
Abstract
In order to develop a clean and effective approach for producing the valuable drug diosgenin from Dioscorea zingiberensis tubers, two successive processes, enzymatic saccharification and microbial transformation, were used. With enzymatic saccharification, 98.0% of starch was excluded from the raw herb, releasing saponins from the network structure of starch. Subsequently, the treated tubers were fermented with Trichoderma reesei under optimal conditions for 156 h. During microbial transformation, glycosidic bonds, which link β-d-glucose or α-l-rhamnose with aglycone at the C-3 position in saponins, were broken down effectively to give a diosgenin yield of 90.6 ± 2.45%, 42.4% higher than that obtained from bioconversion of raw tubers directly. Scaled up fermentation was conducted in a 5.0-l bioreactor and gave a diosgenin yield of 91.2 ± 3.21%. This is the first report on the preparation of diosgenin from herbs through microbial transformation as well as utilizing other available components in the raw material, providing an environmentally friendly alternative to diosgenin production.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Diosgenin, a precursor for partial synthesis of oral contraceptives, sex hormones, and other steroids, is widely used in pharmaceutical industry (Saunders et al. 1986; Oncina et al. 2000; Wang et al. 2007a). Nowadays, many researchers found that diosgenin has antiproliferative and proapoptotic effects on cancer cells (Trouillas et al. 2005; Bertranda et al. 2009; Chiang et al. 2007; Jayadev and Ranjana 2007) or on rheumatoid arthritis synoviocytes (Liagre et al. 2004); it also shows pharmacological activities such as antilipoperoxidative (Jayachandran et al. 2009) and antiskin aging (Yayoi et al. 2009) effects. Preparation of diosgenin from plant tubers has been extensively reported (Zhang et al. 2006; Adham et al. 2009). The main raw material used in industry in China is Dioscorea zingiberensis C. H. Wright (DZW) because of the high content of diosgenin in its tubers (Wang et al. 2008; Huang et al. 2008).
Diosgenin occurs in plants in the form of saponins attaching glucose or rhamnose to aglycone by glycosidic bonds at C-3 and C-26 (Qian et al. 2006). Preparation of diosgenin from saponins mainly depends on hydrolyzation of sugars at these two positions. In industry, sulfuric acid is usually applied in hydrolyzing raw herb to produce diosgenin. This method, however, is associated with many environmental problems due to the high concentration of chemical oxygen demand (COD; 50,000–80,000 mg l−1), SO4 2− (29,700–46,200 mg l−1), and acid (pH 0.18–0.46) in wastewater resulting from the acid hydrolysis process (Zhao et al. 2008; Cheng et al. 2009).
Efforts have been made by many researchers to solve this problem by focusing on clean methods to produce diosgenin. One interesting aspect is recovery of cellulose and starch from raw material using physical or biological approaches before acid hydrolysis, by which 50–70% of COD is reduced in the processing wastewater. Nevertheless, residual COD (10,000–20,000 mg l−1), SO 2−4 , and H+ from acid hydrolysis still lead to water pollution (Wang et al. 2008). Releasing diosgenin through biological transformation (microorganisms and enzymes) is another potential technique (Fernandes et al. 2003). To convert saponins into diosgenin, multi-enzymes (cellulase, β-glucosidase, and pectinase) have been used (Huang et al. 2008). However, this method is not economical because of the high price of commercial enzymes (Wang et al. 2007b). If microorganisms could be applied in transforming saponins, the cost of the biological process would be significantly reduced. Some fungal strains identified as members of the genera Penicillin, Aspergillus, and Curvularia have the ability to cleave sugar chains in saponins (Feng et al. 2005, 2007; He et al. 2006; Zhao et al. 2007; Qi et al. 2009). But the low diosgenin yield is a major drawback for their application (Zhao et al. 2007; Qi et al. 2009). So far, most of the microbial research has focused on identifying steroidal saponins in the bioprocess to reveal the mechanism of the transformation, but few studies have been carried out concerning means of increasing diosgenin yield. Therefore, it is of great importance to develop an effective microbial approach for diosgenin production.
The three main component groups of DZW tubers are starch (30–40%, w/w), cellulose (40–50%), and saponins (2–4%; Wang et al. 2008; Huang et al. 2008). Saponins in plant cells are in the network structure of starch (Cao 2004), which reduces the contact of saponins and microorganisms. This might be one of the reasons for the low diosgenin productivity of microbial transformation in previous studies (Qi et al. 2009). We set out to resolve this problem by excluding starch with enzymatic saccharification from raw material before microbial transformation.
