
Abstract
Evolutionary models of molecular structures must incorporate molecular information at different levels of structural complexity and must be phrased within a phylogenetic perspective. In this regard, phylogenetic trees of substructures that are reconstructed from molecular features that contribute to order and thermodynamic stability show that a gradual model of evolution of 5S rRNA structure is more parsimonious than models that invoke large segmental duplications of the molecule. The search for trees of substructures that are most parsimonious, by their very nature, defines an objective strategy to select models of molecular change that best fit structural data. When combined with additional data, such as the age of protein domains that interact with RNA substructures, these trees can be used to falsify unlikely hypotheses.
Avoid common mistakes on your manuscript.
In recent correspondence, Di Giulio (2010) proposes a model for the evolution of the 5S rRNA molecule that involves an inverse duplication of a primordial molecule. He claims that this model, which follows a similar model for the evolution of tRNA (Di Giulio 1992), is more parsimonious than a gradual evolutionary scenario for the history of the structure of 5S rRNA that we recently proposed (Sun and Caetano-Anollés 2009). The model of Di Giulio is appealing in that it splits the three helical branches of the molecule (domains α, β, and γ) into ancient and derived halves, each having only one domain (ancestral β and derived γ). The duplication of the ancestor molecule (containing domain β) and later the annealing of duplicates by hydrogen bonds are responsible for the assembly of the third domain (domain α), which is therefore derived. However, parsimony considerations and phylogenetic and topological analyses of the structure and sequence of RNA and associated protein molecules do not support the duplication model.
-
(1)
Phylogenetic constraint analysis of the structure of 5S rRNA shows the duplication model is less parsimonious than the gradual evolutionary model (Fig. 1). Using the phylogenetic method of Sun and Caetano-Anollés (2009), 492–565 and 370 additional steps of change are needed to reconstruct trees of substructures that match the inverse duplication model, using either domains or stems as atomized substructures, respectively. This represents a substantial departure from the most parsimonious scenario.
Fig. 1 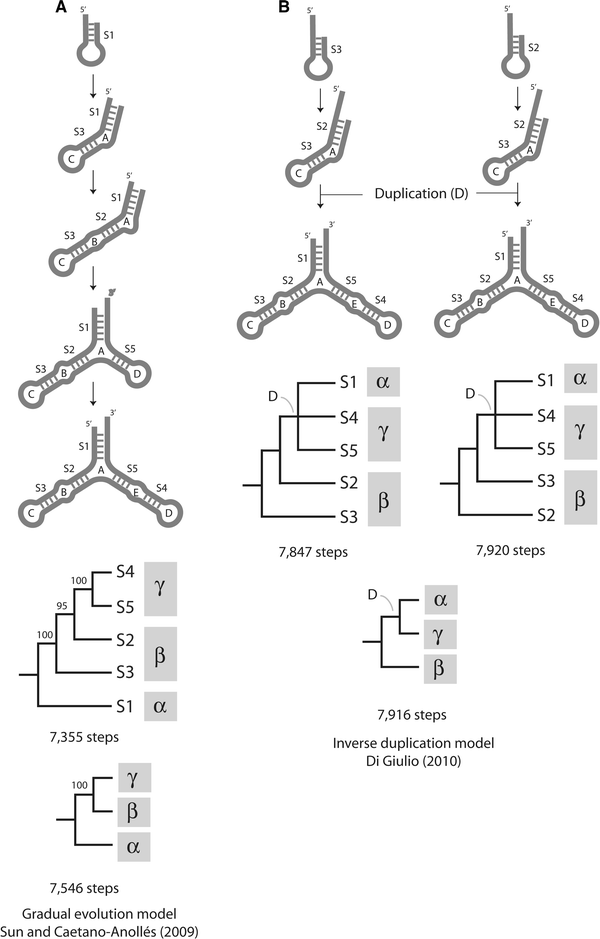
Models of 5S rRNA evolution and trees of substructures. a Gradual evolution model derived directly from the most parsimonious tree of molecular substructures describing the evolution of helical stems (7,355 steps; CI = 0.869, RI = 0.570; RC = 0.495) and corresponding tree of molecular domains (7,546; CI = 0.847; RI = 0.485; RC = 0.410). b Inverse duplication model and corresponding trees of substructures obtained by constraining the topology of trees of stems (7,847–7,920 steps; CI = 0.807–0.814; RI = 0.318–0.351; RC = 0.257–0.286) or trees of domains (7,916 steps; CI = 0.807; RI = 0.320; RC = 0.258) to reconstructions that fit the model. Methods, data matrices, and nomenclature for loops and stems are those described in Sun and Caetano-Anollés (2009). Nodes of constrained trees have bootstrap support values of 100%. CI consistency index, RI retention index, RC rescaled consistency index
-
(2)
The secondary structure of 5S rRNA molecules is not symmetric in that the length of the different helices (S1–S5) is variable. Analysis of the sequence of consensus 5S rRNA molecules (Barciszewska et al. 2001) does not reveal signatures of complementarity between stems S2 and S5 and stems S3 and S4, a feature that is prerequisite for the validity of the duplication model. Furthermore, Di Giulio (2010) has not presented evidence either of cryptic patterns suggestive of ancestral complementarity, such as those in the classical and operational genetic codes in tRNA (Rodin et al. 1993, 1996).
