
Abstract
Hybridization is a widespread phenomenon in many vertebrate groups. Prezygotic isolating mechanisms, probably caused by selection against hybrids with reduced fitness, reduce the likelihood of such events. Although hybrid-reduced fitness relatively to parental species is common, hybridization can also be beneficial, and hybrids sometimes outperform the pure species type. In this study, we examined two potential processes, Hubbs’s principle and male–male competition, which could enhance hybridization in the waterfrog complex and thus explain the proportion of heterospecific pairs collected in a natural pond. Firstly, by collecting 791 frogs in the field to study pair and chorus composition, we showed that in a mixed Rana lessonae–Rana esculenta population, the scarcity of hybrid R. esculenta males did not account for the proportion of heterospecific pairs: indeed, when examining pairing composition in six different choruses, we found that hybrid males were always under-represented and that R. esculenta females were found paired with R. lessonae males. Secondly, we investigated experimentally whether or nor male–male competition mechanism could explain pair formation in waterfrogs. Our mating speed experiment highlights mechanisms that could explain heterospecific pairs in a context of promiscuous mating where scramble competition was intense. To measure the rapidity with which a male grasps a female, we placed males in a grid cage with a female, and the dynamics of pair formation was monitored. R. esculenta males showed a lower pairing success than R. lessonae males as a smaller proportion of them amplexed females, and more time was needed for them to get amplexed. Thus, a less adaptative mechanism than female mate choice may also explain the mating pattern observed in waterfrog species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
When hybridization involves high costs (i.e. when hybrid fitness is lower than that of parental taxa), natural selection should promote reproductive isolation by favouring pre-mating mechanisms, the so-called reinforcement process (Loftus-Hills and Littlejohn 1971; Noor 1995; Saetre et al. 1997; Pfennig and Simovitch 2002). Whereas reinforcement was first considered a rare phenomenon, recent studies suggest that it may have been involved in numerous cases of species isolation (Coyne and Orr 1989; Noor 1997). Gerhardt et al. (1994) attributed the apparent rarity of reinforcement processes to the fact that nearly all studies have focused on courtship displays or signals of males rather than on the way by which females select their mates. If signals diverge significantly and are perceptually distinguishable at the time when sympatry is achieved, there is no logical necessity for further divergence in male signals (Waage 1979). Conversely, selection could mainly operate to sharpen the selectivity of females because they usually have more to loose from a mating mistake than males. When fitness values of hybrid genotypes equal or exceed those of parental species, other mechanisms could promote gene flow between species (Rosenfield and Kodric-Brown 2003).
In animals, the causes of hybridization are to be first approached at the level of mating behaviour (Grant and Grant 1997). Indeed, several behavioural mechanisms such as female mate choice or male–male competition may facilitate interbreeding (Randler 2002): (i) The “scarcity of conspecifics” hypothesis known as Hubbs’ (1955) principle states that hybridization occurs when the availability of conspecific individuals is limited by such ecological factors as resource depletion, disturbance or habitat fragmentation. Females of the rarer species initially reject males of the more common species, but the longer they search for males of their own species the less discriminating they become. As a consequence, they may mate with males of the commoner species as a result of flexibility in mate preferences (Grant and Grant 1992). In this context, hybrid offspring are produced by females of the rare species and males of the common species (Wirtz 1999). (ii) Sexual selection could also promote heterospecific mating through male–male competition. Indeed, competition among males can contribute to hybridization processes since competitive interactions are indicative of male quality. For instance, in the hybrid zone between the white-collared manakin, Manacus candei, and the golden-collared manakin, Manacus vitellinus, sexual selection through asymmetries in male aggression behaviour plays an important role in the dynamics of hybridization (McDonald et al. 2001). Moreover, in a study of hybridization between several species of the genus Cyprinodon, Rosenfield and Kodric-Brown (2003) showed that hybrid males dominate the males of both parental species under certain conditions because of their extreme aggressiveness. Competitive interactions among males may thus promote hybridization.
