
Abstract
Vestibular perceptual thresholds provide insights into sensory function and have shown clinical and functional relevance. However, specific sensory contributions to tilt and rotation thresholds have been incompletely characterized. To address this limitation, tilt thresholds (i.e., rotations about earth-horizontal axes) were quantified to assess canal-otolith integration, and rotation thresholds (i.e., rotations about earth-vertical axes) were quantified to assess perception mediated predominantly by the canals. To determine the maximal extent to which non-vestibular sensory cues (e.g., tactile) can contribute to tilt and rotation thresholds, we tested two patients with completely absent vestibular function and compared their data to those obtained from two separate cohorts of young (≤ 40 years), healthy adults. As one primary finding, thresholds for all motions were elevated by approximately 2–35 times in the absence of vestibular function, thus, confirming predominant vestibular contributions to both rotation and tilt self-motion perception. For patients without vestibular function, rotation thresholds showed larger increases relative to healthy adults than tilt thresholds. This suggests that increased extra-vestibular (e.g., tactile or interoceptive) sensory cues may contribute more to the perception of tilt than rotation. In addition, an impact of stimulus frequency was noted, suggesting increased vestibular contributions relative to other sensory systems can be targeted on the basis of stimulus frequency.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Vestibular perceptual thresholds, quantified using whole-body direction recognition tasks, represent the smallest passive self-motion cue that an individual can reliably perceive (Merfeld 2011). Vestibular thresholds have potential for future clinical applications (Kobel et al. 2021b) as they have been shown to be sensitive to vestibular disorders (Agrawal et al. 2013; Bremova et al. 2016; King et al. 2019) and some threshold measures correlate with balance performance metrics that quantify amount of sway during quiet stance balance tasks (Karmali et al. 2021; Wagner et al. 2021). Past research quantifying vestibular contributions to self-motion perception have assessed vestibular thresholds in the complete absence of vestibular function in order to quantify the maximal extent by which non-vestibular cues (e.g., tactile, somatic graviception) contribute to self-motion perception (Valko et al. 2012). While these initial efforts were able to identify predominant vestibular contributions to self-motion perception, as perceptual thresholds were elevated by ~ 1.5–85 times relative to healthy controls, the experimental design focused on determining the impact of stimulus frequency (i.e., duration of motion stimuli) for only a limited set of tilt and rotation trajectories (Valko et al. 2012). Subsequent investigations have since focused on comprehensively quantifying the factors influencing tilt and rotation thresholds in healthy populations of young adults (Wagner et al. 2022), however, without assessing measures in the complete absence of vestibular function, the contributions of vestibular and non-vestibular contributions to motion perception cannot be fully understood.
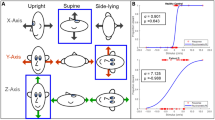
The semicircular canals are sensitive to angular velocity (Fernandez and Goldberg 1971) and due to their anatomic orientations, can be stimulated by rotations about earth-horizontal or earth-vertical axes. Even though both motions include rotation, for rhetoric simplicity, rotations about earth-horizontal axes will be referred to as “tilts” and rotations about earth-vertical axes will be referred to as “rotations”. Using this nomenclature, rotations primarily stimulate the respective canals in the plane of rotation that are sensitive to the angular velocity being applied. In contrast, tilts provide concurrent canal and otolith stimulation due to: (a) the angular velocity that stimulates the canals and (b) the resulting re-orientation of the head relative to gravity (“tilt”), which stimulates the otoliths. Figure 1 depicts roll tilt and roll rotation stimuli.

Depiction of roll tilt (A) and roll rotation (B) experimental paradigms. Axis of rotation is denoted by dashed arrows while rotational stimuli are denoted by solid arrows. Roll tilt stimuli are rotations about an earth-horizontal axis assessed while upright. Axis of rotation for roll tilt is along a naso-occipital axis and is not pictured for illustrative simplicity. Roll rotation stimuli are rotations about an earth-vertical axis assessed while supine
Past efforts quantifying vestibular contributions to perceptual threshold tasks have not assessed potential impact of axis of rotation (i.e., tilts in comparison to rotations in the same plane of motion). However, in the complete absence of vestibular function, yaw rotation thresholds were elevated by ~ 5 to 15 times relative to the normative data, while roll tilt thresholds were only elevated by ~ 1.5–3 times (Valko et al. 2012). Since (1) yaw rotations and roll tilts have different axes of rotation, earth-vertical and earth-horizontal, respectively, and (2) these patients had no residual vestibular function (i.e., canals or otoliths), this suggests a greater role for non-vestibular cues (e.g., tactile/interoceptive) for tilts relative to rotations.
In healthy young adults (≤ 40 years old), our past study (Wagner et al. 2022) quantified the difference between tilts and rotations (i.e., impact of axis of rotation) across multiple frequencies (0.2–2 Hz) for vestibular perceptual thresholds. Overall, we identified an influence of axis of rotation (i.e., difference between tilts and rotation) that was dependent on frequency for roll and pitch motions as well as for and trajectories designed to maximally stimulate the right-anterior and left-posterior (RALP) and left-anterior and right-posterior (LARP) vertical canal pairs. For higher frequencies (i.e., at 2 Hz), tilt thresholds were indistinguishable from rotation thresholds, suggesting that high-frequency tilt responses are mediated primarily by the vertical canals (Wagner et al. 2022)—representing an angular velocity response. At lower frequencies (i.e., below ~ 1 Hz), tilt thresholds were significantly lower than rotation thresholds (Lim et al. 2017; Suri and Clark 2020; Wagner et al. 2022), presumably reflecting beneficial contributions from the added otolith stimulation. However, whether non-vestibular cues might help explain these frequency effects has yet to be fully investigated.
Particularly, an impact of body orientation may play a role as when assessing the vertical canals, tilt and rotation thresholds are assessed using two different body orientations (upright and supine, respectively, as depicted in Fig. 1). Past research has suggested that perceptual sensitivity may be modulated by absolute body orientation (Hummel et al. 2016; MacNeilage et al. 2010) with decreases in perceptual sensitivity (i.e., increased thresholds) noted when perception is measured in non-upright body orientations, including supine (Hummel et al. 2016; MacNeilage et al. 2010). Thus, changes in body orientation could contribute to the differences previously identified between rotation and tilt thresholds.