-
(3)
Phylogenomic analysis of protein domain structure suggests that the age of interactions between 5S rRNA substructures is tightly correlated with the age of associated ribosomal proteins (Sun and Caetano-Anollés 2009), with the most ancient protein domain (protein L21) interacting with domain α, the most ancient 5S rRNA domain according to the gradual evolutionary model. These data contradict the derived nature of domain α, a necessary consequence of the duplication model.
-
(4)
Crystal structures reveal that nearly all of rRNA is helical or approximately helical. In fact, the RNA of the ribosomal ensemble can be effectively considered a three-dimensional (3D) arrangement of ~200 helical elements (Wimberly et al. 2000; Yusupov et al. 2001). With few exceptions (Agmon 2009), disparate helical elements make up crucial functional centers, indicating that the aggrandizement of ribosomal structure is mediated by accretion of helical structure within the context of high order 3D structure (e.g., A-motifs; Bokov and Steinberg 2009). 5S rRNA, the minor subunit, does not escape to this rule. The stems of 5S rRNA interact with different ribosomal proteins. For example, the S1–S5 stems in archaeal molecules interact more or less separately with five ribosomal proteins (Sun and Caetano-Anollés 2009). The functional and structural constraints imposed by these tight interactions make it unlikely that evolutionary change will split 5S rRNA domains.

Di Giulio in his analysis uses economy in the number of steps needed to generate an extant molecule to select for models of molecular evolution. However, this can be deceptive, especially because steps are of different types and may merit differential weighting or other considerations, and because optimality must be exhaustively explored within a space of possible models. We know today that mutational change in biology can be in some cases gradual (e.g., nucleotide substitutions and small indels) and in others punctuated (e.g. entire or segmental genome duplications), and that these modes are not mutually exclusive. There is no reason to believe that gradual and punctuated processes did not exist during the origin of ancient RNA and proteins molecules. Our cladistic approach defines change at the structural level using considerations in thermodynamics and molecular mechanics, with central assumptions also supported by phylogenetics and cosmology (recently reviewed in Sun et al. 2010). In our analysis, each change at the nucleotide level can contribute or interfere with the stability and order of the evolving molecule. Fortunately, the most parsimonious trees of molecular substructures capture these patterns of change and are therefore appropriate tools to test alternative evolutionary scenarios, refuting those that are not supported by structural data. In fact, trees of substructures that are most parsimonious by their very nature define an objective strategy to select the model of molecular change that best fits the data. Given these results, we contend that a large duplication cannot explain the evolutionary growth of the 5S rRNA molecule. This does not mean that smaller duplications (smaller than or equal to the helical stems we have analyzed) did not curb the structure of these molecules.
References
Agmon I (2009) The dimeric proto-ribosome: structural details and possible implications on the origin of life. Int J Mol Sci 10:2921–2934
Barciszewska MZ, Szymanski M, Erdmann VA, Barciszewski J (2001) Structure and functions of 5S rRNA. Acta Biochim Pol 48:191–198
Bokov K, Steinberg SV (2009) A hierarchical model for the evolution of 23S ribosomal RNA. Nature 457:977–980
Di Giulio M (1992) On the origin of the transfer RNA molecule. J Theor Biol 159:199–214
Di Giulio M (2010) A model of the origin of the 5S ribosomal RNA molecule. J Mol Evol (in press)
Rodin S, Ohno S, Rodin A (1993) Transfer RNAs with complementary anticodons: could they reflect early evolution of discriminative genetic code adaptors? Proc Natl Acad Sci USA 90:4723–4727
Rodin S, Rodin A, Ohno S (1996) The presence of codon-anticodon pairs in the acceptor stem of tRNAs. Proc Natl Acad Sci USA 93:4537–4542
Sun F-J, Caetano-Anollés G (2009) The evolutionary history of the structure of 5S ribosomal RNA. J Mol Evol 69:430–443
Sun F-J, Harish A, Caetano-Anollés G (2010) Phylogenetic utility of RNA structure: Evolution’s arrow and emergence of early biochemistry and diversified life. In: Caetano-Anolles G (ed) Evolutionary genomics and systems biology. Wiley & Sons, Inc., Hoboken, pp 329–360
Wimberly BT, Brodersen DE, Clemons WM, Morgan-Warren RJ, Carter AP, Vonrhein C, Hartsch T, Ramakrishnan V (2000) Structure of the 30S ribosomal subunit. Nature 407:327–339
Yusupov MM, Yusupova GZ, Baucom A, Lieberman K, Earnest TN, Cate JHD, Noller HF (2001) Crystal structure of the ribosome at 5.5 angstrom resolution. Science 292:883–896
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sun, FJ., Caetano-Anollés, G. The Origin of Modern 5S rRNA: A Case of Relating Models of Structural History to Phylogenetic Data. J Mol Evol 71, 3–5 (2010). https://doi.org/10.1007/s00239-010-9359-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00239-010-9359-6