In the present study, we investigate pair formation in the European waterfrogs of the Rana lessonae–Rana esculenta hybrid system. The hybridization that results from such breaking of homotypic mating is one of the key conditions of the hybridogenesis model (Schultz 1969; Tunner 1974). While this model is widely accepted for explaining the composition of waterfrog assemblages (Graf and Polls Pelaz 1989), the mechanisms explaining pair formation remain poorly understood. Nevertheless, this system is an excellent model system to test hypotheses on hybridization dynamics because of the high variability of assemblage composition among breeding sites and the high density of males that could impede mate choice by females and promote male–male competition at each chorus. The edible frog R. esculenta (genotype RL) is an interspecific hybrid produced through mating between the pool frog R. lessonae (genotype LL) and the lake frog Rana ridibunda (genotype RR). Mixed R. lessonae–R. esculenta populations represent the most widespread system in the waterfrog complex (L–E system). Until recently, R. ridibunda has been absent from these systems recolonised after the last ice age. These two taxa may easily be confused because morphological identification in the field requires experience and should be confirmed by bioacoustic analysis or genetic analysis. Gamete formation in the RL hybrid obeys the hybridogenesis model (Schultz 1969; Uzzel and Berger 1975). Prior to meiosis, one of the parental genomes (L) is discarded while the other (R) is replicated and clonally transmitted to eggs or sperm. Therefore, the hybrid can only produce viable offspring through sexual parasitism on the parental species. R. esculenta thus regains the lost L genome by mating with R. lessonae, its sexual host. The hybrid coexists with the parental species, and thus, males of both taxa gather at the same breeding ponds where they form choruses that reach up to 400 males. Observations conducted at night revealed a strong competition between males. In such mixed populations, four different mating combinations are possible (Fig. 1). Homotypic matings between LL males and females lead to LL tadpoles. Homotypic RL pairings produce RR offspring, usually with null or low survival, probably because of the homozygosity of deleterious mutations accumulated owing to the lack of recombination (R genome clonally transmitted). Only when RL parents belong to different hemiclones do the tadpoles develop normally (Hotz et al. 1992; Vorburger 2001). All known R. esculenta lineages originate from matings between female R. ridibunda and male R. lessonae. Therefore, clonal R genomes contain an X chromosome, and all resulting offspring of successful matings between hybrids are females. Heterotypic LL × RL matings result in new R. esculenta individuals, but the outcomes differ according to mate gender. Mating between LL male and RL female produced three times more tadpoles (1:1 sex ratio) than the reverse combination (RL male × LL female, all-daughter progeny; Berger 1977; Joly and Moyen unpublished data). Moreover, Berger et al. (1988) showed that sex ratios of the two combinations are not the same because males and females R. esculenta produce only gametes with the female genome (see Fig. 1). Thus, in such mixed populations, there is a conflict over the best mating strategy between R. lessonae and R. esculenta. Whereas preference for the parental species is the only way to produce viable offspring in hybrid RL frogs, such a mating is expected to be avoided by parental LL frogs as the resulting RL hybrids will exclude their parental L genome in the next generation. On the other hand, if a wrong mating induces relatively low cost in males, it could imply a total fitness failure in LL females. Because hybrid individuals are often numerous in mixed assemblages, one expects that heterospecific mating is promoted by behavioural mechanisms. Pairing rules might have been adaptively shaped by natural selection, and heterospecific mate choice in RL females suggests a breaking of genetic correlation between male trait and female preference for male trait. However, such assertion is a postulate which is a by-product of the hybridogenesis model. This model indeed supposes that the mating frequency of hybrid females with parental males is higher than expected on the basis of homotypic mating or random mating. Until now, only female mate choice has been investigated to explain such unbalanced mating frequency. However, because these choice experiments have not been conducted within the usual behavioural context of sexual selection (noisy background of the chorus, possibility of scramble competition for mate capturing), the extrapolation of these results to the outcome of natural pairing remains hypothetical.

Mating combinations and descent in a mixed Rana lessonae (genotype LL)–Rana esculenta (genotype RL) assemblage (females are represented darker than males). Prior to meiosis, R. esculenta excludes the lessonae hemigenome, and thus, its gametes only contain the R genome. Since the L genome is of paternal origin, RL × LL matings lead to daughters only. Only one out of the four mating combinations gives LL tadpoles (homotypic LL pairing). The cross indicates that the other homotypic matings (RL × RL) usually produce non- or little-viable RR female offspring. Both heterotypic combinations result in new hybrid individuals
Avoiding the temptation of an adaptationist logic, our study is the first to repeat extensive observations within natural choruses and to propose and explore two alternative hypotheses to that of adaptive female mate choice. We hypothesised that (i) pairs form according to the proportion of males and females taxa in each taxon at the chorus (the proportion of heterotypic matings is confounded by the scarcity of hybrid males), and (ii) the proportion of heterotypic mating may be explained by interspecific variation in mating speed. We examined natural choruses of the R. lessonae–R. esculenta assemblage. An experimental design was used to estimate the pairing success of males.