Finally, a generalized increase in sensitivity for the vertical canals in comparison to the horizontal canals has been proposed based upon recordings of vestibular afferent neurons (Yang and Hullar 2007). However, direct comparisons between vertical and horizontal canal perceptual metrics have yet to be completed as past studies have historically focused on either assessing the vertical canals (Wagner et al. 2022) or horizontal canals (Grabherr et al. 2008) in isolation. As high-frequency RALP tilt and LARP tilt thresholds reflect predominantly vertical canal function, comparison of these thresholds to yaw rotation thresholds would allow insights into the relative sensitivity to angular velocity of the horizontal and vertical canals. Additionally, as each of these thresholds are measured while upright—horizontal canals sensing rotation about an earth-vertical axis (i.e., rotations) and vertical canals sensing rotation about an earth-horizontal axis (i.e., tilts)—quantification of these measures in the same subjects will allow insights into potential differences in sensitivity between tilts and rotations (i.e., changes in axis of rotation) without confounding the effects of body orientation.
Thus, in order to unravel the relative contributions of the non-vestibular factors that impact tilt and rotation motion perception, this project compared a wide-ranging set of vestibular perceptual thresholds between two patients with a complete absence of vestibular function and two cohorts of healthy controls. Roll rotation and roll tilt thresholds were assessed at both low (i.e., 0.5 Hz) and high (i.e., 2 Hz) frequencies to assess changes in roll perceptual sensitivity modulated on the basis of frequency and axis of rotation (i.e., tilts vs. rotations). Additionally, angular velocity thresholds in three semicircular canal planes were assessed to determine potential differences in perceptual sensitivity driven by differences in horizontal or vertical canal stimulation and differences between tilts and rotations (i.e., changes in axis of rotation). Such efforts aim not only to investigate the vestibular contribution to motion perception, but also to validate threshold measures as an assessment for peripheral end-organ function.
Methods
Participants
Two participants with neuro-fibromatosis type II (NF-2) and complete bilateral surgical ablation of the labyrinths secondary to vestibular schwannoma removal completed vestibular perceptual threshold assessments. Our past research suggests that patients identified with severe bilateral vestibular hypofunction of the horizontal semicircular canals on the basis of traditional clinical testing (e.g., calorics, rotational testing) did not exhibit uniform degradation in function across all peripheral end-organs as evidenced by superior-inferior z-axis translation and roll tilt thresholds which were equivalent to normative data (Priesol et al. 2014). However, both patients included in this study underwent bilateral trans-cochlear nerve sections with labyrinthectomies, ensuring complete bilateral vestibular deafferentation. Both of these patients are current auditory brainstem implant (ABI) users and were included in previous studies of vestibular contributions to motion perception (Valko et al. 2012). These patients were identified through an earlier nationwide search, and substantial efforts were undertaken to find additional patients meeting this strict inclusion criteria. However, we were unable to identify additional qualifying patients locally or nationally who were willing to participate in our research efforts.
Patient A is a 34-year-old female who underwent right- and left-side labyrinthectomies at age 9 and 18, respectively. She currently competes in triathlons and is an avid runner. Patient B is a 37 year-old male. He underwent labyrinthectomies for the right-side at age 5 and at age 20 for the left-side. He reports difficulty perceiving motion when flying and has limp secondary to spinal tumors leading to lower limb weakness. However, he denies difficulty with his balance on a daily basis, is an avid traveler, and is able to ride a bicycle. Previously, as part of the initial study for which these participants were recruited (Valko et al. 2012), both underwent thorough neurological examinations in addition to Semmes–Weinstein monofilament testing to determine pressure sensation of the trunk and buttocks. Despite spinal tumors, no significant differences between both patients to healthy control subjects were noted on pressure thresholds. As well, outside of hearing and vestibular impairments, no significant sensory abnormalities were noted in either patient (Valko et al. 2012).
As the outlined experimental protocol encompassed approximately 5 h of testing for the two bilateral loss patients, roll tilt and roll rotation thresholds and the thresholds for angular velocity stimuli aligned in the planes of the semicircular canals were assessed in two different groups of healthy controls. Roll tilt and roll rotation thresholds were assessed in a group of ten young adults (6F/4M; 27.23 ± 4.77 years); a subset (n = 6) of these data were previously published (Wagner et al. 2022). Thresholds assessing functional canal pairs (i.e., yaw rotation, LARP tilts, RALP tilts) were completed in 32 young adults (15F/17M; 28.10 ± 4.39) as part of a larger ongoing experimental protocol. All control participants denied a history of vestibular disorders, neurological disorders, major health conditions (e.g., cancer), or recent orthopedic injury. All participants provided written informed consent. This study was approved by the Ohio State University Institutional Review Board and was performed in line with the principles of the Declaration of Helsinki.
Motion stimuli and psychophysical threshold tests
Thresholds assessing roll stimuli were assessed using three different motions: (1) “roll tilt” stimuli (i.e., earth-horizontal rotations about a naso-occipital axis) in which the subject was tilted from an upright position, provided a response, and then returned upright prior to the next stimulus, (2) “roll tilt with return” in which the subject was tilted from and returned to upright prior to providing a response, and (3) “roll rotation” thresholds utilizing roll rotations about an earth-vertical axis while supine. Figure 1 depicts roll rotation and generalized roll tilt motion stimuli. Figure 2 provides a depiction of the two roll tilt trajectories.

Depiction of the two roll tilt stimuli experimental paradigms. For both trajectories, approximate head in space is depicted in the upper panel for a leftward tilt while stimulus tilt angle is depicted in the lower panel. For roll tilt stimuli (A), subjects were tilted from upright and provided a response indicating perceived direction of tilt. After the response, subjects were immediately returned to upright prior to the next stimulus presentation. For roll tilt with return stimuli (B), subjects were tilted from upright and then returned to the upright position. After returning to upright, subjects provided response indicating perceived direction of tilt away from upright
Past assessment of roll tilt thresholds in bilateral vestibular loss revealed the smallest elevations relative to other motion stimuli (specifically yaw rotations, y-translation, z-translations) (Valko et al. 2012). This, in turn, suggests an increased contribution of non-vestibular cues. Therefore, we implemented the previously described “roll tilt with return” trajectory to reduce the impact of static cues (e.g., tactile) on motion perception. All three motions were assessed at 0.5 Hz and 2 Hz to assess potential frequency effects. Past data in healthy participants, suggests that tilt responses at 0.5 Hz reflect integration of canal and otolith cues, as tilt thresholds are lower than rotation thresholds, while tilt responses at 2 Hz are dominated by canal angular velocity cues, as thresholds measured using tilt and rotation stimuli are equivalent (Lim et al. 2017; Wagner et al. 2022).
