
Abstract
Amino acids are among the major products in biotechnology in both volume and value, and the global market is growing. Microbial fermentation is the dominant method used for industrial production, and today the most important microorganisms used are Corynebacteria utilizing sugars. For low-prize bulk amino acids, the possibility of using alternative substrates such as methanol has gained considerable interest. In this mini review, we highlight the unique genetics and favorable physiological traits of thermotolerant methylotroph Bacillus methanolicus, which makes it an interesting candidate for overproduction of amino acids from methanol. B. methanolicus genes involved in methanol consumption are plasmid-encoded and this bacterium has a high methanol conversion rate. Wild-type strains can secrete 58 g/l of l-glutamate in fed-batch cultures at 50°C and classical mutants secreting 37 g/l of l-lysine have been selected. The relative high growth temperature is an advantage with respect to both reactor cooling requirements and low contamination risks. Key genes in l-lysine and l-glutamate production have been cloned, high-cell density methanol fermentation technology established, and recently a gene delivery method was developed for this organism. We discuss how this new knowledge and technology may lead to the construction of improved l-lysine and l-glutamate producing strains by metabolic engineering.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Amino acids are used as food and feed supplements, pharmaceuticals, cosmetics, polymer materials, and agricultural chemicals (Ikeda 2003; Faurie and Thommel 2003; Marx et al. 2006). The most important industrial amino acid producer today is the bacterium Corynebacterium glutamicum, which produces about 2 million tons of amino acids per year, above 1 and 0.6 million tons of l-glutamate and l-lysine, respectively (Eggeling and Bott 2005). Several decades of extensive research has resulted in detailed data revealing C. glutamicum metabolism and physiology, and its genome sequence was recently published (Ikeda and Nakagawa 2003; Kalinowski et al. 2003), and also that of the closely related and more thermotolerant Corynebacterium efficiens (Nishio et al. 2003). Genetic tools are well-developed, and this knowledge and technology has been used to generate efficient C. glutamicum amino acid overproducers by metabolic engineering (see Ohnishi et al. 2002; Eggeling and Bott 2005 and references therein). The substrates for C. glutamicum fermentation are generally sugar from agricultural crops.
Methanol is an alternative substrate to sugar for the production of chemical intermediates and commodity microbial products
There is a growing global demand for amino acids and the possibilities to utilize alternative substrates as feedstock in fermentation have therefore gained considerable interest. One-carbon (C1) compounds occur abundantly throughout nature, and methane and methanol are two of the most important C1 compounds from a biotechnological and a bulk chemical point of view (Linton and Niekus 1987; Olah et al. 2006). Compared to molasses, for example, methanol is a pure raw material that can be completely utilized during bacterial fermentations. Today, almost all methanol worldwide is produced from synthesis gas (syn-gas, a mixture of CO and H2) obtained from the incomplete combustion of natural gas. However, new ways for its production directly from natural gas without going through syn-gas and by hydrogenative chemical recycling of CO2 (e.g., from industrial exhausts of fossil fuel burning plants) are being developed. Methanol can also be prepared from biomass, but these processes play only a minor role (Olah et al. 2006). As for any other commodities, methanol prices vary depending on supply and demand, and the gas price is linked to the price of crude oil. Since 1975, the average wholesale price for methanol has fluctuated between 100 and 350 USD per ton (Olah et al. 2006). The supply and, thus, the price of sugar will vary depending on the weather conditions in the major sugar producing regions, as well as agricultural politics. Data collected from the United States Department of Agriculture (http://www.ers.usda.gov/Briefing/Sugar/data.htm) and from Methanex (http://www.methanex.com/products/methanolprice.html) show that over the last 6 years, the price of methanol has been similar to that of raw sugar (Fig. 1). However, mega-methanol production facilities (5,000 tons/day) are now being constructed in regions rich in natural gas, such as the Caribbean, Latin America, and the Middle East. The production costs for methanol in mega-methanol plants have been estimated to be well below 100 USD per ton, and they are expected to allow the price of methanol to remain at a relatively low level as long as natural gas reserves are available. In regions rich in coal such as USA and China, large-scale production of methanol from coal may provide an alternative domestic route for methanol (Olah et al. 2006). The price of molasses as a substrate varies both with geographical location and with the sugar crop and is typically between 50 and 100% of the price of raw sugar on a fermentable sugar basis.

Comparison of the recent monthly price fluctuations of methanol (solid line) and raw sugar (dashed line). Data for the US Gulf Coast from Methanex and from the United States Department of Agriculture (see text)
Amino acid production from methanol by methylotrophs
Methylotrophs comprise the large number of both aerobic and anaerobic microorganisms that can grow on reduced compounds lacking C–C bonds, such as methane and methanol (Anthony 1982; Large and Bamforth 1988). Obligate methylotrophs can exclusively utilize C1 compounds as a sole carbon and energy source, while facultative methylotrophs can utilize both C1 and multicarbon compounds. Genetic tools for many methylotrophs have been established, and engineering of methylotrophs leading to overproduction of different amino acids are reported including l-serine (Izumi et al. 1993; Hagishita et al. 1996), l-threonine (Motoyama et al. 1994), l-glutamate (Motoyama et al. 1993), and l-lysine (Motayama et al. 2001). Representative l-lysine and l-glutamate producing methylotrophs reported in the literature are listed in Table 1. For example, in the Gram-negative obligate methylotroph Methylophilus methylotrophus, the expression of a mutant gene encoding dihydrodipicolinate synthase deregulated in l-lysine inhibition caused increased l-lysine synthesis to about 1 g/l at 37°C (Tsujimoto et al. 2006). By co-expressing a mutant gene encoding an l-lysine transporter they obtained a recombinant strain, strain AS1 (pSEA10), secreting 11.3 g/l of l-lysine from methanol (Gunji and Yasueda 2006). A recombinant mutant, AL119 (pDYOM4-2), of the Gram-negative obligate methylotroph Methylobacillus glycogenes, overexpressing a dihydrodipicolinate synthase partly deregulated in l-lysine inhibition was reported to produce about 8 g/l of l-lysine and 37 g/l of l-glutamate from methanol at 37°C (Motayama et al. 2001). To our knowledge, no commercial methanol-based industrial production process of any amino acids exists; however, this may change in the future based on the success of engineering efficient methylotrophic amino acid-producing strains and also based on cost and supply of methanol vs sugar substrates.