Methods
Study site
We studied an L–E assemblage at Riquet pond located on the Dombes Plateau, 20 km north–east of Lyon, (4°55′30″E, 45°57′N) in the estate of Pierre Vérots Foundation, a private natural reserve covering a 150-ha area. This 5-ha pond was inhabited by several hundreds of waterfrogs that breed here every year. In this assemblage, the ratio is globally LL/RL = 40:60 (Colon 2004).
Field sampling
During the breeding season in May, males gather to form large choruses. We studied six choruses that were monitored from a platform situated 1 m above water surface. From this point, it was easy to spatially delimit the choruses and to count males without disturbing chorus behaviour. Male number was estimated at the beginning of the night for each monitoring session. The six choruses included 148, 85, 74, 177, 111 and 84 males, respectively. Females came singly to a chorus, often by night, got amplexed with a male, and moved towards the centre of the chorus where eggs were laid and fertilised by the male.
Sampling natural pairs made it possible to determine the mating pattern within the studied assemblage. During the breeding seasons of 2002 and 2003, we collected 112 pairs in amplexus before egg deposition in six different choruses during the first night of chorus activity. We brought the pairs in individual boxes to a laboratory located in the near vicinity (less than 5 min of travel) where we performed individual marking by means of numbered elastic waist bands (Emlen 1968). A previous pilot study established the appropriate size of elastic band in relation with frog size to avoid any adverse effect of marking. This technique is well suited for short-term marking anuran amphibians, and Emlen (1968) showed that it allowed observing normal behaviour in natural conditions (the marked males maintained normal calling station and were found in amplexus). Immediate taxon identification of individuals of each pair was performed using phenotypic traits (shape of the metatarsal tubercle; Berger 1977). We considered the females caught while in amplexus to be ready to breed. They were kept in cages placed near the shore of the pond for a few hours before the beginning of the mating speed experiment. At about midnight, we collected more than 94% of all the males present at the six studied choruses except at chorus 4 where only 89% of the males were caught, and we carried them to the laboratory. Forty males of each taxon were marked, kept in cages for a few hours before studying amplexus behaviour.
After the mating speed experiment, we collected toe samples (two phalanges) on each animal for further identification. We performed this after the animal was anaesthetized using cold water (2°C). After being warmed up, the frogs did not show any sign of distress, and the impact of removing only one toe may be small on survival rate (McCarthy and Parris 2004). The toe was immediately frozen and stored at −22°C. Then we performed an allozyme analysis, which allowed unambiguous identification of the taxon (Tunner 1973) and hence provided a check of initial phenotypic assignment (but see Schmeller 2004).
Allozyme analysis
Electrophoresis was performed on the skeletal muscles of the toe. Tissue samples were crushed in a 1.2 g Tris + 0.37 g EDTA + 1 l H2O + 5 ml NADP 1% solution. Migration took place in a Tris citrate gel at pH 6 for 3–5 h under 180 V. Tris citrate buffer composition was 48 g starch (12%), 1.4 ml buffer 1× (composition of the 10× buffer: Tris 270 g, citric acid 181 g, H2O 1,000 ml), 398.6 ml H2O. Staining solutions were prepared using modifications of standard procedures (Pasteur et al. 1987). Two loci were analysed in somatic tissues: lactate dehydrogenase (LDH-1, Enzyme Commission (EC) 1.1.1.27) and mannose-phosphate-isomerase (MPI, EC 5.3.1.8). These enzymes were chosen because they are the most discriminant markers for taxonomic identification between R. lessonae and R. esculenta.
Mating speed experiment
We compared mating swiftness between taxa. In a context of promiscuous mating where scramble competition is intense, the rapidity with which a male grasps a female is a decisive biological trait. On average, 18 h after capture (range 17–20 h), one male and one female were introduced in a grid cage (0.8×0.8×1.1 m) placed near the shore of the pond. At this location, the frogs received most chemical and acoustic cues from a chorus that was active a few metres away. Environmental conditions in the cages were also similar to those prevailing at the active chorus (water and air temperature, photoperiod). After a pair of marked frogs was introduced into each cage, we checked for amplexus every 10 min until amplexus was detected. We do not detect any difference in the response of the two taxa to handling (in terms of latency to displacement or calling behaviour). We noted the time elapsed since the beginning of the experiment, then we removed the frogs that were freed from their waist bands and released in the pond. After 3 h, the frogs still present in the cages were considered as reluctant to amplexus and were also released. Forty RL males and 33 LL males were tested to determine their ability to pair. We matched 21 RL females and 19 LL females with the RL males and 19 RL females and 14 LL females with the LL males.