Additionally, thresholds for angular velocity rotations that aimed to predominantly isolate the functional canal pairs were assessed. The horizontal canals were assessed using 2 Hz yaw rotations and vertical canal pairs were assessed using 2 Hz RALP (right-anterior left-posterior) and LARP (left-anterior right-posterior) tilts (i.e., earth-horizontal rotations aligned 45° between the roll and pitch planes). A frequency of 2 Hz was used as past results in healthy controls suggest that, at this frequency, RALP and LARP tilts reflect predominantly canal function, without substantial otolith contributions (Wagner et al. 2022).
The psychophysical procedures and motion paradigms implemented have been previously published in detail (e.g., Chaudhuri et al. 2013; Grabherr et al. 2008; Karmali et al. 2016). Vestibular perceptual thresholds were quantified using a standard one-interval direction recognition task (e.g., “did I turn right or left?”). Subjects reported perceived motion using buttons in each hand. For some motion conditions, Patient B experienced difficulty pressing buttons after prolonged testing secondary to right hand weakness and verbally reported perceived motion direction that was recorded by the experimenters. All participants were seated in a chair secured to a Moog (Aurora, NY) 6DOF platform. Participants were secured using a five-point harness and the head was secured in a motorcycle helmet fixed to the chair. In order to maximize vestibular contributions to motion testing, the test room was completely dark to eliminate visual cues. To mask directional auditory cues, in addition to passive attenuation provided by the helmet and insert earphones, ~ 60 dB Sound Pressure Level (SPL) of white noise was presented during motion trajectories. Both bilateral loss patients wore their ABI processor during testing and white noise was provided via Bluetooth streaming in order to provide a cue for motion onset and offset.
Provided motion stimuli were single cycles of sinusoidal acceleration ([a(t) = A sin(2πft)], A = amplitude, f = motion frequency) in which peak acceleration (A), peak velocity (vp), and total displacement are proportional (vp = A/πf = 2f∆p or ∆p = 2fvp = A/2πf2). This yields a unidirectional bell-shaped velocity trajectory that yields monotonic unidirectional displacement (Benson et al. 1986; Grabherr et al. 2008). Roll rotation and tilt thresholds began at a starting displacement of 5° for 0.5 Hz stimuli and 2.5° for 2 Hz stimuli. Yaw rotation, RALP tilt, and LARP tilt began at 2.5°. Starting stimuli were chosen to be above threshold, including for bilateral loss patients for most of the motion trajectories. For each threshold block of 100 trials (i.e., 100 motions), only 1 stimulus type (e.g., 0.5 Hz roll rotation) was provided. Subjects were instructed on which motion was being assessed and practice trials in the light and dark were provided until the participants reported comfort with the task and motion stimuli.
A 4-down/1-up (4D/1U) adaptive staircase procedure was implemented in which stimulus size decreases after four consecutive correct responses and increases after each incorrect response in order to estimate perceptual thresholds. Until the first incorrect answer, a 2D/1U staircase which halved stimulus magnitudes after two correct answers was implemented in order to optimize efficiency of stimulus presentation. After this initial staircase, step sizes and stimulus magnitudes were selected using Parameter Estimation by Sequential Testing (PEST) rules (Taylor and Creelman 1967). On average, each test (consisting of 100 trials) required approximately 16 min for 0.5 Hz stimuli, and approximately 8 min for 2 Hz stimuli. Testing sessions were 2-to-3 h with breaks given between each threshold measure unless shorter testing sessions were requested by the participant.
Data analysis
In order to obtain estimate of thresholds, psychometric curves were fit to the binary (e.g., left/right) experimental data (Chaudhuri and Merfeld 2013; Lim and Merfeld 2012; Merfeld 2011). Gaussian cumulative distribution psychometric functions defined by the standard deviation (σ; threshold) and mean (µ; bias) were fit using a maximum likelihood estimate via a bias-reduced generalized linear model and a probit link function. Precision of estimates can be impacted by inclusion of lapses (i.e., errors made independent of the test stimulus magnitude), thus, a lapse-identification algorithm was implemented (Clark and Merfeld 2021). This algorithm implements a standard delete-one jackknife procedure when fitting the psychometric function to identify probable outliers (i.e., lapses). The standard error of parameter estimates were also calculated using this algorithm (Quenouille 1956; Tukey 1958). For all thresholds presented, results are reported in terms of peak stimulus velocity (i.e., deg/s) as past data suggest that the brain relies on velocity for motion perception (Grabherr et al. 2008; Valko et al. 2012). All fits were completed in MATLAB using the Statistics and Machine Learning Toolbox version 11.4.
For control participants, thresholds for control subjects displayed a lognormal distribution in line with earlier studies (Benson et al. 1986; Bermúdez Rey et al. 2016). Thus, we report geometric means and the corresponding 95% confidence intervals (CIs), and statistical analyses for the control participants were performed using log-transformed threshold data. Separate linear mixed effects models were used to assess thresholds for our control participant cohorts. For all models, subject was included as a random effect and fixed effects included motion trajectory (i.e., yaw, RALP, or LARP; roll tilt, roll tilt with return, or supine roll rotation). For roll rotation and roll tilt thresholds, frequency and a frequency by trajectory interaction were included as fixed effects in order to assess potential differences between motion trajectories on the basis of stimulus frequency. Due to the small sample size, the Kenward-Roger method (Kenward and Roger 1997) was used to approximate the denominator degrees of freedom of the F distribution for all analyses. All models were fit via restricted maximum likelihood methods. Post hoc testing with Bonferroni adjustments for multiple comparisons were made when appropriate.
For our patients, due to the small sample size, 95% confidence intervals (CIs) of the threshold parameter using the estimated standard error from the jack knife fit were calculated for each motion condition. Thresholds were considered meaningfully different from each other if the 95% CIs did not overlap. While a conservative approach, this enabled qualitative assessment of whether differences in thresholds were solely by chance. To assess our main hypotheses assessing vestibular contributions to roll rotation, three separate comparisons were assessed: (1) impact of axis of rotation was assessed by comparing 95% CIs of roll tilt thresholds to roll rotation thresholds for both 0.5 Hz and 2 Hz stimuli, (2) impact of frequency was assessed by comparing 0.5–2 Hz for roll tilt and roll rotation thresholds, (3) impact of roll tilt trajectory was evaluated by comparing roll tilt and roll tilt with return thresholds at both frequency stimuli. Finally, to assess vestibular contributions to angular velocity thresholds aligned in the three semicircular canal pairs, we compared 95% CIs between yaw rotation, RALP tilt, and LARP tilt thresholds.
