
Abstract
To detect threats and reduce predation risk prey animals need to be alert. Early predator detection and rapid anti-predatory action increase the likelihood of survival. We investigated how foraging affects predator detection and time to take-off in blue tits (Parus caeruleus) by subjecting them to a simulated raptor attack. To investigate the impact of body posture we compared birds feeding head-down with birds feeding head-up, but could not find any effect of posture on either time to detection or time to take-off. To investigate the impact of orientation we compared birds having their side towards the attacking predator with birds having their back towards it. Predator detection, but not time to take-off, was delayed when the back was oriented towards the predator. We also investigated the impact of foraging task by comparing birds that were either not foraging, foraging on chopped mealworms, or foraging on whole ones. Foraging on chopped mealworms did not delay detection compared to nonforaging showing that foraging does not always restrict vigilance. However, detection was delayed more than 150% when the birds were foraging on whole, live mealworms, which apparently demanded much attention and handling skill. Time to take-off was affected by foraging task in the same way as detection was. We show that when studying foraging and vigilance one must include the difficulty of the foraging task and prey orientation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animals need to be alert and attentive to detect potential threats and reduce predation risk. If a prey detects a predator at an early stage of an attack the hunt is often abandoned (e.g. FitzGibbon 1989; Lingle and Wilson 2001). Additionally, the sooner a prey initiates its flight the greater the probability of making a successful escape (Kenward 1978; FitzGibbon 1989; Krause and Godin 1995). Prey can also become aware of impending danger not detected by themselves by viewing and responding to other group members' escape initiation (e.g. Lima 1994; Hilton et al. 1999).
So being observant is beneficial to avoid becoming a predator meal, but factors such as body posture and activity can reduce preys' attention ability. When guppies (Poecilia reticulata) forage nose-down on the bottom substrate their visual view appears restricted since their escape is delayed compared to when they forage with their body positioned horizontally (Krause and Godin 1996). When animals are engaged in complex activities their ability to focus on potential threats seems reduced since their anti-predatory actions are delayed. This has been observed during escalated fighting (Jakobsson et al. 1995; Brick 1998), social playing (Harcourt 1991; Blumstein 1998), allogrooming (Maestripieri 1993; Cords 1995) and foraging (FitzGibbon 1989, 1990; Krause and Godin 1996; Lima and Bednekoff 1999; Hilton et al. 1999).
Attention ability in blue jays (Cyanocitta cristata) has been investigated under simulated foraging scenarios (Dukas and Kamil 2000). The birds were trained to distinguish and peck on targets on a computer monitor. When a task was difficult (many distracting objects on the monitor) the birds lost some attention for peripheral targets and the authors suggest that, in nature, animals engaged in difficult food tasks incur a higher risk of predation due to impaired predator detection. Complex foraging tasks are also known to reduce scanning behaviour, which probably results in reduced attention to the surroundings (in birds: e.g. Lendrem 1983; Lawrence 1985; Popp 1988).
Blue tits (Parus caeruleus), a small passerine bird, have a varied diet and feed on, for example, insects, spiders, fruits and seeds (Cramp and Perrins 1994). The purpose of our study was to investigate how predator detection and time to take-off in blue tits are influenced by different foraging situations. The birds were subjected to a simulated raptor attack. The impact of body posture was studied by comparing birds feeding in head-up or head-down postures. Birds often feed with their head down and frequently raise their head to scan, which possibly implies that these different postures might influence their view and detection ability. The impact of task difficulty was studied by comparing birds that were either not foraging, foraging on chopped mealworms (Tenebrio molitor) or foraging on whole, live mealworms. How orientation affects predator detection was studied by comparing birds oriented with their side or back towards the predator.
Methods
Birds and experimental set-up
This study was carried out at Tovetorp Zoological Research Station in south-eastern Sweden. Experiments were conducted during two succeeding winters (2001 and 2002) under laboratory conditions. Blue tits were caught using trapping cages and mist nets. Following capture, birds were housed indoors in individual cages (80×40x60 cm) equipped with two perches. Water and food (sunflower seed and suet) were available ad libitum. To give prior experience to the different foraging tasks used in the experiment (see below) each bird was also provided 5–6 mealworms, presented as in the experiments, 3 times/day for a minimum of 2 days.