Statistical analysis
Amplexus pattern in the field
Each male was characterised by three variables: its taxon (LL or RL), the chorus at which it was caught (numbered from 1 to 6) and its mating status (single or in amplexus when captured). We used a log-linear analysis (Proc GEMMOD, SAS Institute) to assess the influence of taxon on the probability of being found in amplexus. For this purpose, we first constructed the saturated model considering all the two- and three-way interactions (chorus × taxon × mating status). Likelihood ratio tests were used to test significance of the interaction terms. We dropped the non-significant interaction terms until obtaining unreductible interactions (McCullagh and Nelder 1989). In addition, for each chorus, we compared the proportions of RL males single and in amplexus (Fisher exact test and χ 2 test).
Mating speed experiment
Because proportions of LL or RL males in amplexus at the end of the experiment were not influenced by female taxon (LL males: 11 paired vs 3 unpaired with LL females and 13 paired vs 6 unpaired with RL females, Fisher exact test: P=0.698; RL males: 6 paired vs 13 unpaired with LL females and 12 paired vs 9 unpaired with RL females, χ 2=2.634, df=1, P=0.105), we pooled female taxa in subsequent analyses. The ability of males to get amplexed was then assessed by χ 2 tests. In the same way, because we found no influence of female taxon on time elapsed (Kolmogorov–Smirnov, two-sample test: P=0.999 for LL males and P=0.111 for RL males), we pooled female taxa in subsequent analyses. We then assessed if the observed difference between RL and LL males for the time delay necessary to obtain an amplexus is statistically significant (Kolmogorov–Smirnov, two-sample test).
Results
Pairing rule in mixed assemblage
Based on allozyme identification of 679 males sampled from six choruses, the LL–RL ratio in the studied pond was 64–36%. The mating pattern did not match this ratio since RL males were found in less than 15% of the observed breeding pairs, both with LL and RL females (Fig. 2). All the other pairs involved LL males that were amplexed with LL, RL and RR females (55, 44 and 1%, respectively). The female R. ridibunda probably came from R. esculenta pairings because no introduction of R. ridibunda was known in the reserve (Fig. 1). We refined this general pattern by taking into account variation among choruses. For this purpose, we performed a log-linear analysis to investigate the relationship between mating frequency and availability of each taxon within each chorus (Table 1 and Fig. 3). Chorus × taxon × mating status and chorus × mating status interactions were removed as non-significant, whereas chorus × taxon and taxon × mating status interactions proved significant. This shows, first, that the proportions of each taxon differed among choruses and, second, that the mating probability of a male was linked to its taxon. With respect to the composition of each chorus, LL males were always over-represented in the amplexed pairs, whereas RL males were under-represented. Taking into account the proportion of LL and RL males in the chorus, the probability for an LL male to mate was 3.7 (CI95% 2.15–6.38) times greater than that of an RL male.

Genetic composition of pairs at the choruses: 112 pairs were caught at six different choruses in a pond with a mixed assemblage (64%LL, 36% RL)

Proportion of RL males observed in amplexus (shaded bars) and available at the chorus (black bars): 679 LL and RL males were collected from six different choruses and 112 of them were in amplexus. Absolute numbers of frogs collected are in bracket
In spite of the low number of pairs collected on each chorus, a chorus by chorus analysis emphasised in three choruses a counter-selection of RL males compared to LL males since pair composition was bias towards LL males: chorus 1 (χ 2 test, χ 2=6.105, df=1, P=0.013), chorus 3 (Exact Fisher test, P=0.035) and chorus 4 (χ 2=4.892, df=1, P=0.027). The same pattern was observed on the other three choruses, but the difference between the two taxa was not statistically significant: chorus 2 (Exact Fisher test, P=0.733), chorus 5 (Exact Fisher test, P=0.559) and chorus 6 (χ 2=2.195, df=1, P=0.138).
Mating speed experiment
In males of both taxa, the propensity to grasp a female was not affected by female taxon. When considering the pooled data set, the proportion of amplexed males differed according to taxon since 45% of the RL males and 72.7% of the LL males mated within 3 h (\(\chi ^{2}_{1} = 5.69\), P=0.017; Fig. 4).