Results
Roll rotation and roll tilt thresholds
Roll rotation and roll tilt thresholds for control participants (n = 10) and patients A and B are shown in Table 1, and biases are shown in Table 2. Thresholds normalized as a ratio relative to the geometric mean of healthy controls are shown in Table 3. Figure 3 displays all roll thresholds across both frequencies and Fig. 4 displays each roll motion (supine roll rotation, roll tilt, roll tilt with return) for control participants and each patient. In bilateral loss patients, roll thresholds were approximately 2–8 times higher relative to control participants. No significant overlap was noted between the 95% confidence intervals for the control participants and the patients, indicative of a meaningful elevation in thresholds, supporting that vestibular inputs were the predominant contributors to roll motion perception for each of the six roll conditions (supine roll rotation, roll tilt, and roll tilt with return—each at both 0.5 and 2 Hz).

Average thresholds for control participants, Patient A and Patient B with complete bilateral vestibular loss at 0.5 Hz and 2 Hz for supine roll rotation (panel A), roll tilt (panel B), and roll tilt with return (panel C). Error bars for controls represent 95% CIs of the mean. For the patients, error bars represent the 95% CI of the threshold parameter estimate

Average thresholds for control participants (panel A) and Patient A (panel B) and Patient B (panel C) with complete bilateral vestibular loss for supine roll rotation, roll tilt, and roll tilt with return at 0.5 Hz and 2 Hz. Error bars for controls represent 95% CIs of the mean. For the patients, error bars represent the 95% CI of the threshold parameter estimate
Impact of axis of rotation
In healthy control participants, our results are consistent with our reported past findings that include a subset (n = 6) of this dataset (Wagner et al. 2022). Overall, there was a significant main effect of axis of rotation (F(1,67.92) = 4.79, p = 0.0321) as supine roll rotation thresholds (i.e., earth-vertical axis of rotation) were significantly higher than upright roll tilt thresholds (i.e., earth-horizontal axis of rotation). A significant frequency by axis of rotation interaction was also seen (F(2, 51.08) = 7.56, p = 0.0013). Roll rotation thresholds at 0.5 Hz were higher than 0.5 Hz upright roll tilt and roll tilt with return thresholds (F(1,65.02) = 9.79, p = 0.0026). Thresholds for 2 Hz roll rotation, 2 Hz roll tilt, and 2 Hz roll tilt with return were not significantly different (F(1,65.12) = 0.01, p > 0.99) (Fig. 3).
In both of the patients with bilateral vestibular loss, a qualitatively similar pattern to our control participants was noted. Thresholds for 0.5 Hz supine roll rotation thresholds were meaningfully higher than both 0.5 Hz roll tilt and 0.5 Hz roll tilt with return thresholds. Roll rotation thresholds for 0.5 Hz stimuli were ~ 8–14 times higher in comparison to normative data while both 0.5 Hz roll tilt thresholds (with and without return) were only ~ 2 times higher. At 2 Hz, Patient B displayed equivalent thresholds for roll rotation, roll tilt, and roll tilt with return. However, for Patient A, 2 Hz roll tilt thresholds for both motion trajectories (i.e., with and without return) were higher than supine roll rotation, as indicated by non-overlapping 95% CIs, suggesting increased sensitivity for higher frequency supine rotations relative to upright tilt thresholds (Fig. 4).
Impact of stimulus frequency
In our control participants, a significant main effect of frequency (F(1,61.39) = 9.17, p = 0.004) was identified across all thresholds, as 2 Hz thresholds were overall lower than 0.5 Hz thresholds (Fig. 3). Roll tilt thresholds were equivalent at 0.5 Hz and 2 Hz for both roll tilt (F(1,51.01) = 0.40 p > 0.99) and roll tilt with return (F(1,51.01) = 1.86, p = 0.452) motion trajectories. However, for supine roll rotations, 2 Hz thresholds were significantly lower than 0.5 Hz thresholds (F(1,51.01) = 43.03, p < 0.001). These results are consistent with a previously published dataset that included a subset of these data (Wagner et al. 2022).
For roll rotations, in our patients with bilateral vestibular loss, 2 Hz thresholds were meaningfully lower than 0.5 Hz thresholds for both Patient A and Patient B. As well, roll tilt with return thresholds for 2 Hz were equivalent to 0.5 Hz thresholds, as indicated by overlapping CIs. However, for roll tilt stimuli, 2 Hz thresholds were meaningfully higher than 0.5 Hz thresholds for both patients.
Impact of roll tilt stimuli paradigm
A significant main effect of trajectory (i.e., roll tilt vs. roll tilt with return) was not seen for healthy controls, as roll tilt and roll tilt with return thresholds were equivalent (F(1,61.46) = 0.05, p = 0.8265) and no significant differences between thresholds for each paradigm were seen at either 0.5 Hz and 2 Hz (p > 0.05). Similarly, for both Patient A and Patient B, thresholds for roll tilt and roll tilt with return were equivalent at 2 Hz, as indicated by overlapping 95% CIs. However, for 0.5 Hz thresholds, Patient A exhibited thresholds for roll tilt with return which were meaningfully elevated relative to roll tilt thresholds by ~ 2.25 times. For Patient B, the 0.5 Hz roll tilt with return threshold was ~ 1.3 times higher than standard roll tilt, which was not considered a meaningful elevation on the basis of overlapping 95% CIs.
Angular velocity thresholds in three canal planes
All 2 Hz angular velocity thresholds for each of the stimuli aligned with the three canal planes for control participants (n = 32) and the two bilateral loss patients are listed in Table 4 and associated biases are listed in Table 5. Thresholds are shown in Fig. 5 for 2 Hz yaw rotation, RALP tilt and LARP tilt. In healthy control participants, 2 Hz yaw rotation thresholds were equivalent to 2 Hz RALP tilt (F(1,74) = 1.02, p = 0.6304) and LARP tilt thresholds (F(1,74) = 4.43, p = 0.1164).

Average angular velocity thresholds for control participants and Patient A and Patient B with complete bilateral vestibular loss for 2 Hz yaw rotation, RALP tilt, and LARP tilt thresholds. Error bars for controls represent 95% CIs of the mean. For the patients, error bars represent the 95% CI of the threshold parameter estimate
Patient A and Patient B had thresholds which were ~ 6–12 times larger than control participants (Table 3). An overlap between 95% CIs between the patients and the controls was not seen for any threshold, suggesting a meaningful elevation. For both Patient A and Patient B, on the basis of 95% CIs, 2 Hz yaw rotation (i.e., earth-vertical rotation) thresholds were meaningfully elevated relative to both 2 Hz RALP tilt and 2 Hz LARP tilt (i.e., earth-horizontal) thresholds, suggesting an impact of axis of rotation. RALP and LARP tilt thresholds were equivalent to each other.