The candidate bacterium Bacillus methanolicus
The methylotrophic bacterium B. methanolicus has many unique traits that make it an important candidate for the bioconversion of methanol into amino acids. In this mini review, we will focus on scientific progress that has been made leading to a better understanding of B. methanolicus physiology and biology, particularly related to the pathway of methanol assimilation, methanol and formaldehyde tolerance, and metabolic routes for production of the amino acids l-lysine and l-glutamate. The ability to use metabolic engineering to create recombinant strains with improved production properties was delayed mainly due to the lack of functional genetic tools. However, a method for constructing recombinant B. methanolicus strains that retained the ability to grow on methanol was recently established, and the current progress and strategies for generating l-lysine and l-glutamate overproducers by metabolic engineering of B. methanolicus is discussed.
The isolation of thermotolerant methylotrophic bacilli later designated B. methanolicus
A mixed culture growing on methanol containing spore-forming bacteria was reported in the 1970s (Snedecor and Cooney 1974), and several reports on isolation of thermotolerant and methylotrophic bacilli appeared in the scientific literature in the late 1980s (Al-Awadhi et al. 1988, 1989; Dijkhuizen et al. 1988; Brooke et al. 1989). Some of these strains were initially classified as Bacillus brevis, but were later reclassified as B. methanolicus (Arfman et al. 1992a). B. methanolicus is a Gram-positive aerobic methylotroph growing at temperatures from 35 to 60°C. These strains can readily be isolated from soil samples, wastewater treatment systems, and volcanic hot springs. The cells are rod-shaped, and they may form oval endospores, but B. methanolicus cells typically sporulate poorly in liquid growth medium at 50°C (Schendel et al. 1990). Most strains of B. methanolicus can grow in 2% but not in 5% NaCl (Arfman et al. 1992a), and wild-type strains such as MGA3 and NOA2 require biotin and vitamin B12 for growth (Schendel et al. 1990). Adaptation to rapid growth in artificial or natural seawater-based media has been reported (Komives et al. 2005), indicating that this bacterium has low cost requirements regarding growth components. Some wild-type strains with high salt tolerance (DFS2, HEN9) that can grow in the presence of 8% ammonium glutamate have been isolated from marsh soil (Brautaset et al. 2004). B. methanolicus strains are restricted methylotrophs, meaning that they can utilize few alternative carbon sources, indicating that methylotrophy is an important metabolic trait of this organism in nature (Arfman et al. 1992a,b).
B. methanolicus uses the ribulose monophosphate pathway for C1 assimilation
In the cells methanol is converted into formaldehyde catalyzed by a methanol dehydrogenase (see below). Among methylotrophs four different routes for formaldehyde assimilation are known (Anthony 1982, 1991; Large and Bamforth 1988; Dijkhuizen et al. 1992). Common to these pathways is that C1 compounds are fixed to molecules that are regenerated by the completion of one cycle; the product is one three-carbon compound synthesized from three C1 compounds. B. methanolicus assimilates formaldehyde by the ribulose monophosphate (RuMP) pathway (De Vries et al. 1990; Kato et al. 2006), which can be divided into three parts (Fig. 2). In the fixation part (Fig. 2a), three formaldehyde molecules are condensed with three molecules of ribulose 5-phosphate (Ru5P) and catalyzed by 3-hexulose-6-phosphate synthase (HPS) to make three hexulose 6-phosphate (H6P) molecules. One of the H6P molecules is further converted to fructose 6-phosphate (F6P) catalyzed by 6-phospho-3-hexuloisomerase (PHI). Two variants of the cleavage part exist, and B. methanolicus uses the fructose-1,6-bisphosphate aldolase (FBPA) variant, where F6P is converted by phosphofructokinase (PFK) to fructose-1,6-bisphosphate (FBP) at the expense of one ATP (Fig. 2b). FBP is then cleaved by FBPA to glyceraldehyde-3-phosphate (GAP) and dihydroxyacetone phosphate (DHAP). DHAP can be converted by glycolytic steps into pyruvate, generating one NAD(P)H and two ATP. GAP, together with two H6P molecules, enters the regeneration part of the pathway, where three Ru5P molecules are regenerated. Two variants of the regeneration part exist (Fig. 2c), and they share the three enzymes transketolase, ribose-5-phosphate isomerase, and ribulose-5-phosphate 3-epimerase (RPE). The difference between the variants is the usage of transaldolase (TA variant) or a sedoheptulose-1,7-bisphosphatase (SBPase variant). Energetically, all four possible variants of the RuMP pathway produce one NAD(P)H per pyruvate generated, and the most favorable combination is the FBPA/TA variant, which in addition yields one ATP.

Outline of the RuMP pathway and illustration of the plasmid-dependent methylotrophy in B. methanolicus. a Upon the completion of one cycle, three formaldehyde (FA) molecules are converted into one three-carbon molecule. The enzymes of the fixation part (see text) are indicated. b Two variants of the cleavage part of the RuMP pathway. The FBPA variant converts F6P to GAP and DHAP. Further conversion of DHAP to pyruvate by glycolytic enzymes is indicated. Alternatively, the 2-keto-3-deoxy-6-phosphogluconate aldolase (KDPGA) variant converts F6P to GAP and pyruvate using enzymes of the Entner–Doudoroff pathway. c Two variants of the regeneration part of the RuMP pathway (see text). d Genetic organization of mdh and the totally seven different RuMP pathway genes identified in B. methanolicus MGA3. B. methanolicus genes and gene products of plasmid (grey background) and chromosomal (black background) origin are shown. Genes: mdh MDH, glpX SBPase, fba FBPA, tkt TKT, pfk PFK, rpe RPE, hps HPS, phi PHI. Enzymes: PGI Phosphoglucose isomerase, GPDH glucose-6-phosphate dehydrogenase, PGD 6-Phosphogluconate dehydratase. Abbreviations: X5P xylulose-5-phosphate, Ri5P ribose-5-phosphate, S7P sedoheptulose-7-phosphate, SBP sedoheptulose-1,7-bisphosphate, E4P erythrose-4-phosphate, 6PG 6-phosphogluconate, G6P glucose-6-phosphate, KDPG 2-keto-3-deoxy-6-phosphogluconate. For other abbreviations, see text
Methanol oxidation and formaldehyde assimilation by B. methanolicus involves the concerted induction of chromosomal and plasmid-encoded genes
B. methanolicus uses an NAD-linked methanol dehydrogenase (MDH; Arfman et al. 1989) to oxidize methanol into formaldehyde, and the B. methanolicus C1 mdh gene has been cloned by De Vries et al. (1992). Interestingly, 12 years later (Brautaset et al. 2004), it was shown that mdh, together with five RuMP pathway genes, are plasmid-borne in B. methanolicus (see below). The mdh gene encodes a 385-amino acid polypeptide representing a type III NAD-dependent alcohol dehydrogenase. MDH activity involves an activator protein denoted ACT, and the genetic and biochemical properties of ACT and MDH have been well-characterized (Arfman et al. 1997; Hektor et al. 2002; Kloosterman et al. 2002). In contrast to mdh, the act gene has a chromosomal origin in B. methanolicus (Brautaset et al. 2004). The MDH protein is a decameric nicotinoprotein, and each subunit contains a bound NAD(H) cofactor, one Zn2+ ion and one or two Mg2+ ions. The in vitro MDH activity is about 40 times higher when purified ACT protein is added; ACT activation involves hydrolytic removal of the nicotinamide mononucleotide moiety of NAD(H) (Kloosterman et al. 2002). Increased NADH/NAD ratios in the cells reduce ACT-mediated activation of MDH, presumably acting as a control mechanism for preventing accumulation of toxic formaldehyde in the cells. The mdh gene is transcribed at very high levels in wild-type B. methanolicus MGA3 cells growing both methylotrophically on methanol or non-methylotrophically on mannitol (Jakobsen et al. 2006). MDH can constitute up to 30% of total protein in the cells (Arfman et al. 1989). These findings indicate that this organism retains a high capacity to oxidize methanol under different nutritional conditions.