Capability of LL and RL males to mate under experimental conditions (each male was placed in a grid cage with one female for 3 h). a Percentage of LL and RL males observed in amplexus by the end of the experiment. b Mating delay (mean±SE) for each male taxon
Female taxon did not influence the time needed to become paired. In contrast, the time necessary to witness an amplexus during the experiment differed according to male taxon. Whereas 89.6 min was on average necessary for RL males to pair, only 40 min was necessary for LL males (Kolmogorov–Smirnov, two-sample test: P=0.013).
Discussion
Our study of mating patterns in the wild clearly shows that pairing did not follow rules of conspecific mating since most females of one of the two taxa were found to breed with heterotypic males. This result is congruent with the expectancy of the hybridogenetic model of hybridization. Our observations failed to demonstrate that Hubb’s principle (1955) could explain the hybridization process in the waterfrog L–E system: the high frequency of mating between hybrid females and parental males was indeed not due to the limited availability of hybrid males. In this way, an increase in the proportions of hybrid males did not induce a decrease in heterospecific pairs.
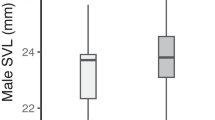
These results have to be considered under the hypotheses of two potential conflicts of interest arising in the mating process. The first conflict results from a different interest between sexes for partner characteristics related to fecundity. Males should mate preferentially with large females because in many anurans, female size is often positively correlated with clutch size (Davies and Halliday 1977; Joly 1991). In 76 pairs of waterfrogs collected in a previous study, the body of RL females was on average 14% longer than that of LL females (Lengagne, unpublished data). In anurans, such a difference leads to a fecundity variation of 66% in Rana temporaria (Joly 1991) or 77% in Bufo bufo (Davies and Halliday 1977). Consequently, there was a benefit for a male to choose a large female, and RL females may act as superstimuli for LL males. A modelling approach showed that in particular conditions, a “bigger is better” strategy can lead males to accept conspecific and heterospecific females, even though this activity bears some costs (Schmeller et al. 2005). However, choosy males are unlikely in waterfrogs since the skewed operational sex ratio (OSR) reduces the relative cost of breeding with a small female. Indeed, male selectivity depends on the probability of finding alternative mates (Johnstone et al. 1996). A surplus of available females means that a male can reject low-quality females without the risk of jeopardizing his opportunity to mate. In waterfrogs, males are always more numerous than females at each chorus, thus making strong size selectivity risky because of low mating probability. On their own, females are expected to choose potential mates according to fecundity or fertility criteria (Davies and Halliday 1977; Nakatsuru and Kramer 1982; Robertson 1990). Studies performed in anurans have identified two main male phenotypic traits involved in pair fecundity. In axillary amplecting species, fertilisation efficiency should be maximised when males and females are relatively close in body length because their cloacae will be close to each other (Licht 1976; Bourne 1993). Male mass could also be a reliable indicator of fertility. Indeed, both Robertson (1990) and Bourne (1993) emphasised mass assortative pairing and suggested that lighter males may provide insufficient sperm to fertilise the whole clutch spawned by large females. Finally, selection will act on females to avoid pairing with larger and heavier males that can impair their swimming and spawning abilities (Robertson 1986).
The second conflict of interest lies in the sexual parasitism of RL individuals on LL ones (hybridogenetic model; Joly 2001). Under the hypothesis that hybridogenesis has been favoured by natural selection, RL frogs are expected to select LL mates because homotypic RL pairings produce low viability offspring. In contrast, LL individuals should avoid mating with RL frogs because of the elimination of the L genome in the germinal line of the resulting hybrids. Hence, a potential conflict between R. lessonae and R. esculenta over the best mating strategy might arise. Homotypic matings between RL individuals were rare in our data set (9%), and we should thus expect the presence of behavioural mechanisms to avoid them. RL females were often found in amplexus with LL males (81% of total matings), a frequency close to that of homotypic LL mating (90%). A consequence of this mating pattern is that the breeding success of R. lessonae males is impeded since 38% of them were proved to mate with a hybrid female and probably lost at least a part of their reproductive investment. Hybrid RL females thus won this conflict. In contrast, LL females won the conflict with RL males because RL male × LL female mating was rare. Despite the strong bias in mating frequency, we cannot conclude that the observed pattern results from natural selection through female mate choice since other hypotheses can explain it.
