
Abstract
Many human motor skills can be represented as a hierarchical series of movement patterns. Awareness of underlying patterns can improve performance and decrease cognitive load. Subjects (n = 30) tapped a finger sequence with changing stimulus-to-response mapping and a common movement sequence. Thirteen subjects (43 %) became aware that they were tapping a familiar movement sequence during the experiment. Subjects who became aware of the underlying motor pattern tapped with greater kinematic and temporal consistency from task onset, but consistency was not sufficient for awareness. We found no effect of age, musical experience, tapping evenness, or inter-key-interval on awareness of the pattern in the motor response. We propose that temporal or kinematic consistency reinforces a pattern representation, but cognitive engagement with the contents of the sequence is necessary to bring the pattern to conscious awareness. These findings predict benefit for movement strategies that limit temporal and kinematic variability during motor learning.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Many human motor skills can be represented as a hierarchical series of movement patterns or representations. Playing a musical instrument, for example, involves individual elements (a single note), intermediate groups (scales or phrases), and a higher-level multidimensional representation (composition). One advantage of these pattern representations is the increased ability to identify and access underlying patterns or covert rules in the motor sequence (Keele and Jennings 1992; Koch 2007). For example, a musician might be aware that a passage from a piece of music is based on an arpeggio, and he can use the pre-learned fingering sequence to perform the excerpt accurately and efficiently. Awareness of underlying patterns in sequences can improve performance speed (Curran and Keele 1993; Wong et al. 2015), reduce the influence of stimulus conflict (Koch 2007), increase neural efficiency (Pascual-Leone et al. 1994), and decrease cognitive load (Verwey 2015).
Pattern awareness is often studied in finger movement sequences because the movements are quick and the response times are believed to reflect underlying control processes (Verwey et al. 2010). Typically, a cyclical sequence is presented one element at a time and subjects respond with a corresponding finger tap. The sequence may be as long as 12 or more elements, as in the classic serial reaction time task (SRT; Nissen and Bullemer 1987), or as short as six or fewer elements, as in the discrete sequence production task (DSP; Verwey 2015). Subjects who become aware of the cyclical stimulus are said to have explicit or declarative knowledge (Pascual-Leone et al. 1994; Doyon et al. 1997; Verleger et al. 2015).
Despite the performance benefit, not all individuals will become aware of underlying patterns in a sequential task. If subjects are informed of the presence or asked to learn the sequence, the majority will attain explicit knowledge of the cyclical stimulus (Curran and Keele 1993; Howard and Howard 2001; Moisello et al. 2011). However, in the absence of instruction, it is common that fewer than half of naïve subjects will report the cyclical sequence within one practice session (Curran and Keele 1993; Moisello et al. 2011; Boutin et al. 2014; Verwey et al. 2014).
Several behavioral factors have been identified as markers of explicit pattern knowledge of the stimuli. Subjects who attain awareness have been shown to divide or “chunk” individual sequence elements into clusters that can be initiated and processed as a larger element (Frensch et al. 2002; Koch 2007; Yordanova et al. 2015). Verleger et al. (2015) showed that subjects who will become aware have higher P3 wave amplitudes in their event-related potentials (ERPs) than those who will not, reflecting more decision making. These results demonstrate that awareness of underlying task structure relies on cognitive engagement in performance and a conscious effort to solve underlying puzzles.
The application of the SRT and DSP to real-world learning remains unclear. Motor learning outside of experimental settings is different from single-element presentation paradigms. In learning a new skill, stimulus elements are not typically presented individually. The musician, for example, has the entire piece of music presented as the instruction. He can plan upcoming movements in the context of the sequence and perform an appropriate response. The musician that has awareness of the underlying patterns in music (e.g., the location of an arpeggio and the corresponding fingering) is able to perform under less cognitive load than the musician responding to each individual note separately. Furthermore, the order of the stimuli is less important than the response to the stimuli. The audience cares about the motor response of the musician, not the order of notes on the sheet music he is reading.