B. methanolicus plasmid pBM19 carries mdh and five RuMP pathway genes
The 19,167-bp and medium-copy number plasmid pBM19 in B. methanolicus MGA3 carries mdh and five RuMP pathway genes (glpX, class II SBPase; fba, class II FBPA; tkt, TKT; pfk, PFK; rpe, RPE; Fig. 2d). Genetic characterization of 13 different wild-type strains indicated that pBM19-like plasmids are common among B. methanolicus species (Brautaset et al. 2004). Two more genes representing the fixation part of the RuMP pathway, hps (encoding HPS activity) and phi (encoding PHI activity), were found on the chromosome organized as an operon (Fig. 2d). By using quantitative polymerase chain reaction, it was demonstrated that these plasmid and chromosomal RuMP pathway genes are 6- to 40-fold upregulated in cells growing on methanol compared to those growing on mannitol (Jakobsen et al. 2006), and curing of pBM19 resulted in loss of the ability to grow on methanol. This represents experimental evidence that B. methanolicus methylotrophy is plasmid-linked. Based on enzyme activity measurements in crude extracts, B. methanolicus was originally reported to use the TA regeneration variant of the RuMP pathway (Arfman et al. 1989; Dijkhuizen et al. 1992). However, considering the predicted functions of the identified RuMP pathway genes, it seems likely that B. methanolicus may use the SBPase regeneration variant (Fig. 2c). The induced state of the RuMP pathway genes is important for the methanol and formaldehyde tolerance level of the cells (see below), and cells pregrown on mannitol or in any other rich media lacking methanol must be gradually adapted to increasing methanol concentrations to avoid cell death (Jakobsen et al. 2006).
pBM19 is crucial for methylotrophic growth and a burden upon non-methylotrophic growth
Comparative growth studies using the pBM19-cured B. methanolicus strain MGA3C-A6 (Brautaset et al. 2004) and its parental wild-type strain, unravelled new and interesting physiological traits of B. methanolicus. MGA3C-A6 cannot grow on methanol, while it has a significantly higher growth rate on mannitol compared to the wild-type (0.37 h−1 vs 0.30 h−1 in shake flasks), indicating that pBM19 represents a burden for cells under growth conditions where its genes are not needed (Jakobsen et al. 2006). The latter assumption was confirmed by 140 generations of continuous growth of wild-type cells in mannitol medium resulting in loss of pBM19 from the culture. Mutant MGA3C-A6 growing on mannitol can tolerate the addition of about tenfold higher concentration of methanol and only half the concentration of formaldehyde compared to the wild-type grown under similar conditions. The high methanol tolerance was reduced to wild-type levels in recombinant MGA3C-A6 cells expressing MDH activity (Jakobsen et al. 2006). This indicates that methanol sensitivity of B. methanolicus is mainly caused by toxic accumulation of formaldehyde generated by the high MDH activity and that the RuMP pathway plays an important role also for the formaldehyde tolerance of the cells. By growing wild-type cells in a mixture of mannitol and methanol, the majority of the total carbon consumed derives from methanol. Under such conditions, the RuMP pathway genes are transcriptionally induced (Jakobsen et al. 2006).
Dissimilatory pathways for formaldehyde oxidation to CO2
Measurements in crude extracts of B. methanolicus have demonstrated phosphoglucose isomerase, glucose-6-phosphate dehydrogenase, and 6-phosphogluconate dehydrogenase activities (Fig. 3), indicating a cyclic dissimilatory pathway for oxidation of formaldehyde into CO2 (Arfman et al. 1989). Later, indication of an alternative pathway for formaldehyde dissimilation has also been reported using [13C] nuclear magnetic resonance by feeding labeled methanol (13CH3OH) to B. methanolicus (Pluschkell and Flickinger 2002). To verify the activity of formate dehydrogenase (FDH), pulses of 13C formate were added to a continuous culture at steady state and 13CO2 enrichment was monitored in the exhaust gas by isotope ratio mass spectroscopy (Pluschkell and Flickinger 2002). These data indicate that formaldehyde can also be oxidized to CO2 by a linear pathway via formate (Fig. 3), presumably catalyzed by formaldehyde dehydrogenase and FDH. In addition to generating reducing power, the latter pathway is assumed to play a role in regulating intracellular formaldehyde levels in the cells upon methanol growth. To date, no genes representing any of these two alternative dissimilatory pathways have been cloned from B. methanolicus.