We used the following experimental set-up. When subjected to an attack, a blue tit sat on a perch 55 cm above ground on a pole in a compartment. An attack was simulated by having a stuffed female sparrowhawk (Accipiter nisus) gliding slowly down outside the compartment towards the blue tit. All sides of the blue tit compartment (2×0.7×2 m high) consisted of nontransparent walls except the one towards the raptor, which was made of a glass screen and a mesh net. On attack, the sparrowhawk appeared through a hole and glided along a wire towards the blue tit perch at an angle of 15°. A thread which we held on to was attached to the sparrowhawk and by releasing the thread the sparrowhawk started gliding and the attack was launched. The sparrowhawk glided 2.2 m in 2.7 s before stopping in front of the perch. Since the distance the hawk travelled was quite short, we chose a slow attack speed in order to increase attack duration and allow us to obtain a large range of detection times. The hawk's gliding produced a faint sound. To exclude audible clues about the attack we recorded fragments of this sound and played it incessantly at similar sound power inside the compartment.
One bird at the time spent 20 min in a cloth bag and was thereafter released into the experimental room and subjected to an attack. A bird was only attacked once whereupon it was ringed and released. Each bird was assigned to one of the following treatments. When being attacked, the blue tit was sitting head-up (upright) and was either not foraging, foraging on chopped mealworms or foraging on whole, live mealworms, or the bird was sitting with its head down foraging on whole, live mealworms. In all these groups we had birds that were oriented with their back and birds that were oriented with their side towards the sparrowhawk. Order of treatments was randomised so as not to introduce any directional bias in case time of day would influence the results.
Impact of body posture
To investigate the impact of body posture on detection and take-off times we compared birds feeding head-down on whole mealworms with birds feeding head-up on whole mealworms when attacked. Five whole, live mealworms were threaded dorsoventrally on a steel wire and presented at the perch at either the height of the birds' head or feet. Mealworms presented in these ways appeared to be a challenging task and therefore birds remained in their postures during launch of the attack and predator detection, and hence any difference in detection ability between groups would probably be due to head position itself since all else was equal.
Impact of foraging task
To investigate the impact of foraging task on detection and take-off times we compared these three treatments: nonforaging, foraging on chopped mealworms and foraging on whole, live mealworms. Mealworms were presented at head height since nonforaging birds were sitting head-up. Consequently, all birds had the same body posture and any difference in detection time would therefore be due to task alone. Nonforaging represented the easiest, chopped mealworms intermediate and whole, live mealworms the most difficult situation. In the chopped treatment five mealworms were each cut into five to nine pieces, depending on size, and presented on a small transparent tray (approximately 3×1.5 cm). The tray had only two edges, none on the side towards the blue tit, so that the birds would not have to bend over it, and none on the side towards the predator when attacked from the side, so that no border should prevent detection. Data for birds in the difficult foraging task treatment (whole mealworms) are the same ones as in the head-up posture treatment.
Impact of orientation
To investigate how the birds' orientation affects detection time and time to take-off we compared birds that were oriented with their back towards the raptor with birds that were side-on to it.
Data recording and analysis
We recorded the blue tits with a Sony Digital Camera DCR-VX1000E. During the experiments the video camera was connected to a screen enabling us to see the birds from outside and to launch the attack when the bird was in the desired position. The video camera had a frame resolution of 0.04 s. We used frame-by-frame analysis to determine the birds' detection time and time to taking off, that is how long from launch of attack until the birds detected the incoming sparrowhawk and how long from launch of attack until they raised their wings and took flight. Detection was defined as the first frame in which a bird froze with its head before taking flight. Mostly the blue tits froze with their head for a couple of frames before taking flight. On a few occasions they took flight after only a quick head movement directed towards the predator and in these cases the frame showing this glance was taken as detection.
Ninety-three birds were used, all males, 58 were young (born the previous spring) whereas 35 were older. The number of young and old birds was distributed between treatments as evenly as possible. The birds were housed indoors between 2 and 8 days before the experiment. All statistical analysis were done using Statistica 5.5 A (Statsoft) and data are presented as mean ± SE.
Results
All blue tits detected the sparrowhawk and responded by taking off into the air. The blue tits behaved differently when engaged in the two mealworm tasks. When foraging on chopped mealworms feeding bouts were rather short. The birds repeatedly and rapidly picked up pieces of mealworms between which they scanned or handled the food in their beak and detected the sparrowhawk. In contrast, when foraging on whole, live mealworms the birds had longer feeding bouts during which they detected the sparrowhawk. The whole mealworms twisted and twirled a lot and the blue tits usually caught a live mealworm by aiming at the head. Once a mealworm was caught they tried to pull it off the wire.