Our goal in this study was to identify the factors that determine which individuals will reach subjective awareness of underlying patterns when the pattern was not the order of stimuli per se, but rather the order of the response. In our task, unlike in the SRT and DSP, all elements of the sequence prompt were presented simultaneously. Subjects tapped a finger sequence with changing stimulus-to-response mapping and a common movement sequence. We expected to find similar predictors of explicit knowledge at the level of the stimuli. Compared to subjects without awareness of the common movement sequence, we expected aware subjects would be younger (Howard and Howard 2001; Verwey et al. 2011), have more musical experience (Hund-Georgiadis and Yves Von Cramon 1999; Verwey 2015), tap with greater timing variance (Koch 2007; Yordanova et al. 2015), make fewer errors, reach maximum performance speed sooner, and tap faster (Nissen and Bullemer 1987; Willingham et al. 1989).
Methods
Participants
Healthy adults (n = 31) were recruited to participate in this study. We assessed handedness with the Edinburgh handedness inventory (Oldfield 1971). All subjects performed tapping experiments with the dominant hand. We defined musical experience as the number of years of instrumental practice for at least 5–7 days a week (Ollen 2006). To confirm normal working memory capacity, we assessed digit span with a custom-designed script that presented single numbers every 1 s. All subjects provided informed written consent prior to participating. This study was approved by the University of Rochester Research Subjects Review Board.
Procedure
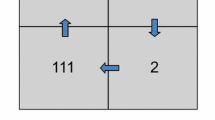
Subjects sat in a chair with feet on the floor, and the dominant forearm rested on a piece of foam about 2 inches thick. We arranged five capacitance switches such that each finger could reach its corresponding switch comfortably. We labeled capacitance switches with the numbers 1 through 5 as shown in Fig. 1. Subjects were allowed to view their hand and the labeled switches throughout the experiment. Tapping prompts appeared as numbers on a computer monitor about 3 feet in front of the subject.

Experimental setup. The display and finger labels on the buttons changed in each block of a sequence, but the actual finger numbers stayed the same. T thumb, I index, M middle, R ring, L little
Subjects wore a Cyberglove (Virtual Technologies, Palo Alto, CA) to measure finger movements. We calibrated the glove to each subject with a procedure similar to previous reports (Hager-Ross and Schieber 2000). Subjects held a series of four standard shapes, while we sampled data from the glove. For each held object, we took a picture in the plane of each finger and measured the joint angles from the metacarpophalangeal (MCP) and proximal interphalangeal (PIP) joints through an image analysis script in MATLAB 8.2 (The Mathworks Inc., Natick, MA, 2013). We also measured the joint angle of the thumb carpometacarpal (CMC) joint. We used linear regression to estimate the relationship between sensor output and joint angle (Hager-Ross and Schieber 2000). In most cases, the glove was too large to render accurate readings from the distal interphalangeal (DIP) joint. Thus, we calculated fingertip position as shown in Fig. 2.

Extrapolation of fingertip location from joint angles (MCP metacarpophalangeal joint, PIP proximal interphalangeal joint). We defined distance d as the length from the PIP to the fingertip with the distal interphalangeal joint straightened
Finger position data were digitally sampled from the Cyberglove at 91 Hz and stored to disk. Contact of a finger with a capacitance switch was detected electronically, time stamped, and stored to disk. Data collection was controlled by Power 1401 and Spike2 version 6 software (Cambridge Electronic Design, Cambridge, UK) with a custom written script.
Tapping sequence
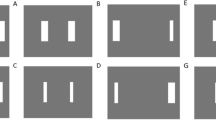
The design of the tapping sequences is shown in Fig. 3. Each subject performed three movement sequences (1, 2, and 3). Each sequence was performed in three blocks (A, B, and C). Each block within a sequence contained a different finger-to-number mapping but consisted of the same sequence of finger movements. Subjects tapped all three blocks of a sequence before moving to the next sequence.

Movement patterns within each sequence are identical, but the displays and finger labels are different. There were a total of 3 sequences (columns) that were each prompted by three different finger-to-number mappings (Blocks A–C, rows). T thumb, I index, M middle, R ring, L little
Each block consisted of at least ten repetitions of a seven-tap sequence. A sequence of seven numbers was displayed on the computer monitor with each number corresponding to one of the five fingers. We instructed the subjects to tap the sequence as quickly and accurately as possible. Correct taps were indicated with high-pitched tones, and incorrect taps were indicated with low-pitched tones. The numbers were displayed on a black background. All the numbers were displayed in white except the instructed one, which was displayed in yellow. After each tap, the next number immediately turned yellow. After the seventh tap in each repetition, the first number turned yellow, prompting the subjects to tap the next repetition.