Outline of the linear and cyclic dissimilatory pathways for formaldehyde. The linear dissimilatory pathway converts formaldehyde (FA) to CO2 by the action of FADH and FDH. The cyclic dissimilatory pathway is a combination of the fixation part of the RuMP pathway and the pentose phosphate pathway. The remaining parts of the RuMP pathway are here indicated in grey. Enzymes: PGDH 6-Phosphogluconate dehydrogenase. For other enzymes and abbreviations, see text and legend under Fig. 2
Methanol consumption rate and theoretical product yield
High substrate consumption rates and high product yields are desirable for all microbial production processes. Based on reported data from a 14-l fed-batch fermentation with B. methanolicus MGA3 in which 50 g dry cell weight per liter was achieved (Schendel et al. 1990), a mean methanol consumption rate of at least 7 g/h can be calculated. Theoretical product yields can be estimated based on assumed biochemical pathways of the organism. However, the dissimilatory metabolism of methanol is not fully elucidated, and as for l-lysine in particular, three different biosynthesis pathways are recognized in prokaryotes (Kindler and Gilvarg 1960; Sundharadas and Gilvarg 1967; White 1983; Berges et al. 1986). Comparisons of theoretical l-lysine yields from methanol by B. methanolicus and from glucose by C. glutamicum are given in Table 2. Most Bacillus species are reported to use the acetylase variant of the l-lysine biosynthesis pathway (Paulus 1993). Although all three variants of the pathway have been suggested to exist in C. glutamicum (Weinberger and Gilvarg 1970; Tosaka and Takinami 1978; Schrumpf et al. 1991), the succinylase and the dehydrogenase variants appear to be the most important (Sonntag et al. 1993; Wehrmann et al. 1998). A comparison of stoichiometric conversions of methanol or glucose to l-lysine is shown in Fig. 4. Table 2 and Fig. 4 show that the yield of l-lysine from methanol by B. methanolicus is similar (0.71 and 0.81 g l-lysine-HCl/g methanol) to the yield from glucose by C. glutamicum (0.68 and 0.82 g l-lysine·HCl/g glucose). Thus, B. methanolicus, with a high methanol consumption rate combined with high theoretical product yields should be a potentially efficient biocatalyst for the production of amino acids from methanol.

Comparison of stoichiometric conversion of methanol (CH3OH) or glucose (C6H12O6) to l-lysine (C6H14O2N2) by B. methanolicus and C. glutamicum, respectively. For B. methanolicus, the acetylase variant of the l-lysine biosynthesis pathway is assumed, while the succinylase and the dehydrogenase variants are assumed for C. glutamicum (see text). Different conversion mechanisms of formaldehyde to formate [distinguished by NAD(P)H generation] are known (Large and Bamforth 1988), giving rise to two possible conversion alternatives for l-lysine from methanol, indicated here by alternatives A and B. The synthesis of one l-lysine molecule from methanol by B. methanolicus or from glucose by C. glutamicum (using the dehydrogenase variant) require two molecules NAD(P)H, while the conversion of glucose to l-lysine by C. glutamicum using the succinylase variant is carbon-limited and yields NAD(P)H in surplus
Fed-batch high cell density cultivation methods at 50°C for B. methanolicus
As stated above, MDH levels in B. methanolicus are high under any conditions indicating that methanol, when available, will be rapidly converted into formaldehyde in the cells. Therefore, to control in vivo accumulation of toxic formaldehyde and achieve high B. methanolicus cell density, continuous feeding of methanol is required when cultivating this organism in a bioreactor. Online methanol analysis is used to keep the methanol level constant at a limited concentration (Schendel et al. 1990; Lee et al. 1996). Too rapid feeding of methanol results not only in formaldehyde toxicity, but also in a substantial fraction of the methanol carbon being dissimilated to CO2 (see Fig. 3), accompanied with an increase in oxygen demand (Pluschkell and Flickinger 2002). Methanol is more reduced than sugars, and methanol fermentations are therefore characterized by high oxygen demands. As heat evolution increases with oxygen consumption, cooling requirements are high for fermentations using methanol compared to sugar. Due to the thermotolerant phenotype of B. methanolicus, a dramatic reduction in cooling water is a major cost benefit of employing this organism compared to using methylotrophic bacteria growing at 30 to 37°C (Motayama et al. 2001; Tsujimoto et al. 2006). At about 200 m3 reactor liquid volume, the cooling water requirements for an organism growing on glucose at 35°C and an organism growing on methanol at 50°C are reported to be similar (Komives et al. 2005). Another effect of increased process temperature is lowered oxygen solubility in the medium. At atmospheric pressure, increasing the temperature from 35 to 50°C decreases the solubility of oxygen about 20%; therefore, there is an increased demand for oxygen supply in the fermentors under such growth conditions. Both laboratory scale fed-batch cultures and chemostats of B. methanolicus have been investigated, and dry cell mass of up to 50 g/l has been achieved (Schendel et al. 1990; Lee et al. 1996; Pluschkell and Flickinger 2002; Brautaset et al. 2003). Optimization of fermentation media composition, inoculum, online monitoring, and feeding strategies are critical to obtaining high and reproducible cell and amino acid yields in bioreactors. Due to the requirements to avoid formaldehyde toxicity and to supply sufficient oxygen, special attention must be paid to aeration and methanol feeding strategies when bioreactor volume increases to the scale used in industrial amino acid production (several hundred cubic meters). The design of large bioreactors should prevent non-homogeneous mixing and poor oxygen mass transfer resulting in localized formaldehyde toxicity and oxygen starvation.
l-Glutamate production by B. methanolicus strains
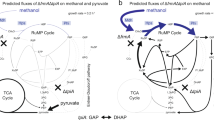
The flow of methanol-derived carbon from oxaloacetate (OAA) is split between transamination to aspartate and formation of citrate (Fig. 5). The citrate cycle (TCA cycle) of organisms harboring the linear dissimilation pathway of formaldehyde (see above) is assumed to play a minor role in energy metabolism during methylotrophic growth (Zatman 1981; Shishkina and Trotsenko 1982). In wild-type B. methanolicus MGA3, intracellular levels of l-glutamate are significant, ∼90 mM, and in fed-batch culture this strain can secrete up to 58 g/l of l-glutamate (Schendel et al. 2000; Brautaset et al. 2003; Table 1). This indicates that a substantial fraction of pyruvate enters the first reactions of the TCA cycle, leading to formation of 2-oxoglutarate (Fig. 5). The B. methanolicus citY gene, encoding an active citrate synthase (CS) II protein, has been cloned and sequenced, and chromosomal inactivation of this gene demonstrated that this bacterium has several forms of CS (Brautaset et al. 2003). 2-Oxoglutarate can be transaminated to l-glutamate or alternatively oxidized by 2-oxoglutarate dehydrogenase (ODHC) of the TCA cycle. Bacillussubtilis has no assimilatory glutamate dehydrogenase activity; ammonium assimilation occurs solely by the glutamine synthetase/glutamate synthase pathway (Fisher 1999). It remains unknown which of these two alternative mechanisms B. methanolicus uses for converting 2-oxaloacetate into l-glutamate. Interestingly, wild-type B. methanolicus MGA3 has low ODHC activity (Brautaset et al. 2003), which is likely one major reason for the high l-glutamate production by this organism. In C. glutamicum, effective l-glutamate export can be achieved by using Tween 40 (polyethylene sorbitan monooleate) or biotin limitations in the growth medium. In a screen of different surfactants only Tween 80 stimulated l-glutamate secretion in B. methanolicus (Hanson et al. 2000; Schendel et al. 2000), and biotin limitation does not increase l-glutamate secretion in this bacterium; it is also required for cell growth.