Discussion
Overall, we showed that vestibular tilt and rotation thresholds, quantified using whole-body direction recognition tasks, were approximately 2–35 times larger for individuals with a complete absence of vestibular function in comparison to young, healthy adults of roughly the same age. As these tasks were designed to minimize non-vestibular sensory cues (i.e., tested in complete darkness, white noise to remove auditory cues), this suggests that in the absence of vestibular function, these patients were able to use available pertinent sensory cues (e.g., tactile, somatosensory) to enable motion perception. Since it seems reasonable to presume that these patients utilize every cue possible to substitute for their total vestibular loss, the obtained thresholds for these patients represent the minimal stimulus amplitudes at which we expect non-vestibular cues to contribute for these threshold tasks when performed by participants with healthy vestibular systems. This primary finding of elevated thresholds in complete absence of vestibular function also confirms the predominant contribution of vestibular inputs to threshold level tilt and rotation motion stimuli.
We also showed that thresholds for tilt (i.e., earth-horizontal rotation) stimuli were less influenced by vestibular loss in comparison to rotation (i.e., earth-vertical rotation) stimuli. This was true for multiple trajectories including roll stimuli as well as stimuli aligned in the anatomic planes of the semicircular canals (i.e., RALP and LARP). As well, lower frequency (i.e., 0.5 Hz) rotation stimuli showed larger elevations in thresholds relative to high-frequency (i.e., 2 Hz) stimuli in the absence of vestibular function. These findings are in line with past studies (Valko et al. 2012) suggesting increased non-vestibular contributions for tilts (i.e., earth-horizontal rotations) and for high-frequency transient stimuli.
Roll rotation and roll tilt thresholds
Impact of axis of rotation
In line with past data (Lim et al. 2017; Wagner et al. 2022), our control subjects demonstrated reduced thresholds for 0.5 Hz roll tilts (i.e., earth-horizontal rotations) relative to roll rotations (i.e., earth-vertical rotations), while roll tilt and roll rotation thresholds were equivalent at 2 Hz. In our bilateral vestibular loss patients, we also identified a reduction in 0.5 Hz roll tilt thresholds for both motion trajectories (i.e., roll tilt and roll tilt with return) in comparison to 0.5 Hz supine roll rotation thresholds.
While a similar trend was identified between our bilateral loss patients and control participants, the magnitude of this effect was starkly different. For the bilateral loss patients, 0.5 Hz roll rotations were approximately 10 × higher than 0.5 Hz roll tilt thresholds while this difference was only approximately a factor of two in healthy adults, suggesting different mechanisms may be driving this effect. Enhanced perception of low-frequency tilts in individuals with intact vestibular function has been posited to reflect multi-sensory integration of canal and otolith cues (Lim et al. 2017; Wagner et al. 2022). For tilt stimuli (i.e., earth-horizontal rotations), which by definition contain rotational cues and re-orientations of the head relative to gravity, the tilt cue is four times larger for 0.5 Hz stimulus than a 2 Hz stimulus for an equal peak velocity stimulus. At lower frequencies of rotation, the canals are less sensitive to rotational stimuli, thus, the absolute displacement of the head relative to gravity increases to approach the level at which otolithic afferents are sensitive to tilts (Lim et al. 2017; Wagner et al. 2022). In our bilateral loss patients (i.e., without otolith afferents), the larger tilt components at lower frequency stimuli may instead introduce more prominent tactile cues (e.g., pressure of shoulders on side of chair, shifting of weight through the back and/or buttocks). Thus, enhanced tilt perception relative to rotation perception (i.e., changes in axis of rotation) at low frequencies in healthy controls likely reflects beneficial contributions of the otoliths enhancing roll tilt motion perception relative to roll rotation, while this qualitatively similar phenomenon in the bilateral loss patients without otolith contributions likely reflects increased availability of tactile cues for the tilts experienced at the higher threshold levels for these patients.
Change in the axis of rotation for a given plane of motion is also accompanied by changes in body orientation (i.e., roll tilt is measured while upright, roll rotation is measured while supine). Past studies of visual motion and whole-body motion perception have posited that absolute body orientation may influence perceptual sensitivity with non-upright orientations leading to a reduction in the reliability of perceptual estimates (Graybiel and Patterson 1955; Hummel et al. 2016; MacNeilage et al. 2010; Mikellidou et al. 2015). While 0.5 Hz rotation thresholds measured while supine were higher than 0.5 Hz tilt thresholds measured while upright, 2 Hz rotation and tilt thresholds were equivalent, despite differences in body orientation during measurement. As such, our data do not support a generalized decrease in perceptual sensitivity when thresholds are measured with the body oriented in non-upright positions (e.g., supine).
Impact of stimulus frequency
For young healthy adults, roll rotation velocity thresholds demonstrated behavior consistent with a high pass filter—as thresholds were lower for higher frequency stimuli (i.e., 2 Hz) relative to lower frequencies (i.e., 0.5 Hz) (Lim et al. 2017; Wagner et al. 2022). Qualitatively similar to our control data, in patients with complete bilateral vestibular loss, we showed that 0.5 Hz supine roll rotation thresholds were meaningfully higher than 2 Hz thresholds. However, for our bilateral loss patients, supine 0.5 Hz roll rotation thresholds were approximately 10 × higher than 2 Hz thresholds whereas for our control subjects 0.5 Hz roll rotation thresholds were only approximately 2 × higher.
These results in our bilateral loss patients are consistent with past threshold assessments across a wider range of frequencies (0.5–5 Hz) which similarly identified a decrease in thresholds with increasing frequency for stimuli introducing transient (i.e. non-static) motion cues including yaw rotation, z-translation, and y-translation (Valko et al. 2012). For roll rotation, as the participants are supine, the higher frequency motion directions may induce larger forces on the body and/or more shifting of the body in the chair and thereby may provide more useful tactile cues. Thus, while the modulation of roll rotation thresholds on the basis on stimulus frequency in younger adults likely reflects differences in the sensitivity to angular velocity signal originating from the vertical canals, in the absence of vestibular function, this may instead be a result of the increased availability of tactile or alternative non-vestibular cues during the higher frequency transient motion stimuli.