For the reasons above, a combined technology of enzymatic saccharification with microbial transformation to prepare diosgenin from DZW was developed in this study. Based on a primary screening of strains from available fungal cultures and soil isolates, Trichoderma reesei which bioconverts saponins effectively was selected. The purpose of this study was to investigate the feasibility of this new approach for diosgenin production in the laboratory.
Materials and methods
Material
Dried DZW tubers were provided by Tianhe Pharmaceutical Co. (Yunxi, Hubei, China). α-Amylase (2,000 IU g−1) and saccharifying enzyme (10,000 IU g−1) were obtained from Beijing Donghua Biotechnology Co. (Beijing, China).
Microorganism
The fungal strain T. reesei (ACCC 30597) was purchased from the Agricultural Culture Collection of China (Beijing, China) and acclimated in a sterilized substrate of 50% multisaponins. The strain was stored at 4°C on potato dextrose agar slant and subcultured routinely every 2 weeks.
To prepare the inocula, spores in a 7-day-old agar slant were suspended in 5 ml of 0.01% Tween 80 solution (107 spores ml−1) and transferred into 100 ml of subculture medium composed of (in grams per liter) 30 sucrose, 3 NaNO3, 1 K2HPO4, 0.5 MgSO4, 0.01 FeSO4, and 0.1 Tween 80. Fungal cells were subgenerated on a temperature controlled shaking incubator at 150 rpm at 30°C for 72 h.
Enzymatic saccharification
Dried DZW tuber (500 g) was cleaned, cut, and ground in a pulverator (model 6020; Beijing Huanyatianyuan Co., Ltd., Beijing, China), uniformly mixed with 3.0 l tap water in a 5.0-l fermenter (SY3005; Shanghai Shiyuan Bioequipment Co., Ltd., Shanghai, China), and boiled for 1 h, then adjusted to pH 6.5 after cooling, incubated at 70°C for 1 h with 10 g of dry amylase. The enzymatic hydrolyzate was adjusted to pH 4.0 at room temperature and then saccharified with 15 g of saccharifying enzyme at 60°C for 8 h, followed by centrifugation at 4,000 rpm for 10 min. The solid residue was washed with water, centrifuged, and dried at 60°C, ground again to pass through a 100-mesh screen to obtain pretreated DZW (PDZW). The contents of ash, moisture, starch, fiber, reducing sugar, and saponins in DZW and PDZW were analyzed according to the methods described by Huang et al. (2008). The morphological features of DZW and PDZW were observed with a FEI-Quanta 200F scanning electron microscope (SEM; FEI Company, USA).
Microbial transformation
Shake flask cultivation
Microbial transformation tests were conducted in 250 ml Erlenmeyer flasks containing 100 ml medium composed of [in percent (weight/volume)] 2.67 peptone, 0.29 K2HPO4, 0.73 Tween 80, 9.77 PDZW (27.1 DZW), and pH 5.8. Each flask was autoclaved, inoculated with 10 ml of subcultured fungal cells, and incubated aerobically in a temperature-controlled shaker at 150 rpm and 30°C for 156 h.Footnote 1 One set of flasks was taken out from the shaker every 12 h. Biomass, reducing sugar, β-glucosidase (EC 3.2.1.21), α-glucosidase (EC 3.2.1.21), diosgenin yield, and saponins in the microbial transformation process were determined to evaluate the transformation efficiency. All experiments were carried out in triplicate and values were averaged. One control flask was prepared, in which no strain was inoculated.
Batch fermentation
The microbial transformation was scaled up in a 5.0-l stirred tank bioreactor (Biotech-5BG automatic fermentor; Shanghai Baoxing Biological Equipment Co., Ltd., Shanghai, China) with a 3.0-l working volume of optimal medium composed of [in percent (weight/volume)] 2.67 peptone, 0.29 K2HPO4, 0.73 Tween 80, 9.77 PDZW (27.1 DZW), and 0.01 silicon antifoam agent. The bioreactor was sterilized at 121°C for 15 min and the initial medium pH was adjusted to 5.8 with 1 M HCl. After inoculation with 300 ml subcultured strain, the fermentation lasted 156 h at 30°C with an agitation rate of 300 rpm and aeration speed of 0.8 vvm, followed by centrifugation at 4,000 rpm for 10 min. The solid was dried at 60°C and ground for extraction of diosgenin.
