmoff et al. 2004).
Experiment 2: the effect of acclimation, temperature, and illumination on behavior
Preparations for the experiment
Wormlion larvae (n = 300; body mass = 0.049 ± 0.036 g; mean ± 1 SD) were collected in November–December 2016. Following collection, the larvae were assigned to six similar-mass groups of 50 individuals each (each group received a combination of three temperatures × two illumination conditions). Each larva was kept in a cup (8-cm diameter) filled with 1.5-cm-deep fine sand. Cups were used rather than trays since only pit characteristics were measured, for which cups are sufficient. Furthermore, cups were preferred here due to their small size, which enabled us to maintain a large sample. Cups were then placed in a climate chamber at 15, 26, or 37 °C under either constant light or shade conditions. The experiments started 24 h after collection.
Experimental set-up
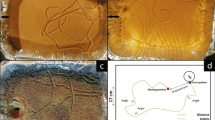
The experiment comprised six treatments: three temperatures × two illumination conditions (Fig. 1). For each treatment, six photographs were taken on three consecutive days before and after thermal acclimation. For each photo, pit construction and area were calculated. Thermal acclimation lasted 5 days under 12:12 L/D, to mimic natural conditions. During each stage, wormlion behavior was followed by photographing the individual cups. After photographing, each larva was placed back in the center of the cup, allowing it to reconstruct a pit. Pit construction and area were documented similarly to the first experiment.

Scheme of experiment 2 (The effect of acclimation, temperature, and illumination on behavior). Behavior was measured during two stages, before and after the 5-day acclimation period. During each stage, we photographed the wormlions using a digital camera every 24 h for three consecutive days (a total of 6 days of observation)
Statistical analyses
Experiment 1
A χ 2 test was used on light/shade preference and pit construction to determine whether either response variable (illumination preference or pit construction tendency and area) were affected by temperature. Pairwise comparisons between temperatures were computed by χ 2 tests, corrected for multiple testing (Bonferroni). The effect of temperature on pit location relative to shade and pit area was tested by two one-way ANOVAs (followed by Bonferroni post hoc tests). For the categorical response variables, three tests were performed, one for each day, while for the continuous variables one test on each individual’s averages over the 3-day test was performed.
A χ2 test was used to determine whether the level of agreement in light/shade preference and pit construction between consecutive days differed between temperatures. When significant, multiple comparisons were performed, as explained above. Intra-class correlation (hereafter, ICC) was used to evaluate the level of consistency across all 3 days in pit location relative to shade and pit area for each temperature. For analysis of the categorical variables, only individuals that had responded by either choosing or constructing a pit on at least 1 day, were included. For the analysis of the continuous variables, only individuals that had responded on all 3 days were included. An arcsine transformation was used for location relative to shade and the square-root transformation for pit area. During the experiment, 23 individuals died, with a similar representation of mortality events across temperatures (see Supplementary Material, Table S1).
Experiment 2
For pit construction, the data of the first day were similarly analyzed separately, before and after acclimation. Two log-linear tests were used with temperature and illumination conditions as explanatory variables and pit construction as the response variable. For pairwise comparisons of temperatures, χ 2 tests following a Bonferroni correction were used. The second and third days showed similar trends but are not presented here. Next, a repeated-measures ANOVA was used to test for differences in average pit area (averaged across the three consecutive days before and after acclimation). Acclimation was referred to as the within-subject variable and temperature and light/shade condition as the between-subject variables. Paired t tests were performed to evaluate differences within each temperature before and after acclimation (Bonferroni corrected). For this analysis, only individuals that had constructed a pit on at least 1 day out of the 3 days in both time periods (before and after acclimation) were included.
Two log-linear tests were used to analyze the level of agreement in pit construction between days 1 and 2, before and after acclimation. The only significant effect was that of temperature (see “Results”); separate χ 2 tests were then used for pairwise comparisons among temperatures (Bonferroni corrected). A χ 2 test was used to determine whether the level of agreement in pit construction between consecutive day pairs 1–2 and 2–3 differed between temperatures before and after acclimation. ICC (intra-class correlation) test was used to evaluate the level of consistency in pit area for each combination of temperature, illumination conditions, and acclimation. Pit area was square root transformed for this analysis. During the experiments 67 individuals died, with significantly more deaths occurring at 37 °C and under light (see analysis in the Supplementary Material, Tables S2, S3).