We defined inter-key-interval (IKI) as the time from one sensor contact to the next sensor contact. Once the performance was stable, as defined by minimum IKI difference between repetitions (a difference smaller than 200 ms and a slope equal to 0), the block ended and the subject moved to the next block. We changed the numbers labeling the finger buttons so that each finger corresponded to a new number. A new display appeared on the monitor, and we instructed subjects to tap with the new finger-to-number mapping. After completion of Block C, Block A of next sequence appeared.
We used the term “awareness” with regard to the motor pattern at the response level to differentiate from “explicit knowledge” which typically refers to the cyclical stimulus. At the conclusion of Blocks B and C in each sequence, we verbally assessed the subjects’ awareness that they had just tapped a sequence they had tapped before (Honda et al. 1998; Moisello et al. 2011). We asked, “did you notice anything special about what you just tapped?” If the subject answered “yes” and was able to tap the sequence from memory, we considered this awareness of the sequence. If the subject answered “no,” we asked whether the sequence was “easier or harder than the one before.”
We designed the sequences so that each sequence had unique finger transitions (for example, an index to middle finger tap appeared only in Sequence 2). Since each sequence was seven numbers long, two fingers tapped twice in each sequence (Sequence 1, middle and ring; Sequence 2, thumb and middle; sequence 3, ring and little). We evaluated both Finger (1–5) and Tap Position (1–7) as independent variables.
Data analysis
We used time stamps from the capacitance switches to measure IKI. For each subject, we calculated the mean IKI over all blocks and sequences. We only measured the IKI between two taps when there was no error. Tapping errors could be errors of substitution (e.g., an index finger tap in place of a middle finger) or of multiple sensors contacted at the same time. We found the mean IKI for each correct tap of the seven-tap positions in each block. We measured IKI evenness by finding the coefficient of variation (CoV; standard deviation divided by the mean) of the mean IKIs in each block. We found the number of repetitions until performance saturation (i.e., IKI was no longer decreasing) by iteratively finding the slope of a robustly fit line through the end of the data. The repetition at which this slope was no longer significantly different than zero was called the saturation point. If the IKI did not decrease with repetition, we set the saturation point to 1.
We measured the temporal consistency in each block through Kendall’s Coefficient of Concordance (Kendall and Smith 1939; Povel and Collard 1982). This procedure ranks the mean IKI for each tap position in a block from 1 (smallest) to 7 (largest) and compares the rank of each tap position across blocks. The Coefficient of Concordance ranges from 0 to 1; the latter indicates complete agreement in timing profiles. We calculated temporal consistency in two ways: (1) within a sequence (A through C in sequence 1, A through C in sequence 2, etc.) and (2) across all nine blocks in all sequences.
We filtered movement data with a fourth-order Butterworth filter with a cutoff frequency of 6 Hz and a sampling frequency of 91 Hz. We used Generalized Procrustes Analysis (GPA) to measure kinematic consistency in each finger (Dryden and Mardia 1998; Loehr and Palmer 2009). Briefly, GPA is a shape-matching process which translates, rotates, and resizes data to minimize differences and finds the root-mean-square of the distance of all the transformed shapes to the mean (or consensus) shape (Fig. 4). We performed GPA on velocity and acceleration trajectories of each fingertip for the 200 ms before the point of minimum acceleration during each correct tap. We chose this length of time to maximize comparability across subjects; GPA requires the same number of data points in each shape, and the majority of IKIs were longer than 200 ms. We used the statistical package “shapes” (Dryden 2015) in R software (R Development Core Team 2015) for GPA analysis. We subtracted the root-mean-square of the Procrustes distance to achieve a Procrustes metric. The Procrustes metric ranges from 0 (no consistency) to 1 (complete consistency). As with the coefficient of concordance, we calculated the Procrustes metric in two ways: (1) for each tap of a finger within a sequence and (2) for each tap of a finger across all sequences.