Microbial transformation pathway for saponins
The main saponins in PDZW and converted products were separated by thin-layer chromatography (TLC) on a preparation plate (50 × 100 mm, Silica gel GF254; Qingdao Haiyang Chemical Co., Ltd., Shandong, China) using CHCl3–CH3OH–H2O = 65:35:10 (lower phase) as developing solvent. The structures of the purified compounds were identified by electrospray ionization mass spectrometry (ESI-MS; HP 1100 LC-MSn Trap SL System; Agilent, USA) and nuclear magnetic resonance (NMR) analysis (Bruker ARX 400 MHz spectrometer; Bruker Corp., Switzerland).
Purified saponin (1 ml, 0.05 mg l−1) was incubated with 1 ml crude microbial enzyme obtained from 96 h fermentation broth in each 10-ml tube at 55°C for various times. Every 24 h, a tube was taken out and the saponins were extracted with 1 ml n-butanol and detected with high-performance liquid chromatography (HPLC)–ESI-MS.
Analysis
Biomass
Because of the presence of PDZW, the biomass in each flask during the microbial transformation could not be detected directly. Intracellular protein concentration was measured using the method proposed by Zhang et al. (2007) with a UV/Vis spectrophotometer (APECORD 200; Analytik Jena, Germany). The biomass was quantified by reference to the calibration curve y = 6.21x + 1.08, R 2 = 0.9021, where x is intracellular protein concentration (grams per liter) and y is biomass (grams per liter).
α- and β-glucosidase
The broth from a fermentation sample in each flask was centrifuged at 4°C and 10,000 rpm for 15 min, and the suspension enzyme extract was harvested for activity analysis. The α- and the β-glucosidase activity were determined, respectively, using 5 mM p-nitrophenyl-α-d-glucopyranoside and 5 mM p-nitrophenyl-β-d-glucopyranoside as substrates (Sigma Aldrich Fine Chemical Co., Ltd., Shanghai, China). One milliliter of appropriately diluted raw enzyme solution was incubated with 1 ml of substrate in 0.1 M citrate buffer (pH 4.8) at 50°C in a water bath for 10 min. The reaction was stopped by adding 1 ml of 1 M Na2CO3. The p-nitrophenol generated was determined at 420 nm spectrophotometry. One unit (IU) of activity was defined as the amount of enzyme that released 1 μmol of p-nitrophenol per minute under the assay conditions.
Diosgenin
The whole fermentation sample medium in each flask (100 ml) was centrifuged, dried at 60°C, extracted with 50 ml of chloroform, and ultrasonicated for 30 min (KQ3200B ultrasonicator; Kunshan Ultrasonic Equipment Co., Ltd., Jiangsu, China), followed by filtration. The diosgenin concentration in filtrate was quantitatively determined by HPLC (Huang et al. 2008).
Diosgenin yield (percent) was calculated with the equation:
where Y is diosgenin yield (percent), D 0 is the diosgenin content in 9.77 g substrate before treatment, and D m and D a are diosgenin content in 9.77 g substrate after microbial and acid treatment, respectively.
Acid hydrolysis conditions: 9.77 g substrate was mixed with 30 ml 1 M H2SO4 and autoclaved at 121°C for 3 h, then filtered and washed with water until it became neutral. The filtrate residue was dried at 60°C and its diosgenin content was determined.
Saponins
TLC was applied to analyze the change of saponins in PDZW during the fermentation process. The ethanol sample extracts (obtained as for the chloroform extracts) were deposited on silica gel 254 plates (Merk, Germany) using chloroform–methanol–water (65:35:10, low phase) as developing solvent. The reaction products were visualized by soaking rapidly with ethanol solution containing 10% (vol/vol) sulfuric acid and heating at 110°C for 2 min.