Results
Experiment 1: The effect of temperature on shade preference
Light/shade preference and pit construction
Over all treatments, wormlions mostly preferred shade over light (Fig. 2a). The tendency to either choose light or not to choose at all (i.e., constructing a pit in the center of the tray where the larva was initially placed) was higher at the low and high temperatures, respectively (χ 2 = 10.74, df = 4, P = 0.029 and χ 2 = 18.55, df = 4, P < 0.001, for days 2 and 3, respectively; day 1 was not significant: χ 2 = 7.54, df = 4, P = 0.110; see Supplementary Material, Fig. S1a, for the results of day 2). Multiple comparisons indicated that on day 2, the wormlions’ tendency to choose light differed between the low and high temperatures (P < 0.005). On day 3, the high temperature resulted in the tendency to choose light differing from that under the two other temperatures (P < 0.007). The tendency to construct a pit was much lower at the high temperature relative to the two other temperatures (χ 2 = 12.33, df = 2, P = 0.002 and χ2 = 27.83, df = 2, P < 0.0001, for days 2 and 3, respectively; day 1 was not significant: χ 2 = 1.97, df = 2, P = 0.374; Fig. 2b; see Supplementary Material, Fig. S1b, for the results of day 2). Multiple comparisons revealed that on day 2, the tendency to construct a pit at the high temperature significantly differed from that at the moderate temperature (P < 0.001), and on day 3 the tendency under high temperature differed from that under the two other temperatures (P < 0.0001 for both).

Experiment 1: Differences in a the proportions of illumination choice, and b pit construction between temperatures on the third day of the experiment. The tendencies to choose light or not to choose at all were higher at the low and high temperatures, respectively. The choice of light and the tendency to construct a pit at the high temperature was lower than at the two other temperatures (low and moderate). Letters denote significant differences among temperatures in the proportion of choice or pits
Location relative to shade and pit area
Temperature did not affect the location relative to shade (F 2,84 = 2.17, P = 0.120) but did affect pit area (F 2,68 = 5.73, P = 0.005). Multiple comparisons revealed that at the moderate temperature, larvae constructed larger pits than at the low temperature (P = 0.005).
Differences in consistency
Levels of agreement in light/shade preference between temperatures on both consecutive day pairs (1–2 and 2–3) did not differ (Table 1). However, the level of agreement in pit construction differed between temperatures between days 2–3 (Table 1), although these differences were solely between the high and the moderate temperatures (P = 0.009). At high temperature, agreement was much lower compared to the moderate temperature. Location relative to shade was consistent only at the low temperature across all three days (P = 0.022), while pit area was consistent at all three temperatures (P < 0.0001 for all; Table 2).
Experiment 2: the effect of acclimation, temperature, and illumination on behavior
Pit construction
The log-linear model indicated that temperature and illumination interacted to influence pit construction tendency before acclimation (χ2 = 6.25, df = 2, P = 0.044): at 37 °C more pits were constructed under light vs. shade, while the opposite held true at 15 and 26 °C (Fig. 3a). Regarding the main effects, illumination was not significant (χ 2 = 1.360, df = 1, P = 0.244), but temperature was significant with fewer pits constructed at 37 °C than at the two other temperatures (X2 = 6.69, P = 0.010). Following acclimation, the temperature-illumination interaction was not significant (χ 2 = 2.77, df = 2, P = 0.250). Illumination and temperature as main effects both had a significant effect (χ 2 = 10.87, df = 1, P < 0.001 and χ 2 = 54.52, df = 2, P < 0.0001, respectively; Fig. 3b). Individuals constructed more pits under shade than light (Fig. 4a), more pits at the moderate temperature than the low temperature, and more pits at the low temperature than the high temperature (P < 0.002 for all; Fig. 4b).