Body posture
We could not detect any significant difference in detection time between birds feeding in head-down and head-up posture on whole mealworms, whether their side (one-way ANOVA: F 1,22=0.68, P=0.42) or their back was oriented towards the predator (one-way ANOVA: F 1,19=0.20, P=0.66) (Table 1). Thus they seem to be able to see equally well with lowered as with raised head so these postures do not appear to significantly influence predator detection in blue tits. Neither could we detect any significant difference in time to take-off between these body postures, whether their side (one-way ANOVA: F 1,22=0.01, P=0.91) or back was oriented towards the predator (one-way ANOVA: F 1,19=0.55, P=0.47) (Table 1). In the following analyses, head-down and head-up detection and take-off times are pooled within each orientation treatment (being oriented towards predator with back or side). Consequently there are only two groups of birds foraging on whole mealworms, one with attack from behind and one with attack from side.
Foraging task and orientation
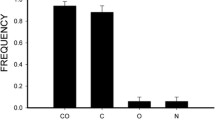
Detection time was affected by both task and orientation, but not by their interaction (two-way ANOVA: task: F 2,87=44.0, P<0.001; orientation: F 1,87=5.93, P=0.017; interaction: F 2,87=1.29, P=0.28) (Table 1, Fig. 1). We could not detect any difference in detection between birds that were not foraging and birds foraging on chopped mealworms (Tukey HSD test: P= 0.998), but birds feeding on whole mealworms detected the predator later than both nonforaging birds (Tukey HSD test: P<0.001) and birds feeding on chopped mealworms (Tukey HSD test: P<0.001) (Table 1, Fig. 1). When birds were oriented with their side towards the predator they detected the sparrowhawk earlier compared to when facing it with their back (Table 1, Fig. 1).

Predator detection times by blue tits (Parus caeruleus) for the different treatments; Nonfor nonforaging, Chop foraging on chopped mealworms, Wh up foraging on whole mealworms head-up and Wh down foraging on whole mealworms head-down. The birds had either their side or their back oriented towards the incoming predator. Ind number of individuals
Time to take-off was affected by task but not by orientation nor by the interaction of task and orientation (two-way ANOVA: task: F 2,87=19.3, P<0.001; orientation: F 1,87=1.54, P=0.22; interaction: F 2,87=0.86, P=0.42) (Table 1). The birds took off at the same time whether they were not feeding or feeding on chopped mealworms (Tukey HSD test: P=0.82), but when feeding on whole mealworms take-off was delayed both compared to when not foraging (Tukey HSD test: P<0.001) and compared to feeding on chopped mealworms (Tukey HSD test: P<0.001) (Table 1).
Discussion
We could not find any effect of body posture on detection ability and time to take-off. The blue tits detected the predator equally well and took off equally soon when they were feeding head-down as head-up. And a very easy foraging task does not seem to restrict detection ability at all whereas a more complicated task restricts detection severely. An assumption underlying many models of anti-predator vigilance is that animals with a head-down posture while feeding are unable to detect incoming predators, but as recently pointed out and shown by Lima and Bednekoff (1999) this assumption is not universally applicable (see their paper for detailed discussion and references). They flew a mounted hawk towards dark-eyed juncos (Junco hyemalis) and showed that birds feeding with their heads down have a rather high ability to detect predators even though it was slightly reduced. Lima and Bednekoff (1999) therefore suggested that there is low-quality and high-quality vigilance. Low-quality vigilance occurs when birds are head down actively feeding and high-quality vigilance occurs when birds raise their head in an overt scan. Since we discriminated between the effects of body posture and foraging task on vigilance, we further suggest that it is the foraging task per se, and not posture, that delays detection in foraging birds, at least in blue tits. Therefore it seems as if the foraging task is one of the main determinants of the quality of vigilance and an individual's detection ability. Nevertheless, by interrupting feeding to scan birds can be more vigilant and increase their ability to detect predators when the task is difficult and there may be advantages associated with raising the head when scanning. The visual field can be obstructed by the environment when head-down (Metcalfe 1984) and then raising of the head becomes necessary. From this reasoning, scanning also becomes necessary when the foraging task is easy even though an easy task does not reduce vigilance. In contrast to our study, feeding guppies in a vertical nose-down body posture flee later from a predator than feeding guppies in a horizontal posture, so their visual view might be affected by these postures (Krause and Godin 1996). This disparity between blue tits and guppies is probably due to the many morphological differences between these taxa. Krause and Godin (1996) also suggest that it might be difficult for fish to perform a fast-start escape in a nose-down posture.