Example of Generalized Procrustes Analysis for a subject that tapped more consistently (top row) and a subject that tapped less consistently (bottom row). a, c Raw data from a subset of index finger taps. b, d Transformed trajectories. b Procrustes metric = 0.731. d Procrustes metric = 0.402
We used repeated measure ANOVAs to determine whether awareness, sequence, block, and finger or tap position had effects on the CoV, error rate, saturation, and IKI. We included awareness (yes/no) as a between-subject factor even though awareness was an outcome variable (Curran and Keele 1993). Sequence, block, and finger or tap position were within-subject factors. We performed post hoc analyses with paired t tests where appropriate. The nonparametric Friedman test with Conover’s post hoc test was used to compare kinematic consistency across fingers since the Procrustes metric is a bounded measure. All post hoc p values were corrected for multiple comparisons using the Bonferroni method.
We used logistic regression best-subsets model building routine in Statistica (Dell Inc 2015) on data from sequences 1 and 2 to identify predictors of awareness. This routine finds the optimal combination of independent variables to accurately predict the dichotomous outcome. We used the Akaike information criterion (AIC) to determine the best model (Akaike 1974). For validation of the model, we used the data from sequence 3 as a testing set and determined the ability to predict awareness.
Results
Thirty-one subjects completed the experiment. Data from one subject were excluded due to a digit span performance <3. The mean age of the subjects was 40.9 ± 18.6 (mean ± standard deviation, range 18–87 years). The mean number of years of musical practice was 5.0 ± 5.6 (range 0–20 years). There were 18 female subjects (60 %). Three subjects performed the tapping experiments with the left hand (10 %).
Performance variables
Age affected IKI and kinematic consistency. Younger subjects tapped with a lower IKI (r = 0.8, p ≪ 0.0001) and a higher Procrustes metric across all sequences (r = −0.6, p = 0.0002). Older subjects showed a nonsignificant tendency to make more errors (r = 0.3, p = 0.06), but there was no correlation between age and the Coefficient of Concordance across sequences (r = −0.05, p = 0.8), or CoV (r = 0.003, p = 1.0). So although IKI and kinematic consistency were affected by age, error rate, temporal consistency, and tapping evenness were not.
Years of musical practice affected IKI, tapping CoV, and kinematic consistency. There was a significant negative correlation between years of practice and mean IKI (r = −0.5, p = 0.006), indicating that subjects with more years of regular musical instrument practice tapped with a lower IKI. There was a significant correlation between musical practice and CoV (r = −0.4, p = 0.03), indicating that subjects with more musical instrument experience tapped more evenly. There was also a significant negative correlation between years of practice and the Procrustes metric across sequences (r = −0.6, p = 0.0004), indicating that subjects with more musical practice tapped more consistently. There was no correlation between years of musical practice and error rate (r = −0.2, p = 0.4) or years of musical practice and the coefficient of concordance across sequences (r = −0.3, p = 0.2).
There was a significant correlation between IKI and the Procrustes metric across sequences (r = −0.6, p < 0.001), but no correlation between IKI and the coefficient of concordance across sequences (r = −0.3, p = 0.09). There was a positive correlation between coefficient of concordance and CoV (r = 0.7, p ≪ 0.001), indicating that subjects who tapped less evenly tapped with greater temporal consistency across sequences and blocks. We found evidence for a speed-accuracy trade-off, with error rate increasing with IKI (r = 0.5, p = 0.005).
There was a significant main effect of finger on error rate [F(4, 116) = 6.5, p < 0.001], but no main effect of sequence [F(2, 58) = 2.9, p = 0.06] or block [F(2, 58) = 0.3, p = 0.7], or Sequence × Block [F(4, 116) = 1.0, p = 0.4]. This means that error rate did not change with a different finger-to-number mapping. There was no difference in error rate between the same finger tapping in different positions within a sequence (paired t tests with Bonferroni correction). Post hoc analysis indicated that the thumb had the most errors (p < 0.05 compared to all other fingers).
All subjects showed evidence of between-repetition pauses and had similar variance in IKI. Tap position had a large influence on the IKI [F(6, 174) = 41.5, p ≪ 0.001]. Post hoc tests revealed a significantly longer IKI occurring between the last tap of a repetition and the first tap of the next repetition (p ≪ 0.001). There was a significant effect of block on IKI [F(2, 58) = 6.9, p = 0.002], with post hoc tests indicating a longer IKI in Block B than Block A (p = 0.02) and Block C (p = 0.003) in each sequence. There was also a significant interaction of Sequence × Block [F(4, 116) = 6.1, p < 0.001]. Post hoc tests indicated a longer IKI in Block A of sequence 1 than Block A of sequence 2 (p < 0.001) or sequence 3 (p < 0.01). There was also a longer IKI in Block B of Sequence 1 than Block B of Sequence 2 (p < 0.05) or Sequence 3 (p < 0.05). There was no effect of block on CoV across sequences [F(2, 58) = 0.112, p = 0.9], so finger-to-number remapping did not affect the evenness of tapping.