Experiment 2: Differences in the proportions of pit construction between temperatures and illumination conditions on the first day a before, and b after thermal acclimation. Before and after acclimation, the tendency to construct a pit was highest at the moderate temperature followed by the low temperature, and lowest at the high temperature regardless of illumination conditions. Shade had a positive effect on pit construction tendency, except for the high temperature before acclimation

Experiment 2: Differences in the proportions of pit construction a between light and shade conditions, and b between temperatures, before and after acclimation. The tendency to construct a pit was higher under shade relative to light conditions, but only after acclimation. There were differences between temperatures in the tendency to construct a pit before and after acclimation. The highest tendency to construct a pit was at the moderate temperature, and the lowest at the high temperature
Pit area
Pit area was affected by illumination condition and temperature but not by their interaction (Table 3). Larvae built larger pits under shaded conditions than under lit ones (Fig. 5a). A post hoc Bonferroni test revealed that pit area at moderate temperature was greater than at the two other temperatures (P < 0.012). Furthermore, larvae constructed smaller pits following acclimation. However, there was a significant interaction between acclimation and temperature. At the high and moderate temperatures, acclimation led to a decrease in pit area, while at the low temperature, there was no significant change (Fig. 5b).

Experiment 2: Differences in average pit area (± 1 SE) between a light and shade conditions, and b between temperatures before and after acclimation. Average pit area was higher under shade relative to light conditions. At the moderate temperature larvae constructed larger pits relative to the two other temperatures (Bonferroni, P < 0.0001 and P = 0.012 between the moderate and the low and high temperatures, respectively). Average pit area decreased following acclimation at the high and moderate temperatures, but did not change at the low temperature. Letters denote significant differences among temperatures in average pit area. Asterisks denote differences within a temperature before and after acclimation
Differences in consistency
The log-linear model indicated that temperature and illumination did not interact to influence pit construction (day pair 1–2) before acclimation (χ 2 = 2.36, df = 2, P = 0.308). The only significant effect was that of temperature (χ 2 = 33.41, df = 2, P < 0.0001): consistency level at the high temperature was lower than at the two other temperatures (P < 0.0001 for both). Illumination too was not significant (χ 2 = 0.054, df = 1, P = 0.817). Following acclimation, the interaction was not significant (χ 2 = 4.20, df = 2, P = 0.123) and illumination had no effect (χ 2 = 0.51, df = 1, P = 0.473). The only significant effect was that of temperature (χ 2 = 13.25, df = 2, P = 0.001): consistency level at the high temperature was lower than at the two other temperatures (P < 0.007 for both). The level of agreement in pit construction between consecutive day pairs (1–2 and 2–3) differed between temperatures regardless of illumination conditions before but not after acclimation, except for day pair 1–2 under shade conditions, which was also significant following acclimation (Table 4). All three temperatures showed significant consistencies in pit area between all 3 days both before and after acclimation and under the two illumination conditions. However, as the high-temperature test contained too low a sample size, its results preclude interpretation (Table 5).
Discussion
As strict sit-and-wait predators with low mobility, wormlions are dependent on their immediate surroundings and strongly respond to microhabitat change. In the above experiments, temperature and light interacted to affect wormlion preference for the shaded microhabitat: although most wormlions selected shade over light, the tendency to choose light was higher at lower temperatures. When kept under constant conditions, there was no interaction between temperature and light, and an independent increase in either one of them decreased pit construction tendency and area. Acclimation interacted with temperature, decreasing pit area dug for the two higher temperatures, but did not lead to a change in area in response to illumination conditions. Finally, behavioral consistency was lower at the high temperature, irrespective of illumination conditions, while acclimation increased behavioral consistency overall.