The degree of difficulty of the foraging task determined the impact on detection and take-off. When the blue tits were feeding on chopped mealworms they detected the predator and escaped as early as when they were not feeding showing that foraging does not necessarily restrict vigilance (Table 1, Fig. 1). Feeding on whole mealworms dramatically delayed detection with between 151 and 168% (0.5–0.7 s), and take-off was delayed with between 31 and 53% (0.3–0.5 s). Whole, live mealworms are probably a much more difficult and challenging foraging task than chopped mealworms. Reduced vigilance when engaged in a difficult task might be due to limitation in the nervous system in the amount of information that can be processed simultaneously. Some sensory information received is left unprocessed when engaged in something difficult which results in lost attention to other things (Desimone 1998; for discussion see Dukas and Kamil 2001). Our results support the suggestion from Dukas and Kamil (2000) and we provide evidence that foragers engaged in difficult food tasks incur a higher risk of predation due to impaired predator detection. Noticeably, even when the blue tits were side-on to the sparrowhawk and the hawk was in their visual field detection and take-off were delayed when the task was difficult. In line with our results, fish feeding on high densities of prey with a large confusion effect more often overlook a predator compared to when feeding on prey at lower densities (Milinski 1984; Godin and Smith 1988).
What quantitative effect might a difficult foraging task have in nature? Raptors are common predators on small birds and they often hunt by surprise (Rudebeck 1950; Cresswell 1993, 1996). Accipiter hawks have been recorded to attack at 5–28 m/s (Goslow 1971; Hilton et al. 1999). If a blue tit is attacked at 28 m/s a 0.5-s delay in taking flight (Table 1) would result in the predator advancing an additional 14 m which would severely influence the blue tit's escape probability. A 0.3-s delay in taking off would result in an extra 8 m of advancement. At the other extreme, if a predator attacks more slowly at 5 m/s a 0.5-s delay results in 2.5 m and a 0.3-s delay in 1.5 m advancement. Small differences are known to be of great importance in other predator-prey systems. Thomson's gazelles (Gazella thomsoni) which manage to flee from an attacking cheetah (Acinonyx jubatus) flee on average 1.2 s earlier than gazelles which are captured (FitzGibbon 1989). A 2 cm extra approach by a cichlid predator before a guppy flees results in twice as high probability of capture (Krause and Godin 1996).
How the blue tits were oriented towards the attacking predator affected their detection ability. When attacked from behind predator detection was delayed with between 26% and 36% (0.1–0.3 s) compared to when attacked from the side (Table 1, Fig. 1). Blue tits most likely have a large visual field since their eyes are positioned at the sides of their heads. However, their view is probably restricted backwards which might explain delayed detections when attacked from behind. Though side-oriented birds detected the predator earlier than back-oriented birds there was not any significant difference in time to take-off between them. This means that birds with their back towards the predator took a shorter time from detecting the predator to actually fleeing than birds with their side towards it. This might be due to the short attack distance. When having their back towards the predator detection becomes delayed and since the predator is therefore very close perhaps they cannot afford to take as long as side-oriented birds before taking flight.
Vigilance is a vast field which has not yet been thoroughly explored (Lima and Bednekoff 1999). Our study has yielded further knowledge and we suggest that head-down posture itself need not significantly decrease detection ability in foraging blue tits, whereas body orientation relative to the direction of the predator's attack does. We also show that the degree of difficulty of the foraging task determines the impact of foraging on predator detection ability. An easy foraging task enables prey to concurrently have full attention directed towards discovering predators whereas a difficult task severely delays both detection and time to take-off, which probably increases the risk of being caught by a predator. Future studies of vigilance and predation risk need to include prey orientation and foraging task.