For healthy controls, thresholds for 0.5 Hz and 2 Hz roll tilt and roll tilt with return motion trajectories were equivalent—we have also previously identified these behaviors for multiple planes of motion (i.e., roll, pitch, RALP, LARP) for traditional tilt stimuli (Wagner et al. 2022). Also, qualitatively similar to our control data, equivalent thresholds were seen for 0.5 and 2 Hz roll tilt with return (but not roll tilt) thresholds in our bilateral loss patients. This suggests that the availability of non-vestibular cues was not modulated on the basis of stimulus frequency in the absence of a static tilt cue. However, for traditional roll tilt thresholds, 2 Hz thresholds were significantly higher than 0.5 Hz thresholds, potentially suggesting that, in the absence of vestibular function, the static tilt force introduced by the roll tilt stimuli contributed to lower frequency tilts, but not for higher frequency tilt stimuli. This difference modulated on the basis of frequency may suggest a difference in temporal integration of non-vestibular cues, particularly tactile cues. This may also simply reflect that forces are proportional to acceleration and acceleration increases with frequency for the same peak velocity. As roll tilt and roll tilt with return thresholds are equivalent at 2 Hz in our bilateral loss patients, indicating by overlapping 95% CIs, this may instead solely reflect inherent variability in performance and our small sample size.
Impact of motion trajectory
Previous assessment of vestibular thresholds in patients without vestibular function revealed the smallest impact of vestibular ablation relative to normative data for the perception of roll tilt stimuli (Valko et al. 2012). As briefly discussed in “Methods”, this previous study employed a commonly implemented roll tilt paradigm (Fig. 2A) which introduces a static tilt cue as the subject is tilted from upright and provides a response indicating the perceived direction of motion prior to returning to upright. We, therefore, implemented a novel tilt paradigm in which subjects were tilted and returned almost immediately to upright prior to providing a response (“roll tilt with return”, Fig. 2B). This motion trajectory paradigm essentially removes the availability of a static tilt cue by providing a nearly continuous angular rotation stimulus with maximal static tilt occurring transiently for just a fraction (i.e., 1/60th) of a second. For a constant peak velocity stimulus, the tilt cue is 4 × smaller at 2 Hz than 0.5 Hz, thus, if static tilt cues are contributing to perceptual sensitivity, we would expect to see to see a larger impact of this novel stimuli for 0.5 Hz than 2 Hz tilts.
In healthy controls, although thresholds were ~ 1.2 times higher for roll tilt with return motion in comparison to roll tilt thresholds in healthy controls, no statically significant differences were noted in thresholds measured using these two motion trajectories for either both 0.5 Hz or 2 Hz tilts. This is in agreement with our past reported finding in a smaller sample size (n = 6) which were also included in these analyses for multiple planes of motion (Wagner et al. 2022), this suggests that removal of the static tilt cue through this paradigm does not consistently influence perceptual sensitivity in individuals with intact vestibular function.
In our bilateral vestibular loss patients, roll tilt thresholds were overall elevated by approximately 2.3–6.5 times relative to our normative data, while roll tilt with return thresholds were approximately 1.8–10.3 times higher. As thresholds are elevated with removal of vestibular cues, this suggests that both motion paradigms capture a portion of vestibular contributions to motion perception. As well, while roll tilt with return thresholds tended to be higher than traditional roll tilt thresholds by ~ 1.5 times, no meaningful differences between thresholds obtained from the different paradigms were seen at 2 Hz for either of our bilateral loss patients, as indicated by overlapping CIs. However, for 0.5 Hz stimuli, where a potential impact of motion trajectory is expected to be captured due to the larger tilt displacement, Patient A exhibited a threshold which was ~ 2.5 times higher for roll tilt with return in comparison to roll tilt. This elevation was considered a meaningful differences on the basis of our conservative analysis. Patient B demonstrated a 0.5 Hz roll tilt with return threshold which was ~ 1.3 times higher than the traditional roll tilt paradigm, which was not considered a meaningful elevation on the basis of overlapping CIs. As one of our bilateral loss patients demonstrated an elevation in threshold with removal of the static tilt cue for 0.5 Hz stimuli using the roll tilt with return paradigm, this suggests that static tilt cues may influence perceptual sensitivity at higher tilt displacements (i.e., at lower frequencies). However, as disparate behaviors were exhibited between our two patients, this methodological question pertaining to best practices for the delivery of tilt stimuli should be further explored. Particularly, response confidence may play a role in influencing thresholds obtained during these paradigms. In the tilt with return paradigm, the participants may be able to resolve motion ambiguity due to the longer exposure to motion and potential comparisons between the tilt stimulus and return to center. A measure of confidence in response may more completely capture differences in motion perception obtained through these two paradigms.
Angular velocity thresholds in three-canal planes
In our healthy control participants, 2 Hz yaw rotation thresholds were equivalent to 2 Hz RALP tilt and 2 Hz LARP tilt thresholds, suggesting that for high-frequency motion stimuli, velocity thresholds targeting the horizontal canals are equivalent to thresholds targeting the vertical canal pairs. In our bilateral loss patients, yaw rotation thresholds were meaningfully higher than both RALP and LARP tilts, while both vertical canal measures (i.e., RALP and LARP tilt thresholds) were equivalent. This suggests greater non-vestibular contributions to tilts (i.e., earth-horizontal rotations), potentially reflecting contributions of the static tilt and increased useful tactile cues (e.g., pressure cues on the head from the helmet) introduced by the tilt stimuli. Of note, during daily locomotion, the predominant frequency of yaw head rotations is ~ 1 Hz compared to ~ 2 Hz for pitch head rotations (Grossman et al. 1988; Carriot et al. 2014; Zobeiri et al. 2021). Thus, our bilateral loss patients routinely experience head pitch at frequencies assessed in this paradigm and less routinely experience yaw rotations at the tested frequency. As patients also presumably must up-weight non-vestibular cues for motion perception more so than those with a healthy vestibular system, regular exposure to head pitch at the assessed frequency may have played a role in facilitating non-vestibular motion perception.
As the obtained threshold measures are ~ 6–12 times larger in bilateral loss patients, the concomitant tilt stimulus is substantially larger than those experienced at threshold level for individuals with intact vestibular function, suggesting that these non-vestibular cues can contribute to perception of tilts at relatively large stimulus magnitudes. However, as all thresholds were meaningfully higher in bilateral loss patients, and healthy controls responded at much smaller tilt magnitudes with putatively decreased contributions of tactile cues, this suggests that in healthy controls, angular velocity thresholds in the planes of the vertical semicircular canals are predominantly mediated by vestibular inputs.