Low temperature increased wormlion preference for the usually non-preferred lit microhabitat, following our expectations. This response may be adaptive, as the negative effects of too low a temperature can be moderated by the opportunity to warm up under direct sunlight. This suggests that wormlions adjust their exposure to light in order to reach some optimal temperature, which remains to be discovered. In turtles, Graham and Hutchison (1979) demonstrated an inverse relationship, in which a longer photoperiod led to a higher temperature preference, explained as adaptation to seasonal spring warming. Such a relationship between temperature and illumination might suggest that wormlions possess some sort of seasonal sensitivity, responding adaptively to cooling temperatures in winter by tolerating direct sunlight more than in summer (Bradshaw and Holzapfel 2007). Since illumination and temperature are correlated in nature, illumination preference could be a by-product of either temperature preference or a thermal constraint. It could also be linked to a third factor, such as humidity level, which is also correlated with temperature and illumination. A future study could use a design of changing illumination levels and then test for the preferred temperature. Examining wormlion behavior under an extreme temperature and illumination regime in the field should be carried out in order to determine whether the responses observed in the laboratory indeed reflect those occurring in nature.
Following our prediction, wormlions constructed more and larger pits at moderate temperature and shade conditions, which we interpret to be the optimal conditions for this species. This is based on the assumption that maximal performance is achieved under such conditions (Huey 1991; Angilletta 2009, ch. 3). The tendency to construct a pit was the lowest at the high temperature, both under constant illumination conditions and when choice was allowed. The high temperature could have led to high activity cost and desiccation risk. Despite the natural association between temperature and day length (Coppack and Pulido 2004; Bradshaw and Holzapfel 2007), the simultaneous exposure to high temperature and constant light did not generate a greater, additive negative effect on performance. We interpret this finding as the result of our unnatural illumination regime. Mortality rate was higher at the light and high-temperature treatments relative to shade and the other temperatures, but the effect of a combination of high temperature and light was no greater than that of each of these two in isolation. This finding complements those for the pit construction and area patterns, in which high temperature and light were both unsuitable but there was usually no additive effect.
Following acclimation, more pits were constructed at the highest temperature. This suggests that thermal acclimation improved performance, and the wormlions’ response therefore corresponds to the definition of acclimation (Bowler 2005). Pit area at both moderate and high temperatures decreased following acclimation, reaching similar pit areas at all temperatures. This could be interpreted as additional evidence of acclimation, indicating that wormlions might thus achieve an “average pit size,” irrespective of temperature. Acclimation did not interact with illumination. It is possible that our constant illumination conditions were too extreme to enable fast acclimation, whereas temperature fluctuates on a more daily basis than photoperiod, facilitating daily acclimation (Overgaard and Sørensen 2008). On the other hand, conditions that consistently change (light/dark) might lead to lower levels of acclimation, because the cue prior to acclimation is not reliable (based on the comparison between acclimation in terrestrial and aquatic habitats which are less and more thermally stable, respectively; Wilson and Johnston 2000). The wormlions’ natural habitat conditions are fairly stable, but more stable concerning illumination than temperature, due to the strong preference of wormlions for shade (Dor et al. 2014; Devetak and Arnett 2015). This might explain the absence of their acclimation to illumination.
Consistency of pit construction was lower at the high temperature across all other conditions. This supports our prediction of low behavioral consistency under unfavorable conditions, similar to the elevated phenotypic divergence induced by stress (Köhler et al. 2009). The pattern, however, did not resemble an inverse U-shape but, rather, a sharp decline at the high temperature. It is possible that the lower temperature was not perceived as unfavorable, because 15 °C is a common winter temperature in the wormlions’ habitat of origin. Furthermore, such a low temperature does not involve desiccation risk, which might represent the most serious danger at high temperatures. Future studies may consider reducing the lower temperature further and examining whether this has any effect on consistency.
Consistency of pit area was always high, except for the high temperature and constant illumination. This latter lack of consistency could have stemmed from the low number of constructed pits, making this result not indicative of true behavioral consistency. Similarly, extreme temperatures limited the ability of a lizard species to right itself and prevented the evaluation of locomotor performance (Angilletta and Robson 2002). The specific combination of constant illumination and high temperature might have caused a synergic negative effect on the behavior and consistency of some foraging activity by the wormlions. A synergism of two factors or more is common in nature, such as an exposure to toxins and thermal stress (Holmstrup et al. 2010). Microhabitat choice in our first experiment was measured in two ways: categorical location (light vs. shade) and exact location relative to shade. These two variables demonstrated different consistency levels among temperatures. Categorical location was similarly consistent across temperatures, but location relative to shade was consistent only at the low temperature. The latter result supports our prediction that behavioral consistency would decrease with temperature. The differences between the two variables, categorical location and location relative to shade, could have stemmed from the exclusion of individuals that did not move at all from their initial position in location relative to shade, which reduced consistency.