For saturation point, there was a significant effect of Sequence [F(2, 58) = 4.2, p = 0.02] and Block [F(2, 58) = 3.3, p = 0.04], but no significant interaction [Sequence × Block, F(4,116) = 1.7, p = 0.2]. Post hoc tests revealed a later saturation point in Sequence 1 than in Sequence 2 or 3 (p < 0.05). Saturation in Block 3 occurred earlier than in Blocks 1 and 2, but formal post hoc analysis revealed no significant difference on saturation point by Block (p > 0.05). This analysis did not show progressive improvement across sequences and blocks, but did provide some evidence for a practice-based reduction in time to performance plateau.
For the coefficient of concordance within each sequence (comparing Blocks A through C), there was no within-subject effect of sequence [F(2, 58) = 2.0, p = 0.1]. We compared the average coefficient of concordance within each sequence with the coefficient of concordance across all blocks and all sequences. These two values were highly correlated (r = 0.8, p ≪ 0.001). This means that subjects who tapped with the same temporal profile within a given sequence were more likely to tap with the same profile across sequences (i.e., tap position and not finger order determined the timing profile).
There was a significant main effect of finger on the Procrustes metric across sequences [χ 2(4) = 26.3, p ≪ 0.001]. Conover’s post hoc test with Bonferroni correction revealed greater kinematic variability in the little finger compared to all others (p < 0.001). The index finger tapped with the greatest kinematic consistency (p < 0.05).
Awareness
Thirteen subjects (43 %) became aware that they were tapping a familiar movement sequence during the experiment. However, not every aware subject demonstrated awareness in Blocks B and C of every sequence. In other words, awareness in one sequence did not predict awareness in the other sequences. Furthermore, awareness could be lost or gained during the course of the experiment. Of the aware subjects, three (23 %) had awareness during Blocks B and C of every sequence. The remaining ten aware subjects (77 %) had awareness during at least one block of one sequence, but not in every block of every sequence (Fig. 5).

Awareness could be gained and lost throughout experiment. Numbers in circles represent number of subjects. Thirteen subjects (43 %) attained awareness of the common sequence at some point during the experiment. Awareness in one sequence did not guarantee awareness in the next
Age and years of musical practice did not differ between subjects who attained awareness and those who did not. The average age of subjects with awareness was 34.3 ± 15.4, and the average age of subjects without awareness was 46.0 ± 19.6 [t(28) = −1.8, p = 0.1]. The average number of years of musical practice in subjects with awareness was 5.2 ± 5.0, and the average number of years of musical practice in subjects without awareness was 4.9 ± 6.2 [t(28) = 0.2, p = 0.9].
Awareness did not affect error rate [Awareness, F(1,28) = 1.0, p = 0.3; Finger × Awareness, F(4,112) = 0.6, p = 0.7], saturation point [Awareness, F(1,28) = 2.0, p = 0.2; Block × Awareness, F(2, 56) = 1.6, p = 0.2], CoV [Block × Awareness, F(2,56) = 0.9, p = 0.4], the length of the IKI between repetitions [Tap Position × Awareness, F(6,168) = 0.8, p = 0.6], or the length of IKI with finger-to-number remapping [Block × Awareness, F(2, 56) = 1.1, p = 0.3].
Age, musical practice, median IKI, CoV, coefficient of concordance, Procrustes metric, saturation point, and error rate from sequences 1 and 2 were used as factors in the best-subset model builder (Table 1). We used the values for the coefficient of concordance and Procrustes metric as determined within sequences 1 and 2. The model which best predicted awareness contained the coefficient of concordance and the Procrustes metric (AIC = 35.5, Table 2). The next two best models included musical practice and mean ITI, but the Procrustes metric is highly correlated with both these variables. We chose the first model because these correlations suggest that median IKI and musical experience are not independently contributing to the outcome. The raw data from the coefficient of concordance and Procrustes metric in sequences 1 and 2 are shown in Fig. 6a.