Overview of biochemical pathways from pyruvate to l-glutamate and l-lysine in B. methanolicus, with relevant enzymes and intermediates indicated (see text). The conversion of aspartate semi-aldehyde to l-lysine is indicated by the seven enzymatic steps of the acetylase/succinylase variants of the l-lysine biosynthetic pathway. It is not known which mechanism, glutamate dehydrogenase or glutamine synthase/glutamate synthetase, is employed by B. methanolicus (see text)
l-Lysine production by B. methanolicus strains
Generation of classical B. methanolicus mutants overproducing l-lysine
Researchers at the University of Minnesota constructed a series of classical mutants that overproduce various levels of l-lysine during the 1990s. For example, by using repetitive cycles of random mutagenesis and selection of B. methanolicus wild-type strain NOA2, mutant NOA2#13A52-8A66 was obtained. This homoserine auxotrophic mutant is resistant to the l-lysine analogue S-2-aminoethyl cysteine (AEC) and the diaminopimelic acid (DAP) analogue diaminobutyrate, and it requires supply of both threonine and methionine in the growth medium for rapid growth. Mutant NOA2#13A52-8A66 was capable of secreting 37 g/l l-lysine and 17 g/l l-glutamate in 11-l fed-batch methanol cultures (Lee et al. 1996; Hanson et al. 1996); to date, this is still the highest l-lysine production reported by any methylotrophic bacterium (Table 1).
CS and pyruvate carboxylase play important roles in C-flow from the key intermediate pyruvate
To achieve high l-lysine production, it is important that a high fraction of pyruvate is converted into aspartate (Fig. 5). Reducing CS levels by inactivation of citY, which encodes CSII, has been shown to strongly reduce l-glutamate production in B. methanolicus (Brautaset et al. 2003). Interestingly, the citY mutant strain could grow well on methanol medium as long as supplied with excess l-glutamate in the growth medium (Brautaset et al. 2003). This manipulation, however, had no positive effect on production rate or yield, most probably due to lack of deregulation of key enzymes in the l-lysine biosynthetic pathway. We believe that the citY-deficient mutant should be an interesting host for additional mutations aiming at constructing l-lysine-overproducing B. methanolicus strains that do not simultaneously co-secrete l-glutamate. Pyruvate carboxylase (PC) activity has been reported to be one major bottleneck for high l-lysine production in C. glutamicum to supply OAA (Peters-Wendish et al. 2001; Ohnishi et al. 2002). PC activity was significantly elevated in the l-lysine-overproducing B. methanolicus mutant NOA2#13A52-8A66 compared to in the wild-type strain. Both the wild type B. methanolicus and l-lysine-overproducing mutant NOA2#13A52-8A66 have high intracellular levels of OAA, about 450 mM, indicating a high activity of PC in this bacterium (Brautaset et al. 2003). Also, genetic analysis of mutant NOA2#13A52-8A66 has demonstrated that it has no mutations in the citY gene. Thus, it seems plausible to suggest that metabolic engineering to further improve l-lysine production of B. methanolicus should focus on deregulating key enzymes in the l-lysine biosynthetic pathway downstream of PC and OAA.
The l-lysine biosynthetic pathway from aspartate
Deregulation of aspartate kinase (AK) has been the most important step in the development of l-lysine-overproducing strains (Pfefferle et al. 2003), and screening for AEC resistance has led to the development of AK-deregulated and l-lysine-overproducing mutants of B. subtilis (Chaudhuri et al. 1983). C. glutamicum has one AK while bacilli typically have three AK isoenzymes that are regulated in a distinct manner (Paulus 1993; Belitsky 2002 and references therein). The B. methanolicus genes lysC (encoding AKII) and lysA (encoding DAP decarboxylase), representing the first and the last step in the l-lysine biosynthetic pathway (Fig. 5), respectively, have been cloned and characterized (Schendel and Flickinger 1992; Mills and Flickinger 1993). Biochemical characterizations of the lysA gene product confirmed that the protein is also inhibited by l-lysine (Mills and Flickinger 1993), and both lysC and lysA may represent interesting targets for genetic manipulations to improve l-lysine production by B. methanolicus.
Establishment of a gene delivery system and its application to improve methanol consumption in B. methanolicus
The characterization of the restriction modification system (Cue et al. 1996) and the reporting of a gene delivery method by using protoplast transformation (Cue et al. 1997) were the first reports of molecular biology techniques for recombinant manipulation of B. methanolicus. However, due to incompatibility problems with the constructed shuttle vectors and very poor transformation efficiencies (Brautaset et al. 2004), no recombinant B. methanolicus strains were published using these methodologies. The characterization of the natural B. methanolicus MGA3 plasmid pBM19 (see above) gave some biological explanations to these problems. The Escherichia coli–B. methanolicus shuttle vector pDQ508 constructed by Cue et al. (1997) contains replication elements of pBM19, and selecting for pDQ508 caused loss of pBM19 and concomitant lost ability of recombinant cells to grow on methanol (Brautaset et al. 2004). However, a new electroporation method for gene delivery by using the E. coli–B. subtilis shuttle plasmid pHP13 was recently established by Jakobsen et al. (2006). This method involves selection in liquid medium and implements an adaptation strategy when transferring recombinant cells from rich to defined methanol media, to avoid lethal formaldehyde accumulation and death of recombinant cells. By using this method, recombinant B. methanolicus strains that retain the ability to grow on methanol can be constructed (Jakobsen et al. 2006). This has resulted in new possibilities to use metabolic engineering strategies to improve production properties of this organism.