In summary, we demonstrate here the combined effect of several environmental factors on wormlion behavior and the interactions among them. We see the three most important findings as: (1) wormlions seek to compensate for low temperature by increasing their preference for lit microhabitats; (2) thermal acclimation improves pit construction rate but leads to a decrease in pit area. Consequently, when studying acclimation, several possible responses should be measured to accurately indicate its effect; (3) the consistency of wormlion behavior dropped at the unfavorable high temperature, and it remains to be determined whether consistency in general decreases with an increase in the gradient of the unfavorable condition. The synergism between unfavorable environmental factors is important to predict an animal’s responses to environmental change. Climate change, for example, might involve both increasing temperature, increase in the frequency of rare events (e.g., heat waves), and a decrease in precipitation (Easterling et al. 2000; Walther et al. 2002). It is thus important to study several effects in parallel, with the response, as found here, not necessarily always being additive.
References
Abràmoff MD, Magalhães PJ, Ram SJ (2004) Image processing with ImageJ. J Biophotonics Int 11:36–42
Adams MR (2000) Choosing hunting sites: web site preferences of the orb weaver spider, Neoscona crucifera, relative to light cues. J Insect Behav 13:299–305
Adar S, Dor R, Scharf I (2016) Habitat choice and complex decision making in a trap-building predator. Behav Ecol 27:1491–1498
Angilletta MJ (2009) Thermal adaptation: a theoretical and empirical synthesis. Oxford University Press
Arnett AE, Gotelli NJ (2001) Pit-building decisions of larval ant lions: effects of larval age, temperature, food, and population source. J Insect Behav 14:89–97
Bell AM, Hankison SJ, Laskowski KL (2009) The repeatability of behavior: a meta-analysis. Anim Behav 77:771–783
Biro PA, Beckmann C, Stamps JA (2010) Small within-day increases in temperature affects boldness and alters personality in coral reef fish. Proc R Soc B 277:71–77
Bolnick DI, Svanbäck R, Fordyce JA, Yang LH, Davis JM, Hulsey CD, Forister ML (2003) The ecology of individuals: incidence and implications of individual specialization. Am Nat 161:1–28
Bowler DE, Benton TG (2005) Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biol Rev 80:205–225
Bowler K (2005) Acclimation, heat shock and hardening. J Therm Biol 30:125–130
Bradshaw WE, Holzapfel CM (2007) Evolution of animal photoperiodism. Annu Rev Ecol Evol Syst 38:1–25
Brodie ED, Russell NH (1999) The consistency of individual differences in behaviour: temperature effects on antipredator behaviour in garter snakes. Anim Behav 57:445–451
Brown LR, Moyle PB (1991) Changes in habitat and microhabitat partitioning within an assemblage of stream fishes in response to predation by Sacramento squawfish (Ptychocheilus grandis). Can J Fish Aquat Sci 48:849–856
Cerri RD (1983) The effect of light intensity on predator and prey behaviour in cyprinid fish: factors that influence prey risk. Anim Behav 31:736–742
Clark RG, Shutler D (1999) Avian habitat selection: pattern from process in nest-site use by ducks? Ecology 80:272–287
Coppack T, Pulido F (2004) Photoperiodic response and the adaptability of avian life cycles to environmental change. Adv Ecol Res 35:131–150
Devetak D (2008) Substrate particle size-preference of wormlion Vermileo vermileo (Diptera: Vermileonidae) larvae and their interaction with antlions. Eur J Entomol 105:631–635
Devetak D, Arentt AE (2015) Preference of antlion and wormlion larvae (Neuroptera: Myrmeleontidae; Diptera: Vermileonidae) for substrates according to substrate particle sizes. Eur J Entomol 112:500–509