Mechanisms involved in the mating pattern
In anurans, sexual selection is primarily expressed through both male–male competition and mate choice. The balance between these two mechanisms depends, at least partly, on the temporal pattern of breeding events (Wells 1977). In explosive breeders, the brief breeding activity and the OSR reduce the opportunity for female mate choice, although male–male competition is the primary determinant of male mating success (Wells 1977; Halliday 1983). In prolonged breeders, females arrive at the breeding site progressively over several weeks, and this leads to a highly skewed OSR. Males generally remain active for the duration of the whole breeding season with strong male–male competition. Females may take advantage of male clumping since this allows them to better compare a large number of potential mates within a short period of time. Waterfrogs may be considered as prolonged breeders since their reproductive period lasts from the end of April to the end of June. However, this reproduction period is divided into several bursts. A chorus, which is an area with high male density, lasts only one or a few days, and here, spawning is highly synchronised (similarity with explosive breeding). This complex situation does not allow separating the respective effects of mate choice and scramble competition.
Mate choice by females has been the object of several studies. All experiments performed in a plexiglas tank or arena showed that both females spent significantly more time with LL than with RL males, whereas males showed no preference for either LL or RL females (Abt and Reyer 1993; Roesli and Reyer 2000; Engeler and Reyer 2001). Therefore, in experimental conditions, the preference of R esculenta females for LL males converges with theoretical predictions (hybrid females should be under strong selection to avoid mating with hybrid males). Nevertheless, the conclusions raised by these studies are based on a small part of the data set used. Indeed, only 36% of the frogs tested showed a response in Engeler and Reyer (2001), and 4 out of 11 females exhibited an unambiguous preference for LL males in Abt and Reyer (1993). This raises the question of the relevance of mate choice mechanism to fully explain the strong mating pattern observed.
Male–male competition has been poorly investigated in waterfogs (Blankenhorn 1977). In anurans, large body size provides a competitive advantage that translates into greater mating success (Davies and Halliday 1978, 1979; Loman and Madsen 1986). Moreover, energetic constraints might also favour large body size so that the largest individuals remain active for longer period than smaller ones (Murphy 1994). Despite such potential competitive advantages due to larger body size, RL males were under-represented in pairs collected in the field, suggesting that male contest or chorus tenure is not involved in the mating system. Our experiment on mating speed highlights other mechanisms that could explain why RL males are often excluded from breeding in waterfrogs since male behaviour influences pairing success. In natural conditions, where male density at the chorus is often very high with sometimes up to 30 males/m2, we may assume that LL males obtain a higher breeding success from the greater swiftness with which they grasp a female. Blankenhorn (1977) first noted that LL and RL males differ by their sexual behaviour with the LL males having a higher sexual activity than RL males. Studying one chorus (90% RL–10% LL males) for 3 days, he also suggested that spatial distribution of LL and RL males in the chorus was not random. LL males were located at the edges and were assumed to be more successful because they were in the best position to grasp females coming to the chorus from the bank. It would be interesting to test the general scope of these results.
Our results clearly show a breaking of the rule of homotypic mating. From an adaptationist point of view (Joly 2001), one could consider that hybrid females choose their mates to maintain a hybrid lineage and to avoid fitness depression implied by hybrid homotypic mating. However, this attractive interpretation supposes, first that mate choice by females is the dominating mechanism of sexual selection, and then that the choice rules of hybrid females have been shaped by natural selection to escape within-taxon preference. Instead of suggesting a breaking of the genetic correlation between male trait and female preference for the trait, a more parsimonious interpretation is that male–male competition is the dominant mechanism. If the parental species differ by their mating systems (promiscuous mating based on scramble competition in R. lessonae and lek behaviour based on female mate choice in R. ridibunda), it is possible that RL females were prevented from expressing mate choice because of the permanent harassment of LL males and their better mating speed than the hybrid ones. Such a difference between taxa in male behaviour may be sufficient to explain the mating pattern observed. The males of the parental species would first benefit from their ability to overpass hybrid males in scramble competition before being then misled since heterotypic mating drives their genes to a dead-end. This last scenario supposes that a female frog does not dispose of any mechanism for identifying and controlling the male that grasps her. This question of female control is of crucial importance for the whole understanding of the hybridogenetic complex. Although if females may be prevented from expressing any choice by the frenzy with which LL males attempt amplexus in our studied choruses, the relative role of male–male composition and female choice could depend on chorus composition and general natural history of the different waterfrog populations.
Sexual behaviour and sexual selection are key mechanisms to understand the evolution of such hybridization systems, and further experiments are still needed to disentangle the divergent hypotheses about the behavioural mechanisms of mate selection. Testing these hypotheses will also bring new light on the genetic correlation between the rules of mate choice and the sexual characters involved in this choice.