Results and probability of awareness. a Raw data for each subjects from sequences 1 and 2. Each dot represents the average coefficient of concordance and Procrustes metric measured within each sequence. Dashed lines indicate median. b Probability curves from the logistic model determined from sequences 1 and 2. Data from sequence 3 are included as validation for the model
To validate our model, we determined the probability of awareness with coefficient of concordance and Procrustes metric from Block 3. This model was also able to correctly predict which subjects would attain awareness in sequence 3 in 80 % of cases (Table 3). The data from sequence 3 are shown with the probability curves from the logistic model determined from sequences 1 and 2 in Fig. 6b.
Discussion
Awareness of underlying patterns in motor sequences is known to improve performance. In this motor task, awareness of the pattern at the motor level was associated with temporal and kinematic consistency. Subjects performed a series of finger-tapping tasks with a common motor response. Unlike SRT tasks where the pattern is at the level of the stimulus, we found no effect of age, musical experience, inter-key-interval, or CoV on awareness. Subjects who eventually became aware of the common movement sequence would either divide the seven-item sequence into the same sub-divisions, or they would move their fingers along the same trajectory for each tap.
None of the subjects with a coefficient of concordance or Procrustes metric below the median in sequences 1 and 2 group attained awareness, indicating that consistency in either the temporal or kinematic domain was necessary for awareness. However, not every subject with a large coefficient of concordance or Procrustes metric had awareness, and awareness could be gained or lost from one block or sequence to the next. In other words, consistency is not sufficient for awareness.
Consistency predicts awareness
Previous studies of explicit pattern knowledge at the stimulus level have indicated a reverse directionality between performance consistency and awareness: that awareness itself leads to more consistent performance (Koch 2007; Verwey et al. 2011; Verwey 2015). This phenomenon has been shown mostly in the temporal domain. Koch (2007) showed that explicit sequence knowledge can lead to the development of motor chunks which reduce stimulus–response conflict. However, explicit knowledge can occur without chunking in patients with basal ganglia stroke (Boyd et al. 2009), so the relationship between chunking and explicit knowledge is still unclear. Yordanova et al. (2015) were able to predict which subjects would reach explicit knowledge of the sequence stimulus through an analysis of tapping evenness. Subjects who tapped with a higher temporal CoV were more likely to realize the underlying sequence pattern. The authors speculated this was due to the increased chunking during motor learning and that the chunking reflected internal hypothesis testing as to the regularity of the sequence.
In our study, temporal and kinematic consistency in the first block of each sequence was able to predict which subjects would go on to attain awareness of the motor pattern. Our findings corroborate the proposal that behavioral differences in motor performance can predict sequence knowledge at task onset (Yordanova et al. 2015). Although CoV was not part of the best predictive model for awareness in our study, CoV was highly correlated with the coefficient of concordance and might be reflecting the same phenomenon. Therefore, we expand the findings of (Yordanova et al. 2015) and suggest that segmentation of sequences into consistent chunks is part of the model that predicts awareness of underlying patterns in the motor response.
Along with temporal consistency, consistency in the kinematic domain in the first block of each sequence can predict pattern awareness. The kinematic trajectory of each finger in subjects with large Procrustes metric subjects remained consistent despite the location of that finger in the tapping sequence. For example, a subject moved the index finger consistently whether the tap was preceded by the middle finger (as in sequence 1) or the thumb (as in sequence 2). We do not know of previous work connecting kinematic consistency to motor pattern awareness at the response level. However, it is well established that deliberate control over movements interrupts automatic motor control processes and results in greater movement variability (Singer et al. 1993; Wulf and Prinz 2001). Automatic control is further supported by the strong correlation between the Procrustes metric and both IKI and musical experience, since automatic control is known to allow faster performance (Bapi et al. 2000; Hikosaka et al. 2002), and more likely to occur if subjects have experience with finger sequences (Hikosaka et al. 2002; Verwey 2003; Parsons et al. 2005; Chaffin et al. 2007; Verwey 2015).
It is possible that greater automatic motor control in subjects tapping with kinematic consistency freed up cognitive resources to search for underlying patterns in the response (Norman and Shallice 1986). Indeed, more autonomous performance has been associated with increased explicit knowledge of a sequence stimulus with incongruent stimulus-to-response mappings (Koch 2007). Again, we expand this finding and suggest that greater kinematic consistency represents greater automatic motor control, which, with temporal consistency, predicts awareness of patterns in the motor response.