Dingemanse NJ, Wolf M (2013) Between-individual differences in behavioural plasticity within populations: causes and consequences. Anim Behav 85:1031–1039
Dor R, Rosenstein S, Scharf I (2014) Foraging behaviour of a neglected pit-building predator: the wormlion. Anim Behav 93:69–76
Dubois Y, Blouin-Demers G, Shipley B, Thomas D (2009) Thermoregulation and habitat selection in wood turtles Glyptemys insculpta: chasing the sun slowly. J Anim Ecol 78:1023–1032
Easterling DR, Meehl GA, Parmesan C, Changnon SA, Karl TR, Mearns LO (2000) Climate extremes: observations, modeling, and impacts. Science 289:2068–2074
Gaston KJ, Bennie J, Davies TW, Hopkins J (2013) The ecological impacts of nighttime light pollution: a mechanistic appraisal. Biol Rev 88:912–927
Geister TL, Fischer K (2007) Testing the beneficial acclimation hypothesis: temperature effects on mating success in a butterfly. Behav Ecol 18:658–664
Gilliam JF, Fraser DF (1987) Habitat selection under predation hazard: test of a model with foraging minnows. Ecology 68:1856–1862
Graham TE, Hutchison VH (1979) Effect of temperature and photoperiod acclimatization on thermal preferences of selected freshwater turtles. Copeia 1979:165–169
Gvoždik L, Puky M, Šugerkova M (2007) Acclimation is beneficial at extreme test temperatures in the Danube crested newt, Triturus dobrogicus (Caudata, Salamandridae). Biol J Linn Soc 90:627–636
Halliday WD, Blouin-Demers G (2014) Red flour beetles balance thermoregulation and food acquisition via density-dependent habitat selection. J Zool 294:198–205
Heinrich B, Heinrich MJ (1984) The pit-trapping foraging strategy of the ant lion, Myrmeleon immaculatus DeGeer (Neuroptera: Myrmeleontidae). Behav Ecol Sociobiol 14:151–160
Herberstein ME, Fleisch AF (2003) Effect of abiotic factors on the foraging strategy of the orb-web spider Argiope keyserlingi (Araneae: Araneidae). Austral Ecol 28:622–628
Holmstrup M, Bindesbøl AM, Oostingh GJ et al (2010) Interactions between effects of environmental chemicals and natural stressors: a review. Sci Total Environ 408:3748–3762
Huey RB (1991) Physiological consequences of habitat selection. Am Nat 137:91–115
Katz N, Subach A, Pruitt JN, Scharf I (2016) Habitat preference of wormlions and their behavioural repeatability under illumination/shade conditions. Ecol Entomol 41:716–726
Kaufmann JS, Bennett AF (1989) The effect of temperature and thermal acclimation on locomotor performance in Xantusia vigilis, the desert night lizard. Physiol Zool 62:1047–1058
Kavanau JL (1969) Influences of light on activity of small mammals. Ecology 50:548–557
Klokočovnik V, Hauptman G, Devetak D (2016) Effect of substrate temperature on behavioural plasticity in antlion larvae. Behaviour 153:31–48
Köhler HR, Lazzara R, Dittbrenner N, Capowiez Y, Mazzia C, Triebskorn R (2009) Snail phenotypic variation and stress proteins: do different heat response strategies contribute to Waddington's widget in field populations? J Exp Zool Part B 312:136–147
Kotler BP, Brown JS, Hasson O (1991) Factors affecting gerbil foraging behavior and rates of owl predation. Ecology 72:2249–2260
Kotler BP, Brown JS, Subach A (1993) Mechanisms of species coexistence of optimal foragers: temporal partitioning by two species of sand dune gerbils. Oikos:548–556
Kronfeld-Schor N, Dominoni D, de la Iglesia H, Levy O, Herzog ED, Dayan T, Helfrich-Forster C (2013) Chronobiology by moonlight. Proc R Soc Lond 280:20123088
Lichtenstein JL, DiRienzo N, Knutson K et al (2016) Prolonged food restriction decreases body condition and reduces repeatability in personality traits in web-building spiders. Behav Ecol Sociobiol 70:1793–1803
Longcore T, Rich C (2004) Ecological light pollution. Front Ecol Environ 2:191–198
Martin TE (1998) Are microhabitat preferences of coexisting species under selection and adaptive? Ecology 79:656–670