References
Abt G, Reyer HU (1993) Mate choice and fitness in a hybrid frog: Rana esculenta females prefer Rana lessonae males over their own. Behav Ecol Sociobiol 32:221–228
Bergen K, Semlitsch RD, Reyer HU (1997) Hybrid female matings are directly related to the availability of Rana lessonae and Rana esculenta males in experimental populations. Copeia 1997:275–283
Berger L (1977) Systematics and hybridization in the Rana esculenta complex. In: Taylor DH, Guttman SI (eds) The reproductive biology of amphibians. Plenum, New York, pp 367–388
Berger L, Uzzell T, Hotz H (1988) Sex determination and sex ratios in western Palearctic water frogs: XX and XY female hybrids in the Pannonian basin? Proc Acad Nat Sci Philadelphia 140:220–239
Blankenhorn HJ (1977) Reproduction and mating behavior in Rana lessonae–Rana esculenta mixed populations. In: Taylor DH, Guttman SI (eds) The reproductive biology of amphibians. Plenum, New York, pp 389–409
Bourne GR (1993) Proximate costs and benefits of mate acquisition at leks of the frog Ololygon rubra. Anim Behav 45:1051–1059
Colon L (2004) L’hybridogénèse dans le complexe Rana klepton esculenta. Apports des marqueurs génétiques. Ph.D. thesis, University of Lyon 1, France
Coyne JA, Orr HA (1989) Patterns of speciation in Drosophila. Evolution 43:362–381
Davies NB, Halliday TR (1977) Optimal mate selection in the toad Bufo bufo. Nature 269:56–58
Davies NB, Halliday TR (1978) Deep croaks and fighting assessment in toads Bufo bufo. Nature 274:683–685
Davies NB, Halliday TR (1979) Competitive mate searching in male common toads, Bufo bufo. Anim Behav 27:1253–1267
Emlen ST (1968) Technique for marking anuran amphibians for behavioral studies. Herpetologica 24:172–173
Engeler B, Reyer HU (2001) Choosy females and indiscriminate males: mate choice in mixed populations of sexual and hybridogenetic water frogs (Rana lessonae, Rana esculenta). Behav Ecol 12:600–606
Gerhardt HC, Dyson M, Tanner SD, Murphy CG (1994) Female treefrogs do not avoid heterospecific calls as they approach conspecific calls: implications for mechanisms of mate choice. Anim Behav 47:1323–1332
Graf J-D, Polls Pelaz M (1989) Evolutionary genetics of the Rana esculenta complex. In: Dawley RM, Bogart JP (eds) Evolution and ecology of unisexual vertebrates. New York State Museum Bulletin, Albany, pp 298–302
Grant PR, Grant BR (1992) Hybridization of bird species. Science 256:193–197
Grant PR, Grant BR (1997) Hybridization, sexual imprinting, and mate choice. Am Nat 149:1–28
Halliday T (1983) Do frogs and toads choose their mates? Nature 306:226–227
Hotz H, Beerli P, Spolsky C (1992) Mitochondrial DNA reveals formation of nonhybrid frogs by natural matings between hemiclonal hybrids. Mol Biol Evol 9:610–620
Hubbs CL (1955) Hybridization between fish species in nature. Syst Zool 4:1–20
Johnstone RA, Reynolds JD, Deutsch JC (1996) Mutual mate choice and sex differences in choosiness. Evolution 50:1382–1391
Joly P (1991) Variation in size and fecundity between neighbouring populations in the common frog, Rana temporaria. Alytes 9:79–88
Joly P (2001) The future of the selfish hemiclone: a neodarwinian approach to waterfrog evolution. Mitt Mus Nat Kd Berl 77:31–38
Licht LE (1976) Sexual selection in toads (Bufo americanus). Can J Zool 54:1277–1284
Loftus-Hills JJ, Littlejohn MJ (1971) Pulse repetition rate as the basis for mating call discrimination by two sympatric species of Hyla. Copeia 1971:154–156
Loman J, Madsen T (1986) Reproductive tactics of large and small male toads Bufo bufo. Oikos 46:57–61
McCarthy MA, Parris KM (2004) Clarifying the effect of toe clipping on frogs with Bayesian statistics. J Appl Ecol 41:780–786
McCullagh M, Nelder PJA (1989) Generalized linear models. Chapman & Hall, London
McDonald DB, Clay RP, Brumfield RT, Braun MJ (2001) Sexual selection on plumage and behavior in an avian hybrid zone: experimental tests of male–male interactions. Evolution 55:1443–1451