HPLC–ESI-MS
HPLC–ESI-MS analysis was carried out on an Agilent 1100 unit (Agilent, USA) with LC–MS equipped with an electrospray interface (HP 1100 LC-MSn Trap SL System; Agilent, USA) in negative ionization mode (0–16 min) and positive ionization mode (16–25 min) using a Zorbax Eclipse XDB-C18 LC column (Agilent 2.1 × 150 mm, particle size 5 μm, pore size 80 Å, monomeric, double-endcapped) at 40°C. The mobile phase was methanol/water 85:15 (vol/vol) with a flow rate of 1 ml min−1 for 25 min.
Results
Enzymatic saccharification
The major function of enzymatic saccharification phase was to exclude starch from DZW tubers in the form of sugar. Chemical components in DZW and PDZW were analyzed to evaluate the saccharification efficiency (Table 1). DZW tubers contained 37.5% starch and 3.03% saponins, while about 2.11% starch and 9.25% saponins were detected in PDZW. We obtained 0.36 g of PDZW per 1 g treated DZW tuber. During the saccharification process, 98.0% starch was excluded from tubers, the content of saponins increased by 9.90% as the saponins were released from the network structure of starch. This explanation was investigated by examination of the morphological changes of the DZW before and after enzymatic pretreatment (Fig. 1). Starch in DZW (Fig. 1a) was oval to ellipsoid in shape with an unsmooth surface as it contained some saponins. After pretreatment, little starch was existed in PDZW (Fig. 1b) and some granule with smaller size was detected. With enzymatic saccharification, 0.24 g sugar was obtained from 1 g DZW.

SEM micrographs of D. zingiberensis C. H. Wright (a) tubers and pretreated D. zingiberensis C. H. Wright (b) tubers

Time course of reducing sugar, biomass, β-glucosidase, α-glucosidase, and diosgenin in shake flask cultivation of PDZW (square) and DZW (circle) by T. reesei. Data were expressed as mean values. The standard deviations were less than 10%
Microbial transformation
Shake flask cultivation
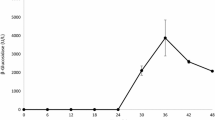
In the microbial transformation phase, it was noted that the concentration of reducing sugar, biomass, β-glucosidase, α-glucosidase, and diosgenin yield changed in an interesting way (Fig. 2). When T. reesei was grown on PDZW, reducing sugar in suspension decreased during the first 48 h, and the microorganism utilized this carbon source to build up its own biomass, growing exponentially. At 72 h, most of the reducing sugar was excluded from the zymotic fluid, and this lack of a carbon source induced the culture to produce β-glucosidase and α-glucosidase to hydrolyze sugar chains from saponins, leading to an increase in reducing sugar and diosgenin yield. In this phase, the fungal biomass increased slowly, reached a maximum at 84 h, and then decreased slightly. The highest α- and β-glucosidase activity was obtained at 84 and 96 h, whereas the maximum diosgenin yield of 90.6 ± 2.45% (83.9 ± 3.47 mg g−1 PDZW, 30.2 ± 1.25 mg g−1 DZW) occurred after 156 h of fungal growth.
The change of saponins with time during microbial transformation was also estimated (Fig. 3). Before bioconversion, four main saponins (Fig. 3 S0, compounds 1–4) were separated from PDZW by prepared TLC and identified as diosgenin-3-O-{[β-d-glucopyranosyl(1→4)]-[α-l-rhamnopyranosyl(1→4)]-[α-l-rhamnopyranosyl(1→2)]}-β-d-glucopyranoside, diosgenin-3-O-{[α-l-rhamnopyranosyl (1→2)]-[β-d-glucopyranosyl(1→3)]}-β-d-glucopyranoside, diosgenin-3-O-{[α-l-rhamnopyranosyl(1→2)]-[α-l-rhamnopyranosyl(1→4)]}-β-d-glucopyranoside, and diosgenin (see supplemental material Figs. S1, S2, and S3). In the fermentation phase with T. reesei, compounds 1–3 decreased gradually, while the content of compound 4 increased. Furthermore, one new compound (Fig. 3 S6.5) appeared with a high R f value and was verified as diosgenin-3-O-β-d-glucopyranoside (compound 5). The structures of compounds 1–5 were shown in Fig. 4. Generally, most saponins in PDZW were in the form of glycoside and transformed to aglycone during fermentation by T. reesei.