References
Blumstein DT (1998) Quantifying predation risk for refuging animals: a case study with golden marmots. Ethology 104:501--516
Brick O (1998) Fighting behaviour, vigilance and predation risk in the cichlid fish Nannacara anomala. Anim Behav 56:309--317
Cords M (1995) Predator vigilance costs of allogrooming in wild blue monkeys. Behaviour 132:559--569
Cramp S, Perrins CM (1994) The birds of the western Palearctic, vol 9, Oxford University Press, Oxford
Cresswell W (1993) Escape responses by redshanks, Tringa totanus, on attack by avian predators. Anim Behav 46:609--611
Cresswell W (1996) Surprise as a winter hunting strategy in sparrowhawks Accipiter nisus, peregrines Falco peregrinus and merlins F. columbarius. Ibis 138:684--692
Desimone R (1998) Visual attention mediated by biased competition in extrastriate visual cortex. Philos Trans R Soc Lond B 353:1245--1255
Dukas R, Kamil AC (2000) The cost of limited attention in blue jays. Behav Ecol 11:502--506
Dukas R, Kamil AC (2001) Limited attention: the constraint underlying search image. Behav Ecol 12:192--199
Fitzgibbon CD (1989) A cost to individuals with reduced vigilance in groups of Thomson's gazelles hunted by cheetahs. Anim Behav 37:508--510
Fitzgibbon CD (1990) Why do hunting cheetahs prefer male gazelles? Anim Behav 40:837--845
Godin JG J, Smith SA (1988) A fitness cost of foraging in the guppy. Nature 333:69--71
Goslow Jr. GE (1971) The attack and strike of some North American raptors. Auk 88:815--827
Harcourt R (1991) Survivorship costs of play in the south-american fur-seal. Anim Behav 42:509--511.
Hilton GM, Cresswell W, Ruxton GD (1999) Intraflock variation in the speed of escape-flight response on attack by an avian predator. Behav Ecol 10:391--395
Jakobsson S, Brick O, Kullberg C (1995) Escalated fighting behaviour incurs increased predation risk. Anim Behav 49:235--239
Kenward RE (1978) Hawks and doves: factors affecting success and selection in goshawk attacks on woodpigeons. J Anim Ecol 47:449--460
Krause J, Godin JGJ (1995) Predator preferences for attacking particular prey group sizes: consequences for predator hunting success and prey predation risk. Anim Behav 50:465--473
Krause J, Godin JGJ (1996) Influence of prey foraging posture on flight behavior and predation risk: predators take advantage of unwary prey. Behav Ecol 7:264--271
Lawrence SE (1985) Vigilance during easy and difficult foraging tasks. Anim Behav 33:1373--75
Lendrem DW (1983) Predation risk and vigilance in the blue tit (Parus careuleus). Behav Ecol Sociobiol 14:9--13
Lima SL (1994) On the personal benefits of antipredatory vigilance. Anim Behav 48:734--736
Lima SL, Bednekoff PA (1999) Back to the basics of antipredatory vigilance: can nonvigilant animals detect attack? Anim Behav 58:537--543
Lingle S, Wilson WF (2001) Detection and avoidance of predators in white-tailed deer (Odocoileus virginianus) and mule deer (O. hemionus). Ethology 107:125--147
Maestripieri D (1993) Vigilance costs of allogrooming in macaque mothers. Am Nat 141:744--753
Metcalfe NB (1984) The effects of habitat on the vigilance of shorebirds: is visibility important? Anim Behav 32:981--985
Milinski M (1984) A predator's costs of overcoming the confusion-effect of swarming prey. Anim Behav 32:1157--1162
Popp JW (1988) Effects of food-handling time on scanning rates among American goldfinches. Auk 105:384--385
Rudebeck G (1950) The choice of prey and modes of hunting of predatory birds with special reference to their selective effect. Oikos 2:65--88
Acknowledgements
We thank Patrik Lindenfors for statistical advice and Christer Wiklund, two anonymous referees, Peter Bednekoff and Sven Jakobsson for valuable comments improving the manuscript. We also thank the Schöns, the Berglunds and the Bergmans for letting us trap birds in their gardens. This study was supported by grants from Elis Wide's (Gustaf Danielsson's) Foundation (Swedish Ornithological Society) to U.K. and conducted with permission No. 66–98 from the Swedish ethical board for conducting behavioural research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by P.A. Bednekoff
Rights and permissions
About this article
Cite this article
Kaby, U., Lind, J. What limits predator detection in blue tits (Parus caeruleus): posture, task or orientation?. Behav Ecol Sociobiol 54, 534–538 (2003). https://doi.org/10.1007/s00265-003-0665-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-003-0665-5