Improved methanol consumption and methanol tolerance level of B. methanolicus by metabolic engineering
The unique genetic organization of B. methanolicus RuMP pathway genes (see above) may lead to new possibilities to improve the methanol assimilation rate of this bacterium by genetic engineering. To achieve high amino acid production, an efficient assimilation of the methanol in favor of wasteful dissimilation into CO2 is desirable (see Fig. 3). In contrast to the pBM19-borne RuMP genes, which all appear in multiple copies, the operon encoding hps and phi (Fig. 2d) is present in a single copy in the cells (Brautaset et al. 2004). Therefore, in an attempt to increase the RuMP pathway activity in the cells, this chromosomal operon was cloned on the medium copy number vector pHP13 and transformed into B. methanolicus wild-type MGA3 by electroporation (see above). The resulting recombinant strain displayed about threefold increased coupled HPS + PHI activity and had a significantly improved growth rate on a range of high methanol concentrations (up to 1,440 mM) compared to the wild-type (Jakobsen et al. 2006). The recombinant cells also displayed significantly higher tolerance level to high methanol concentrations in the growth media before cell growth was impaired. Together, these data demonstrate that the coupled HPS + PHI activity of the RuMP pathway (Fig. 2a) plays an important role in regulating the methanol assimilation rate and the methanol tolerance level in the cells (Jakobsen et al. 2006).
Conclusions and perspectives
During the recent years considerable scientific progress has resulted in an increased understanding of the unique genetics and physiology of thermotolerant B. methanolicus. In particular, the metabolic routes for methanol consumption are well-characterized and this knowledge has led to better cultivation strategies, new and useful insight into the mechanisms for formaldehyde and methanol sensitivity, and has also been valuable for the development of recombinant methodologies for B. methanolicus strains. The usefulness of this acquired biological knowledge and new technology has been demonstrated by the construction of recombinant B. methanolicus mutants with improved methanol consumption rate and increased formaldehyde tolerance level. Key genes involved in both l-glutamate and l-lysine production have been cloned and characterized, and the construction and characterization of classical mutants overproducing l-lysine has gained important insight into the intermediary methanol metabolism related to high amino acid production of B. methanolicus. Such mutants should also represent valuable sources useful for inverse metabolic engineering strategies involving mapping of beneficial mutations resulting in improved amino acid production. By using the genetic tools, mutant genes can be reintroduced into a defined genetic background to obtain recombinant strains with improved amino acid production. Such methods have been successfully used to generate effective l-lysine overproducers of C. glutamicum (Ohnishi et al. 2002).
As we see it, the major limitations for constructing industrial competitive producer strains of B. methanolicus at present are the lack of a genome sequence and also a genetic toolbox that should be further expanded. The total number of B. methanolicus genes sequenced is so far relatively small (De Vries et al. 1992; Schendel and Flickinger 1992; Mills and Flickinger 1993; Brautaset et al. 2003, 2004). A genome sequence is required for global analyses (transcriptome, metabolome, proteome) of methanol metabolism in B. methanolicus and its amino acid overproducing mutants and for more efficient targeting of key enzymes important for high amino acid production. By also increasing the genetic toolbox of this bacterium, e.g., to include suicide vectors for homologous recombination, adjustable expression systems, and obtain higher transformation efficiencies, the potential for manipulating genes encoding key enzymes involved in conversion of methanol into l-lysine and l-glutamate could be fully explored. The documented high methanol consumption rate and the high capability of B. methanolicus strains to overproduce both l-glutamate and l-lysine from methanol at elevated temperature should make the new era of using metabolic engineering to improve these properties exciting from a scientific and from an applied point of view.
References
Al-Awadhi NT, Egli T, Hamer G (1988) Growth-characteristics of a thermotolerant methylotrophic Bacillus sp (NCIB-12522) in batch culture. Appl Microbiol Biotechnol 29:485–493
Al-Awadhi N, Egli T, Hamer G, Wehrli E (1989) Thermotolerant and thermophilic solvent-utilizing methylotrophic aerobic bacteria. Syst Appl Microbiol 11:207–216
Anthony C (1982) The biochemistry of methylotrophs. Academic Press, New York
Anthony C (1991) Assimilation of carbon by methylotrophs. In: Goldberg I, Rokem JS (eds) Biology of methylotrophs. Butterworth-Heinemann, Boston
Arfman N, Watling EM, Clement W, van Oosterwijk RJ, de Vries GE, Harder W, Attwood MM, Dijkhuizen L (1989) Methanol metabolism in thermotolerant methylotrophic Bacillus strains involving a novel catabolic NAD-dependent methanol dehydrogenase as a key enzyme. Arch Microbiol 152:280–288
Arfman N, Dijkhuizen L, Kirchhof G, Ludwig W, Schleifer K-H, Bulygina ES, Chumakov KM, Govorukhina NI, Trotsenko YA, White D, Sharp RJ (1992a) Bacillus methanolicus sp. nov., a new species of thermotolerant, methanol-utilizing, endospore-forming bacteria. Int J Sys Bacteriol 42:439–445
Arfman N, de Vries KJ, Moezelaar HR, Attwood MM, Robinson GK, van Geel M, Dijkhuizen L (1992b) Environmental regulation of alcohol metabolism in thermotolerant methylotrophic Bacillus strains. Arch Microbiol 157:272–278
Arfman N, Hektor HJ, Bystrykh LV, Govorukhina NI, Dijkhuizen L, Frank J (1997) Properties of a NAD(H)-containing methanol dehydrogenase and its activator protein from Bacillus methanolicus. Eur J Biochem 244:426–433