While we found similar perceptual thresholds for motions aligned in the planes of the horizontal and vertical canals in healthy controls, afferent recordings suggest that the vertical canals display increased sensitivity relative to the horizontal canals in animals (Yang and Hullar 2007). This suggests that perceptual thresholds in humans did not capture potential differences seen in afferent sensitivity between peripheral end-organs in animal models. This finding is in contrast to translation thresholds (Agrawal et al. 2013; Kobel et al. 2021a), which have revealed differences in measures with predominant utricular or saccular contributions that mirror known differences in peripheral afferent sensitivity believed to reflect differences in maculae size between the otoliths. In past animal models, vestibular afferent sensitivity has been previously demonstrated to be proportional to canal radius (ten Kate et al. 1970; Yang and Hullar 2007). In multiple species, including humans, the horizontal canals have been found to have demonstrated a smaller radius in comparison to vertical canals (Curthoys et al. 1977; ten Kate et al. 1970). On the basis of observed canal radii in humans (Curthoys et al. 1977), vertical canal sensitivity should be ~ 1.25 times higher than the sensitivity of the horizontal canals. However, on the basis of estimated macula size (Naganuma et al. 2001, 2003), utricular sensitivity should be ~ 2 times higher than saccular sensitivity in humans. As such, the larger difference between utricular and saccular sensitivity may have allowed identification via behavioral perceptual measures. Future studies should investigate the differences between vestibular thresholds that preferentially target the semicircular canals across a wider range of frequencies to fully assess behavioral differences between horizontal and vertical canal measures.
Limitations and future work
A limitation of this study was the small number of bilateral loss patients (n = 2) that we were able to include in this analysis. However, we required a complete bilateral absence of vestibular function in order to state with certainty that the thresholds measured reflected the maximal extent to which extra-vestibular sensory systems could contribute to motion perception. This approach was taken in order to permit the identification of stimulus magnitudes at which these non-vestibular contributions (e.g., tactile, somatic graviception) become relevant for motion perception. Both of the patients included in this current study were included in an initial study quantifying vestibular contributions to motion perception across a wider frequency range (Valko et al. 2012). These patients were selected on the basis of having undergone bilateral labyrinthectomies, and not solely bilateral nerve sections, in order to ensure complete bilateral vestibular deafferentation. We attempted to identify additional patients with total bilateral vestibular ablation both locally and nationally through multiple searches over several years, but we were unable to do so, reflecting the rarity of this patient population. Due to the large differences in vestibular thresholds relative to normative data, we posit that our conclusion that the vestibular system serves as the primary contributor to rotation and tilt motion perception is valid. However, we acknowledge that we cannot conclusively determine that those with healthy vestibular systems are solely using vestibular cues for motion perception during these perceptual tasks despite experimental controls to limit non-vestibular sensory contributions. We can only conclusively state that the thresholds obtained in our healthy participants were well below the level at which sensory information became available and useful for motion perception in our bilateral loss patients.
Additionally, our two patients are exceptional performers in terms of their daily function. Particularly, Patient A is competing at a high level of athleticism and Patient B is a frequent, independent traveler. As well, as both underwent labyrinthectomies over 15 years prior, they have likely optimized use of non-vestibular sensory cues for motion perception and postural control. While our experimental design aimed to limit non-vestibular sensory contributions through testing in complete darkness, thus removing visual cues, and through playing auditory white noise during motion, reducing useful auditory cues, the interface between the body and the chair for testing is unavoidable. Thus, in our experimental paradigm, tactile cues for motion perception cannot be eliminated. As our bilateral loss patients likely exhibit an exceptional ability to use non-vestibular cues for motion perception, it is possible that they were able to use non-vestibular cues to a greater extent than the healthy controls.
As well, in the included patients, a neurological exam and test of pressure sensation was last assessed almost 10 years prior when both participants previously completed vestibular perceptual threshold testing (Valko et al. 2012). In theory, changes in sensory function from these initial assessments could have influenced our results. However, overall elevations in roll tilt and yaw rotation thresholds relative to normative samples were similar between time points, suggesting that other changes in sensory function were unlikely to impact our results.
A more complete study design would have included measuring RALP and LARP rotations (i.e., rotations about an earth-vertical axis while supine and rotated 45 degrees) in addition to the measured tilts in our bilateral loss patients to more clearly target isolated vertical canal function. However, bilateral loss patients completed the current test battery (~ 5 h) in addition to a separate test battery assessing translation perception (~ 10 h). Thus, these participants completed approximately 15 h of testing over several days, while visiting the Ohio State University from out-of-state. We chose to not include RALP and LARP rotations as our past research suggested that plane of motion (i.e., roll, pitch, RALP, LARP) did not systematically impact rotation or tilt thresholds (Wagner et al. 2022). For our patients, 2 Hz roll tilt thresholds, for both motion trajectories, were equivalent to RALP and LARP tilt thresholds, suggesting a similar lack of impact of motion plane in our bilateral loss patients. However, future studies should directly test the potential effect of motion plane on tilt and rotation thresholds.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Agrawal Y, Bremova T, Kremmyda O, Strupp M, MacNeilage PR (2013) Clinical testing of otolith function: perceptual thresholds and myogenic potentials. JARO J Assoc Res Otolaryngol 14:905–915
Benson AJ, Spencer MB, Stott JR (1986) Thresholds for the detection of the direction of whole-body, linear movement in the horizontal plane. Aviation, space, and environmental medicine 57:1088–1096