TLC of time course of saponins in shake flask cultivation of PDZW by T. reesei. Lanes S0, S3, S4, S5, S6, S6.5, and S7 show the EtOH extract of PDZW after incubation with T. reesei for 0, 72, 96, 120, 144, 156, and 168 h. Lane A shows the EtOH extract of acid hydrolyzed PDZW. Lane S shows the diosgenin standard. 1, 2, 3, and 5 indicate saponins, and 4 indicates diosgenin

Proposed microbial transformation pathways of compounds 1–3 to diosgenin (compound 4)
The microbial transformation pathway (Fig. 4) of compounds 1–3 was further studied (see supplemental material Figs. S4, S5, and S6). The hydrolysis of compound 1 was accomplished with three steps: firstly at the terminal β-d-glucosidic bond, secondly at the α-1,2-linked or α-1,4-linked glucosidic bond in the sugar chain at the C-3 position, and thirdly at the β-glycosidic bond which links β-d-glucose with the aglycone. The hydrolysis of compound 2 was commenced at the end β-1,3-glucosidic bond, followed at the α-1,2-glycosidic in the sugar chain, and finalized at the β-glycosidic bond at C-3 position, and then diosgenin was released. The hydrolyzing pathway of compound 3 was similar to that of compound 1.
Batch fermentation
Scaling up was then conducted to bioconvert saponins at a large scale. Under optimum conditions, a 91.2 ± 3.21% yield of diosgenin (84.5 ± 4.37 mg g−1 PDZW, 30.4 ± 1.57 mg g−1 DZW) was obtained.
Discussion
Starch accounts for ∼40% of the total mass of DZW tubers; it is an available carbon source in the fermentation industry but wasted in traditional microbial transformation methods (Qi et al. 2009; Liu et al. 2009). Sugars resulting from enzymatic saccharification process can be used as a carbon source for the production of ethanol, lactic acid, citric acid, and vitamin B2 (Wang et al. 2008; Huang et al. 2008).
The effect of saccharification on the subsequent microbial transformation was estimated. When DZW tubers were used as substrates, high biomass and low enzyme activity were derived (Fig. 2), compounds 1–3 were detected in the microbial hydrolyzate, and a low diosgenin yield resulted (56.4 ± 1.79%, Fig. 5 S1–S4). Incubation under the same conditions, using PDZW as substrate, increased the diosgenin yield by 42.4% to reach 90.6 ± 2.45%. Two reasons accounted for the increasing. First, with amylase, saponins were released from the network structure of starch (Table 1; Fig. 1), which made them more accessible to enzymes (Zhang et al. 2009). Second, starch in DZW is a more attractive carbon source for microorganisms and protects saponins from being hydrolyzed by the strain. This explanation was confirmed by the decrease of diosgenin yield with the addition of glucose to the fermentation medium of PDZW (Fig. 5 S5). The result was also supported by similar findings in the biotransformation of protopanaxadiol-type saponins, during which saponins usually acts as the only carbon source (Hasegawa et al. 1996, 1997; Bae et al. 2003).

TLC of saponins in DZW (S1); microbial hydrolyzate of DZW (S2); PDZW (S3); microbial hydrolyzate of PDZW obtained in medium 1 (S4; 2.67% peptone, 0.29% K2HPO4, 0.73% Tween 80) and medium 2 (S5; 2% glucose, 2.67% peptone, 0.29% K2HPO4, 0.73% Tween 80). 1, 2, 3, and 5 indicate saponins, and 4 indicates diosgenin
During microbial transformation, sugar chains, which connected to the C-3 hydroxyl group in saponins, were cleaved stepwise from the terminal sugar until diosgenin was released. The use of microorganisms for production of modified saponins has been reported extensively (Wang et al. 2007a; Chen et al. 2008; Cheng et al. 2008). These studies mostly use monosaponin as substrate and yield multiple products. In our study, multiple substrates were bioconverted to a single product. This required a more complicated enzyme system. Enzymes produced by T. reesei not only exhibited an excellent performance on cleaving the terminal β-d-glucose or α-l-rhamnose in sugar chain but also hydrolyzed the β-glycosidic linkage between aglycone and β-d-glucose effectively. The stereo- and regioselectivities of the enzymes in this system are worth further study.
In conclusion, with respect to resource utilization and clean production, a promising approach for preparation of diosgenin from DZW tubers was accomplished by a successive process of enzymatic saccharification and microbial transformation. Results showed that enzymatic saccharification was an effective method for excluding starch from raw herb and enhancing the subsequent microbial transformation. Saponins in PDZW were converted to diosgenin effectively by T. reesei with a diosgenin yield of 91.2 ± 3.21% in the 5.0-l bioreactor. The described process provides an attractive alternative for clean diosgenin production in industry.