Martin TE (2001) Abiotic vs. biotic influences on habitat selection of coexisting species: climate change impacts? Ecology 82:175–188
Mettke-Hofmann C, Ebert C, Schmidt T, Steiger S, Stieb S (2005) Personality traits in resident and migratory warbler species. Behaviour 142:1357–1375
Mohammadi S (2010) Microhabitat selection by small mammals. Adv Biol Res 4:283–287
Morris DW (2003) Toward an ecological synthesis: a case for habitat selection. Oecologia 136:1–13
Overgaard J, Sørensen JG (2008) Rapid thermal adaptation during field temperature variations in Drosophila melanogaster. Cryobiology 56:159–162
Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob Ecol Biogeogr 12:361–371
Pittendrigh CS (1993) Temporal organization: reflections of a Darwinian clock-watcher. Annu Rev Physiol 55:17–54
Pruitt JN, Demes KW, Dittrich-Reed DR (2011b) Temperature mediates shifts in individual aggressiveness, activity level, and social behavior in a spider. Ethology 117:318–325
Pruitt JN, DiRienzo N, Kralj-Fišer S, Johnson JC, Sih A (2011a) Individual-and condition-dependent effects on habitat choice and choosiness. Behav Ecol Sociobiol 65:1987–1995
Salmon M (2003) Artificial night lighting and sea turtles. Biologist 50:163–168
Scharf I, Hollender Y, Subach A, Ovadia O (2008) Effect of spatial pattern and microhabitat on pit construction and relocation in Myrmeleon hyalinus (Neuroptera: Myrmeleontidae) larvae. Ecol Entomol 33:337–345
Scharf I, Dor R (2015) The effects of starvation and repeated disturbance on mass loss, pit construction, and spatial pattern in a trap-building predator. Ecol Entomol 40:381–389
Sih A, Bell A, Johnson JC (2004) Behavioral syndromes: an ecological and evolutionary overview. Trends Ecol Evol 19:372–378
Spencer RJ, Thompson MB (2003) The significance of predation in nest site selection of turtles: an experimental consideration of macro-and microhabitat preferences. Oikos 102:592–600
Stahlschmidt Z, O’Leary ME, Adamo S (2014) Food limitation leads to risky decision making and to tradeoffs with oviposition. Behav Ecol 25:223–227
Steinberg CEW (2012) Stress ecology: environmental stress as ecological driving force and key player in evolution. Springer, Heidelberg
Sundararaj BI, Vasal S (1976) Photoperiod and temperature control in the regulation of reproduction in the female catfish Heteropneustes fossilis. J Fish Board Can 33:959–973
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJ, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Wheeler WM (1930) Demons of the dust. Norton & Company Inc Publishers, New York
Wilson RS, James RS, Johnston IA (2000) Thermal acclimation of locomotor performance in tadpoles and adults of the aquatic frog Xenopus laevis. J Comp Physiol B 170:117–124
Wolf M, Weissing FJ (2012) Animal personalities: consequences for ecology and evolution. Trends Ecol Evol 27:452–461
Yu S, Bell D, Sternberg M, Kutiel P (2008) The effect of microhabitats on vegetation and its relationships with seedlings and soil seed bank in a Mediterranean coastal sand dune community. J Arid Environ 72:2040–2053
Ziv Y, Abramsky Z, Kotler BP, Subach A (1993) Interference competition and temporal and habitat partitioning in two gerbil species. Oikos 66:237–246
Acknowledgements
This research was supported by the US-Israel Binational Science Foundation to IS and JNP (grant no. 2013086) and the Israel Science Foundation (grant no. 442/16) to IS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. C. Choe
Electronic supplementary material
ESM 1
(DOCX 216 kb)
Rights and permissions
About this article

Cite this article
Katz, N., Pruitt, J.N. & Scharf, I. The complex effect of illumination, temperature, and thermal acclimation on habitat choice and foraging behavior of a pit-building wormlion. Behav Ecol Sociobiol 71, 137 (2017). https://doi.org/10.1007/s00265-017-2362-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-017-2362-9