Bermúdez Rey MC, Clark TK, Wang W, Leeder T, Bian Y, Merfeld DM (2016) Vestibular perceptual thresholds increase above the age of 40. Front Neurol 7
Bremova T, Caushaj A, Ertl M, Strobl R, Böttcher N, Strupp M, MacNeilage PR (2016) Comparison of linear motion perception thresholds in vestibular migraine and Menière’s disease. Eur Arch Otorhinolaryngol 273:2931–2939
Carriot J, Jamali M, Chacron MJ, Cullen KE (2014) Statistics of the Vestibular Input Experienced during Natural Self-Motion: Implications for Neural Processing. J Neurosci 34:8347–8357. https://doi.org/10.1523/JNEUROSCI.0692-14.2014
Chaudhuri SE, Merfeld DM (2013) Signal detection theory and vestibular perception: III. Estimating unbiased fit parameters for psychometric functions. Exp Brain Res 225:133–146
Chaudhuri SE, Karmali F, Merfeld DM (2013) Whole body motion-detection tasks can yield much lower thresholds than direction-recognition tasks: implications for the role of vibration. J Neurophysiol 110:2764–2772
Clark TK, Merfeld DM (2021) Statistical approaches to identifying lapses in psychometric response data. Psychonomic Bull Rev 28:1–25
Curthoys IS, Blanks RHI, Markham CH (1977) Semicircular canal functional anatomy in cat, Guinea pig and man. Acta Otolaryngol 83:258–265
Fernandez C, Goldberg JM (1971) Physiology of peripheral neurons innervating semicircular canals of the squirrel monkey. II. Response to sinusoidal stimulation and dynamics of peripheral vestibular system. J Neurophysiol 34:661–675
Grabherr L, Nicoucar K, Mast FW, Merfeld DM (2008) Vestibular thresholds for yaw rotation about an earth-vertical axis as a function of frequency. Exp Brain Res 186:677–681
Graybiel A, Patterson JR JL (1955) Thresholds of stimulation of the otolith organs as indicated by the oculogravic illusion. J Appl Physiol 7:666–670
Grossman GE, Leigh RJ, Abel LA, Lanska DJ, Thurston SE (1988) Frequency and velocity of rotational head perturbations during locomotion. Exp Brain Res 70:470–476. https://doi.org/10.1007/BF00247595
Hummel N, Cuturi LF, MacNeilage PR, Flanagin VL (2016) The effect of supine body position on human heading perception. J vis 16:19–19
Karmali F, Chaudhuri SE, Yi Y, Merfeld DM (2016) Determining thresholds using adaptive procedures and psychometric fits: evaluating efficiency using theory, simulations, and human experiments. Exp Brain Res 234:773–789
Karmali F, Goodworth AD, Valko Y, Leeder T, Peterka RJ, Merfeld DM (2021) The role of vestibular cues in postural sway. J Neurophysiol 125:672–686
Kenward MG, Roger JH (1997) Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 53:983–997
King S, Priesol AJ, Davidi SE, Merfeld DM, Ehtemam F, Lewis RF (2019) Self-motion perception is sensitized in vestibular migraine: pathophysiologic and clinical implications. Sci Rep 9:1–12
Kobel MJ, Wagner AR, Merfeld DM (2021a) Impact of gravity on the perception of linear motion. J Neurophysiol 126:875
Kobel MJ, Wagner AR, Merfeld DM, Mattingly JK (2021b) Vestibular thresholds: a review of advances and challenges in clinical applications. Front Neurol 12:203
Lim K, Merfeld DM (2012) Signal detection theory and vestibular perception: II. Fitting perceptual thresholds as a function of frequency. Exp Brain Re 222:303–320
Lim K, Karmali F, Nicoucar K, Merfeld DM (2017) Perceptual precision of passive body tilt is consistent with statistically optimal cue integration. J Neurophysiol 117:2037–2052
MacNeilage PR, Banks MS, DeAngelis GC, Angelaki DE (2010) Vestibular heading discrimination and sensitivity to linear acceleration in head and world coordinates. J Neurosci 30:9084–9094
Merfeld DM (2011) Signal detection theory and vestibular thresholds: I. Basic theory and practical considerations. Exp Brain Res 210:389–405
Mikellidou K, Cicchini GM, Thompson PG, Burr DC (2015) The oblique effect is both allocentric and egocentric. J Vis 15:24–24. https://doi.org/10.1167/15.8.24
Naganuma H, Tokumasu K, Hashimoto S, Okamoto M, Yamashina S (2001) Three-dimensional analysis of morphological aspects of the human saccular macula. Annal Otol Rhinol Laryngol 110:1017–1024
Naganuma H, Tokumasu K, Hashimoto S, Okamoto M, Yamashina S (2003) Three-dimensional analysis of morphological aspects of the human utricular macula. Annal Otol Rhinol Laryngol 112:419–424
Priesol AJ, Valko Y, Merfeld DM, Lewis RF (2014) Motion perception in patients with idiopathic bilateral vestibular hypofunction. Otolaryngol Head Neck Surg (united States) 150:1040–1042
Quenouille MH (1956) Notes on bias in estimation. Biometrika 43:353–360
Suri K, Clark TK (2020) Human vestibular perceptual thresholds for pitch tilt are slightly worse than for roll tilt across a range of frequencies. Exp Brain Res 238:1499–1509
Taylor M, Creelman CD (1967) PEST: Efficient estimates on probability functions. J Acoust Soc Am 41:782–787
ten Kate JH, van Barneveld HH, Kuiper JW (1970) The dimensions and sensitivities of semicircular canals. J Exp Biol 53:501–514
Tukey J (1958) Bias and confidence in not quite large samples. Ann Math Statist 29:614
Valko Y, Lewis RF, Priesol AJ, Merfeld DM (2012) Vestibular labyrinth contributions to human whole-body motion discrimination. J Neurosci 32:13537–13542
Wagner AR, Kobel MJ, Merfeld DM (2021) Impact of canal-otolith integration on postural control. Front Integrative Neurosci 15
Wagner AR, Kobel MJ, Merfeld DM (2022) Impacts of rotation axis and frequency on vestibular perceptual thresholds. Multisens Res 1:1–29
Yang A, Hullar TE (2007) Relationship of semicircular canal size to vestibular-nerve afferent sensitivity in mammals. J Neurophysiol 98:3197–3205
Zobeiri OA, et al (2021) Effects of vestibular neurectomy and neural compensation on head movements in patients undergoing vestibular schwannoma resection. Sci rep 11:1–14
Acknowledgements
We would like to thank our bilateral loss participants for their time, effort, and willingness to travel to participate in our research. We also thank Bob Grimes and Michael Hall for their technical assistance.
Funding
This research was supported by National Institute on Aging R01- AG073113 and Department of Defense Congressionally Directed Medical Research Programs (CDMRP) Award Number W81XWH192000. MK was supported in part by a The Ohio State University Graduate School’s Alumni Grants for Graduate Research and Scholarship (AGGRS) Program.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection was performed by MK and AW. Data analysis and the first draft of the manuscript was written by MK. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All study procedures were approved by the Ohio State University Institutional Review Board.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Additional information
Communicated by Bill J Yates.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kobel, M.J., Wagner, A.R. & Merfeld, D.M. Evaluating vestibular contributions to rotation and tilt perception. Exp Brain Res 241, 1873–1885 (2023). https://doi.org/10.1007/s00221-023-06650-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-023-06650-5