Belitsky BR (2002) Biosynthesis of amino acids of the glutamate and aspartate families, alanine and polyamines. In: Sonenshein AL, Hoch JA, Losick R (eds) Bacillus subtilis and its closest relatives: from genes to cells. ASM Press, Washington DC, pp 203–232
Berges DA, DeWolf WE, Dunn GL, Newman DJ, Schmidt SJ, Taggart JJ, Gilvarg C (1986) Studies on the active sites of succinyl-CoA: tetrahydrodipicolinate N-succinyltransferase. J Biol Chem 261:6160–6167
Brautaset T, Williams MD, Dillingham RD, Kaufmann C, Bennaars A, Crabbe E, Flickinger MC (2003) Role of the Bacillus methanolicus citrate synthase II gene citY in regulating the secretion of glutamate in l-lysine-secreting mutants. Appl Environ Microbiol 69:3986–3995
Brautaset T, Jakobsen ØM, Flickinger MC, Valla S, Ellingsen TE (2004) Plasmid-dependent methylotrophy in thermotolerant Bacillus methanolicus. J Bacteriol 186:1229–1238
Brooke AG, Watling EM, Attwood MM, Tempest DW (1989) Environmental control of metabolic fluxes in thermotolerant methylotrophic Bacillus strains. Arch Microbiol 151:268–273
Chaudhuri A, Mishra AK, Nanda G (1983) Lysine excretion by S-(2-aminoethyl)l-cysteine resistant mutants of Bacillus subtilis. Acta Microbiol Pol 32:37–45
Cue D, Lam H, Hanson RS, Flickinger MC (1996) Characterization of a restriction-modification system of the thermotolerant methylotroph Bacillus methanolicus. Appl Environ Microbiol 62:1107–1111
Cue D, Lam H, Dillingham RL, Hanson RS, Flickinger MC (1997) Genetic manipulation of Bacillus methanolicus, a gram positive, thermotolerant methylotroph. Appl Environ Microbiol 63:1406–1420
De Vries GE, Kües U, Stahl U (1990) Physiology and genetics of methylotrophic bacteria. FEMS Microbiol Rev 6:57–102
De Vries GE, Arfman N, Terpstra P, Dijkhuizen L (1992) Cloning, expression, and sequence analysis of the Bacillus methanolicus C1 methanol dehydrogenase gene. J Bacteriol 174:5346–5353
Dijkhuizen L, Arfman N, Attwood MM, Brooke AG, Harder W, Watling EM (1988) Isolation and initial characterization of thermotolerant methylotrophic Bacillus strains. FEMS Microbiol Lett 52:209–214
Dijkhuizen L, Levering PR, De Vries GE (1992) The physiology and biochemistry of aerobic methanol-utilizing Gram-negative and Gram-positive bacteria. In: Murrell JC, Dalton H (eds) Methane and methanol utilizers. Plenum, New York
Eggeling L, Bott M (2005) Handbook of Corynebacterium glutamicum. Taylor & Francis, Boca Raton
Faurie R, Thommel J (eds) (2003) Microbial production of l-amino acids. Advances in biochemical engineering, biotechnology vol. 79. Springer, Berlin Heidelberg New York
Fisher SH (1999) Regulation of nitrogen metabolism in Bacillus subtilis: vive la difference! Mol Microbiol 32:223–232
Gunji Y, Yasueda H (2006) Enhancement of L-lysine production in methylotroph Methylophilus methylotrophus by introducing a mutant lysE exporter. J Biotechnol June 15 127(1):1–13
Hagishita T, Yoshida T, Izumi Y, Mitsunaga T (1996) Efficient l-serine production from methanol and glycine by resting cells of Methylobacterium sp. Strain MN43. Biosci Biotechnol Biochem 60:1604–1610
Hanson RS, Dillingham R, Olson P, Lee GH, Cue D, Schendel FJ, Bremmon C, Flickinger MC (1996) Production of l-lysine and some other amino acids by mutants of Bacillus methanolicus. In: Lidstrom ME, Tabita FR (eds) Microbial growth on C1 compounds. Kluwer, The Netherlands, pp 227–236
Hanson RS, Flickinger MC, Olson P, Hur W, Al-Tahoo N, Bremmon C (2000) Production of glutamate by wild type Bacillus methanolicus US Patent 6,110,713
Hektor HJ, Kloosterman H, Dijkhuizen L (2002) Identification of a magnesium-dependent NAD(P)(H)-binding domain of the nicotinoprotein methanol dehydrogenase from Bacillus methanolicus. J Biol Chem 277:46966–46973
Ikeda M (2003) Amino acid production processes. Adv Biochem Eng Biotechnol 79:1–35
Ikeda M, Nakagawa S (2003) The Corynebacterium glutamicum genome: Features and impacts on the biotechnological processes. Appl Microbiol Biotechnol 62:99–109
Izumi Y, Yoshida T, Miyazaki SS, Mitsunaga T, Oshiro T, Shimao M, Miyata A, Tanabe T (1993) l-serine production by a methylotroph and its related enzymes. Appl Microbiol Biotechnol 39:427–432
Jakobsen ØM, Benichou A, Flickinger MC, Valla S, Ellingsen TE, Brautaset T (2006) Upregulated transcription of plasmid and chromosomal ribulose monophosphate pathway genes is critical for methanol assimilation and methanol tolerance in the methylotrophic bacterium Bacillus methanolicus. J Bacteriol 188:3063–3072
Kalinowski J, Bathe B, Bartels D, Bischoff N, Bott M, Burkovski A, Dusch N, Eggeling L, Eikmanns BJ, Gaigalat L, Goesmann A, Hartmann M, Huthmacher K, Kramer R, Linke B, Mchardy AC, Meyer F, Mockel B, Pfefferle W, Puhler A, Rey DA, Ruckert C, Rupp O, Sahm H, Wendisch VF, Wiegrabe I, Tauch A (2003) The complete Corynebacterium glutamicum ATCC 13032 genome sequence and its impact on the production of l-aspartate-derived amino acids and vitamins. J Biotechnol 104:5–25
Kato N, Yurimoto H, Thauer RK (2006) The physiological role of the ribulose monophosphate pathway in bacteria and archaea. Biosci Biotechnol Biochem 70:10–21
Kindler SH, Gilvarg C (1960) N-succinyl-l-α,ɛ-diaminopimelate acid deacylase. J Biol Chem 235:3532–3535
Kloosterman H, Vrijbloed JW, Dijkhuizen L (2002) Molecular, biochemical, and functional characterization of a nudix hydrolase protein that stimulates the activity of nicotinoprotein alcohol dehydrogenase. J Biol Chem 277:34785–34792
Komives CF, Cheung LY-Y, Pluschkell SB, Flickinger MC (2005) Growth of Bacillus methanolicus in seawater-based media. J Ind Microbiol Biotechnol 32:61–66
Kono K, Oki T, Kitai A, Ozaki A (1972) Process for producing l-glutamic acid by fermentation. US Patent 3663370
Large PJ, Bamforth CW (1988) Methylotrophy and biotechnology. Wiley, New York
Lee GH, Hur W, Bremmon CE, Flickinger MC (1996) Lysine production from methanol at 50°C using Bacillus methanolicus: modeling volume control, lysine concentration and productivity using a three-phase continuous simulation. Biotechnol Bioeng 49:639–653