Notes
The fermentation medium components and condition parameters in this study were optimized for high diosgenin yield and will be reported later.
References
Adham NZ, Zaki RA, Naim N (2009) Microbial transformation of diosgenin and its precursor furostanol glycosides. World J Microbiol Biotechnol 25:481–487
Bae EA, Kim NY, Han MJ, Choo MK, Kim DH (2003) Transformation of ginsenoside to compound K (IH-901) by lactic acid bacteria of human intestine. J Microbiol Biotechnol 13:9–14
Bertranda J, Liagreb B, Bégaud-Grimaudc G, Jauberteaua MO, Beneytoutb JL, Cardot PJP, Battuc S (2009) Analysis of relationship between cell cycle stage and apoptosis induction in K562 cells by sedimentation field-flow fractionation. J Chromatogr B 877:1155–1161
Cao YF (2004) Studies on the morphogenesis and the development of the structure of rhizome of Dioscorea zingiberensis and their relationship with the accumulation of diosgenin. Northwest University, Xi’an (in Chinese)
Chen GT, Yang M, Song Y, Lu ZQ, Zhang JQ, Huang HL, Wu LJ, Guo DA (2008) Microbial transformation of ginsenoside Rb1 by Acremonium strictum. Appl Microbiol Biotechnol 77:1345–1350
Cheng LQ, Na JR, Bang MH, Kim MK, Yang DC (2008) Conversion of major ginsenoside Rb1 to 20(S)-ginsenoside Rg3 by Microbacterium sp. GS514. Phytochemistry 69:218–224
Cheng P, Zhao HZ, Zhao B, Ni JR (2009) Pilot treatment of wastewater from Dioscorea zingiberensis C.H. Wright production by anaerobic digestion combined with a biological aerated filter. Bioresour Technol 12:2918–2925
Chiang CT, Way TD, Tsai SJ, Lin JK (2007) Diosgenin, a naturally occurring steroid, suppresses fatty acid synthesis expression in HER2-over expressing breast cancer cells through modulating Akt, mTOR and JNK phosphorylation. FEBS Lett 581:5735–5742
Feng B, Ma BP, Kang LP, Xiong CQ, Wang SQ (2005) The microbiological transformation of steroidal saponins by Curvularia lunata. Tetrahedron 61:11758–11763
Feng B, Hu W, Ma BP, Wang YZ, Huang HZ, Wang SQ, Qian XH (2007) Purification, characterization and substrate specificity of a glucoamylase with steroidal saponin-rhamnosidase activity from Curvularia lunata. Appl Microbiol Biotechnol 76:1329–1338
Fernandes P, Cruz A, Angelova B, Pinheiro HM, Cabral JMS (2003) Microbial conversion of steroid compounds: recent developments. Enzyme Microb Technol 32:688–705
Hasegawa H, Sung JH, Matsumiya S, Uchiyama M (1996) Main ginseng saponin metabolites formed by intestinal bacteria. Planta Med 62:453–457
Hasegawa H, Sung JH, Benno Y (1997) Role of human intestinal Prevotella oris in hydrolyzing ginseng saponins. Planta Med 63:436–440
He XJ, Qiao A, Liu B, Wang XL, Wang GH, Qu GX, Liu RH, Yao XS (2006) Bioconversion of methyl protodioscin by Penicillium melinii cells. Enzyme Microb Technol 38:400–406
Huang W, Zhao HZ, Ni JR, Zuo H, Qiu LL, Li H (2008) The best utilization of D. zingiberensis C. H. Wright by an eco-friendly process. Bioresour Technol l99:7407–7411
Jayachandran KS, Vasanthi HR, Rajamanickam GV (2009) Antilipoperoxidative and membrane stabilizing effect of diosgenin, in experimentally induced myocardial infarction. Mol Cell Biochem 327:203–210
Jayadev R, Ranjana PB (2007) Diosgenin, a naturally occurring furostanol saponin suppresses 3-hydroxy-3-methylglutaryl CoA reductase expression and induces apoptosis in HCT-116 human colon carcinoma cells. Cancer Lett 255:194–204