Murphy CG (1994) Chorus tenure of male barking treefrogs, Hyla gratiosa. Anim Behav 48:763–777
Nakatsuru K, Kramer DL (1982) Is sperm cheap? Limited male fertility and female choice in the lemon Tetra (Pisces, Characidae). Science 216:753–755
Noor MAF (1995) Speciation driven by natural selection in Drosophila. Nature 375:674–675
Noor MAF (1997) How often does sympatry affect sexual isolation in Drosophila? Am Nat 149:1156–1163
Pasteur N, Pasteur G, Bonhomme F, Catalan J, Britton-Davidian J (1987) Manuel technique de génétique par éléctrophorèse de protéines. Lavoisier, Paris
Pfennig KS, Simovitch MA (2002) Differential selection to avoid hybridization in two toad species. Evolution 56:1840–1848
Randler C (2002) Avian hybridization, mixed pairing and female choice. Anim Behav 63:103–119
Robertson JGM (1986) Female choice, male strategies and the role of vocalizations in the australian frog Uperoleia rugosa. Anim Behav 34:773–784
Robertson JGM (1990) Female choice increases fertilization success in the Australian frog Uperoleia laevigata. Anim Behav 39:639–645
Roesli M, Reyer HU (2000) Male vocalization and female choice in the hybridogenetic Rana lessonae/Rana esculenta complex. Anim Behav 60:745–755
Rosenfield JA, Kodric-Brown A (2003) Sexual selection promotes hybridization between Pecos pupfish, Cyprinodon pecosensis and sheepshead minnow, C. variegatus. J Evol Biol 16:595–606
Saetre GP, Kral M, Bures S (1997) Differential species recognition abilities of males and females in a flycatcher hybrid zone. J Avian Biol 28:259–263
Schmeller DS (2004) Tying ecology and genetics of hemiclonally reproducing waterfrogs (Rana, Anura). Ann Zool Fenn 41:681–687
Schmeller DS, O’Hara R, Kokko H (2005) Male adaptive stupidity: male mating pattern in hybridogenetic frogs. Evol Ecol Res 7:1039–1050
Schultz RJ (1969) Unisexuallity and polyploidy in the teleost Poeciliopsis (Poecilidae) and other vertebrates. Am Nat 103:605–619
Tunner HG (1973) Das albumin und andere bluteiweisse bei Rana ridibunda Pallas, Rana lessonae Camerano, Rana esculenta Linné und deren hybriden. Z Zoolog Syst Evol Forsch 11:219–233
Tunner HG (1974) Die klonale struktur einer wasserfroschpopulation. Z Zoolog Syst Evol Forsch 12:309–314
Uzzell T, Berger L (1975) Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenetic associate, Rana esculenta. Proc Acad Nat Sci Philadelphia 127:13–24
Vorburger C (2001) Non-hybrid offspring from matings between hemiclonal hybrid waterfrogs suggest occasional recombination between clonal genomes. Ecol Lett 4:628–636
Waage JK (1979) Reproductive character displacement in Calopteryx (Odonata: Calopterygidae). Evolution 33:104–116
Wells KD (1977) The social behaviour of anuran amphibians. Anim Behav 25:666–693
Wirtz P (1999) Mother species–father species: unidirectional hybridization in animals with female choice. Anim Behav 58:1–12
Acknowledgements
The study was supported by the Centre National de la Recherche Scientifique (C.N.R.S.). We are indebted to the Foundation Vérots for allowing us to carry out this study in their reserve. We are grateful to Albane Colin, Julien Mousset and Aurélien Jamoneau for their help in the field. The study benefited from valuable advice by Tim Halliday, Jakob Christensen-Dalsgaard, Sandrine Plénet, Jean-Paul Léna, Bernard Kaufmann and three anonymous referees. We thank Jean-Paul Léna for his help in statistical analysis, and we are grateful to Eric Pattee, Bernard Kaufmann and Tim Halliday for the improvement of the English. This study was conducted with the approval of Préfecture de l’Ain in accordance with the current laws in France.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. Christensen-Dalsgaard
Rights and permissions
About this article
Cite this article
Lengagne, T., Grolet, O. & Joly, P. Male mating speed promote hybridization in the Rana lessonae–Rana esculenta waterfrog system. Behav Ecol Sociobiol 60, 123–130 (2006). https://doi.org/10.1007/s00265-005-0148-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-005-0148-y