Linton JD, Niekus HGD (1987) The potential of one-carbon compounds as fermentation feedstocks. Antonie Van Leeuwenhoek 53:55–63
Marx A, Wendisch VF, Buchholz S (2006) Protein line and amino acid-based product family trees. In: Kamm B, Gruber PR, Kamm M (eds) Biorefineries—industrial processes and products. Status quo and future directions. Wiley-VCH, Weinheim, pp 201–216
Mills DA, Flickinger MC (1993) Cloning and sequence analysis of the meso-diaminopimelate decarboxylase gene from Bacillus methanolicus MGA3 and comparison to other decarboxylase genes. Appl Environ Microbiol 59:2927–2937
Motoyama H, Anazawa H, Katsumata R, Araki K, Teshiba S (1993) Amino acid production from methanol by Methylobacillus glycogens mutants: isolation of l-glutamic acid hyperproducing mutants from M. glycogens strains, and derivation of L-threonine and l-lysine-producing mutants from them. Biotechnol Biochem 57:82–87
Motoyama H, Yano H, Ishino S, Anazawa H, Teshiba S (1994) Effects of the amplification of the genes coding for the l-threonine biosynthetic enzymes of the l-threonine production from methanol by a gram negative obligate methylotroph, Methylobacillus glycogens. Appl Microbiol Biotechnol 42:67–72
Motoyama H, Yano H, Terasaki Y, Anazawa H (2001) Overproduction of l-lysine from methanol by Methylobacillus glycogens derivatives carrying a plasmid with a mutated dapA gene. Appl Environ Microbiol 67:3064–3070
Nakayama K, Kobata M, Tanaka Y, Nomura T, Katsumata R (1976) Process for the production of l-glutamic acid. US Patent 3939042
Nishio Y, Nakamura Y, Kawarabayasi Y, Usuda Y, Kimura E, Sugimoto S, Matsui K, Yamagishi A, Kikuchi H, Ikeo K, Gojobori T (2003) Comparative complete genome sequence analysis of the amino acid replacements responsible for the thermostability of Corynebacterium efficiens. Genome Res 13:1572–1579
Olah GA, Goeppert A, Prakash GKS (2006) Beyond oil and gas: the methanol economy. Wiley-VCH, Weinheim
Ohnishi J, Mitsuhashi S, Hayashi M, Ando S, Yokoi H, Ochiai K, Ikeda M (2002) A novel methodology employing Corynebacterium glutamicum genome information to generate a new l-lysine-producing mutant. Appl Microbiol Biotechnol 58:217–223
Paulus H (1993) Biosynthesis of the aspartate family of amino acids. In: Sonenshein AL (ed) Bacillus subtilis and other Gram-positive bacteria: biochemistry, physiology, and molecular genetics. American Society for Microbiology, Washington DC, pp 237–267
Peters-Wendish PG, Schiel B, Wendisch VF, Katsoulidis E, Möckel B, Sahm H, Eikmanns BJ (2001) Pyruvate carboxylase is a major bottleneck for glutamate and lysine production by Corynebacterium glutamicum. J Mol Microbiol Biotechnol 3:295–300
Pfefferle W, Mockel B, Bathe B, Marx A (2003) Biotechnological manufacture of lysine. Adv Biochem Eng Biotechnol 79:59–112
Pluschkell SB, Flickinger MC (2002) Dissimilation of [13C]methanol by continuous cultures of Bacillus methanolicus MGA3 at 50°C studied by 13C NMR and isotope-ratio mass spectrometry. Microbiology 148:3223–3233
Schendel FJ, Flickinger MC (1992) Cloning and nucleotide sequence of the gene coding for aspartokinase II from a thermophilic methylotrophic Bacillus sp. Appl Environ Microbiol 58:2806–2814
Schendel FJ, Bremmon CE, Flickinger MC, Guettler M, Hanson RS (1990) l-Lysine production at 50°C by mutants of a newly isolated and characterized methylotrophic Bacillus sp. Appl Environ Microbiol 56:963–970
Schendel FJ, Dillingham R, Hanson RS, Sano K, Matsui K (2000) Production of glutamate using wild-type Bacillus methanolicus. US patent 6083728
Schrumpf B, Schwarzer A, Kalinowski J, Puhler A, Eggeling L, Sahm H (1991) A functionally split pathway for Lysine synthesis in Corynebacterium glutamicum. J Bacteriol 173:4510–4516
Shishkina VN, Trotsenko YA (1982) Multiple enzymic lesions in obligate methanotrophic bacteria. FEMS Microbiol Lett 13:237–242
Snedecor B, Cooney CL (1974) Thermophilic mixed cultures of bacteria using methanol for growth. Appl Microbiol 27:1112–1117
Sonntag K, Eggeling L, De Graaf AA, Sahm H (1993) Flux partitioning in the split pathway of lysine in Corynebacterium glutamicum. Eur J Biochem 213:1325–1331
Sundharadas G, Gilvarg C (1967) Biosynthesis of 2,6-diaminopimelate acid in Bacillus megaterium. J Biol Chem 242:2988–3983
Tosaka O, Takinami K (1978) Pathway and regulation of lysine biosynthesis in Brevibacterium lactofermentum. Agric Biol Chem 42:95–100
Tsujimoto N, Gunji Y, Ogawa-Miyata Y, Shimaoka M, Yasueda H (2006) l-lysine biosynthetic pathway of Methylophilus methylotrophus and construction of an L-lysine producer. J Biotechnol 124:327–337
Wehrmann A, Phillipp P, Sahm H, Eggeling L (1998) Different modes of diaminopimelate synthesis and their role in cell wall integrity: a study with Corynebacterium glutamicum. J Bacteriol 180:3159–3165
Weinberger S, Gilvarg C (1970) Bacterial distribution of the use of succinyl and acetyl blocking groups in diaminopimelate acid biosynthesis. J Bacteriol 101:323–324
White PJ (1983) The essential role of diaminopimelate dehydrogenase in the biosynthesis of lysine by Bacillus sphaericus. J Gen Microbiol 129:739–749
Zatman LJ (1981) A search for patterns in methylotrophic pathways. In: Dalton H (ed) Microbial growth on C1 compounds: proceedings of the Third International Symposium held in Sheffield, UK, 12–16 August 1980. Heyden London, pp 42–54
Acknowledgements
The writing of this mini-review was supported by a grant from the Research Council of Norway with support from the University of Minnesota BioTechnology Institute.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Brautaset, T., Jakobsen, Ø.M., Josefsen, K.D. et al. Bacillus methanolicus: a candidate for industrial production of amino acids from methanol at 50°C. Appl Microbiol Biotechnol 74, 22–34 (2007). https://doi.org/10.1007/s00253-006-0757-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00253-006-0757-z