Liagre B, Vergne-Salle P, Corbiere C, Charissoux JL, Beneytout JL (2004) Diosgenin, a plant steroid, induces apoptosis in human rheumatoid arthritis synoviocytes with cyclooxygenase-2 overexpression. Arthritis Res Ther 6:R373–R383
Liu L, Dong YS, Qi SS, Wang H, Xiu ZL (2009) Biotransformation of steroidal saponins in Dioscorea zingiberensis C. H. Wright to diosgenin by Trichoderma harzianum. Appl Microbiol Biotechnol. doi:10.1007/s00253-00902098-1
Oncina R, Botía JM, Del Río JA, Ortuño A (2000) Bioproduction of diosgenin in callus cultures of Trigonella foenum-graecum L. Food Chem 70:489–492
Qi SS, Dong YS, Zhao YK, Xiu ZL (2009) Qualitative and quantitative analysis of microbial transformation of steroidal saponins in Dioscorea zingiberensis. Chromatographia 69:865–870
Qian SH, Yuan LH, Yang NY, OuYang PK (2006) Study on steroidal compounds from Dioscorea zingiberensis. Chinese Traditional Herbal Drugs 29:1174–1176 (in Chinese)
Saunders R, Cheetham PSJ, Hardman R (1986) Microbial transformation of crude fenugreek steroids. Enzyme Microb Technol 8:549–555
Trouillas P, Corbiere C, Liagre B, Duroux JL, Beneytout JL (2005) Structure–function relationship for saponin effects on cell cycle arrest and apoptosis in the human 1547 osteosarcoma cells: a molecular modeling approach of natural molecules structurally close to diosgenin. Bioorg Med Chem Lett 13:1141–1149
Wang FQ, Li B, Wang W, Zhang CG, Wei DZ (2007a) Biotransformation of diosgenin to nuatigenin-type steroid by a newly isolated strain, Streptomyces virginiae IBL-14. Appl Microbiol Biotechnol 77:771–777
Wang H, Liu L, Guo YX, Dong YS, Zhang DJ, Xiu ZL (2007b) Biotransformation of piceid in Polygonum cuspidatum to resveratrol by Aspergillus oryzae. Appl Microbiol Biotechnol 75:763–768
Wang YX, Liu H, Bao JG, Hong Y, Yang ZH, Zhang CX (2008) The saccharification-membrane retrieval-hydrolysis (SMRH) process: a novel approach for cleaner production of diosgenin derived from Dioscorea zingiberensis. J Cleaner Prod 16:1133–1137
Yayoi T, Naoko K, Akinori H, Megumi T, Hideyo U, Shinichi W (2009) Novel effects of diosgenin on skin aging. Steroids 74:504–511
Zhao YT, Xu ZL, Dai CC, Tang XL, Xia B, Wang Q (2007) Selection of the microorganism for dioscin hydrolyze. Journal of Chinese Medical Material 30:905–909 (in Chinese)
Zhao HZ, Cheng P, Zhao B, Ni JR (2008) Yellow ginger processing wastewater treatment by a hybrid biological process. Process Biochem 43:1427–1431
Zhang CX, Wang YX, Yang ZH, Xu MH (2006) Chlorine emission and dechlorination in co-firing coal and the residue from hydrochloric acid hydrolysis of Dioscorea zingiberensis. Fuel 85:2034–2040
Zhang Q, Lo CM, Ju LK (2007) Factors affecting foaming behavior in cellulase fermentation by Trichoderma reesei Rut C-30. Bioresour Technol 98:753–760
Zhang YQ, Tang LR, An X, Fu EH, Ma CF (2009) Modification of cellulase and its application to extraction of diosgenin from Dioscorea zingiberensis C. H. Wright. Biochem Eng J. doi:10.1016/j.bej.2009.07.006
Acknowledgments
This work was supported by the National “11th-5-Year” Plan Project of China (2006BAB04A14).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary material
(DOC 312 kb)
Rights and permissions
About this article
Cite this article
Zhu, YL., Huang, W., Ni, JR. et al. Production of diosgenin from Dioscorea zingiberensis tubers through enzymatic saccharification and microbial transformation. Appl Microbiol Biotechnol 85, 1409–1416 (2010). https://doi.org/10.1007/s00253-009-2200-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-009-2200-8