Temporal pattern depends on sequence length more than sequence content
We found it surprising that the temporal profile in some subjects remained consistent despite the different finger movements in each sequence. For example, a subject who lengthened the IKI between tap position 3 and 4 did so whether the finger transition was ring to thumb (as in sequence 1) or thumb to index (as in sequence 2). Previous groups have also found that IKI latencies do not rely on finger transitions (Povel and Collard 1982; Loehr and Palmer 2007). Without obvious stimulus divisions (Cohen et al. 1990), the temporal profile or chunking pattern of a sequence is determined by the sequence length and varies by subject (Sakai et al. 2003; Fendrich and Arengo 2004). None of the sequences contained obvious segmentations (e.g., 3–2–3, 1–2–3) or contiguous taps of the same finger, so subjects used the sequence length as the determinate for tapping rhythm.
Temporal profiles in discrete tapping sequences are based on the structural pattern of the sequence and are known to reflect internal hierarchical sequence representation (Povel and Collard 1982; Rosenbaum et al. 1983; Cohen et al. 1990; Koch and Hoffmann 2000; Sakai et al. 2003; Loehr and Palmer 2007). Subjects who had a high coefficient of concordance\tapped with a consistent within-subject temporal profile in each block of each sequence, regardless of finger order. In other words, these subjects maintained a similar representation for each block.
The relationship between temporal and kinematic consistency
The negative relationship between the coefficient of concordance and the Procrustes metric in aware subjects suggests a trade-off between temporal and kinematic consistency, even in Block A of each sequence. Such a trade-off is not evident in subjects without awareness. This further supports that there is a fundamental difference in the performance of aware subjects that precedes awareness. The idea that temporal and kinematic domains are reciprocal has been seen before and likely depends on the task goal (Loehr and Palmer 2007). Balasubramaniam et al. (2004) showed that subjects instructed to move their fingers with high temporal accuracy moved with more variable motion trajectories. However, in the absence of temporal restriction, subjects’ finger trajectory was smoother and more symmetric.
Effect of awareness on learning and performance
All subjects in our task, regardless of awareness, showed evidence of sequence learning. The first time subjects performed this task (Block A of sequence 1), and they tapped with a longer IKI. Similarly, subjects took longer to reach performance plateau in sequences 1 than sequences 2 or 3. In other words, subjects generally tapped faster and plateaued sooner with practice. There was also evidence for a decreasing remapping effect with learning. The first remapping (Block B of sequence 1) resulted in substantial IKI increase, but subsequent remapping had a smaller effect on IKI, especially in Block C of each sequence.
None of this learning evidence was changed with awareness. Aware subjects were not necessarily able to learn the sequence faster, and the benefits of awareness with regard to learning the finger-to-number remappings were limited.
Sequence awareness at the stimulus level is typically associated with performance benefit (Curran and Keele 1993; Koch 2007; Ghilardi et al. 2009; Wong et al. 2015). There is usually a decrease in reaction time with awareness, sometimes accompanied by a stark change in performance indicating the point of knowledge attainment (Brooks et al. 1995; Honda et al. 1998; Frensch et al. 2002). With regard to the directions in our motor task (e.g., “tap as quickly and accurately as possible”), we found no benefit of sequence awareness on performance. Aware subjects did not make fewer errors than non-aware subjects, and the difference in IKI was evident in the first block of each sequence, or before there was even the possibility of awareness. In other words, fast tapping preceded awareness.
The first reason we might not see additional benefit from awareness with regard to error rate or IKI in our task is that the underlying pattern was a movement sequence as opposed to the order of stimuli. We explicitly revealed the sequence and stimulus-to-response mapping in our experiment by labeling each finger button with the new number, reducing the cognitive load since subjects did not have to solve the mapping or sequence itself. Incompatible mapping in the SRT has been shown to drastically reduce explicit knowledge and performance, presumably because of the difficulty in the stimulus-to-response associations (Willingham 1999; Koch and Hoffmann 2000). Since we presented the entire stimulus and the remapping throughout the experiment, the attentional demands were minimized.
Second, the stimuli in our task were symbolic (numbers on a screen) and were mapped to spatial responses (five spatially arranged keys). In many tasks that show a benefit of awareness, both stimuli and responses are arranged spatially (Nissen and Bullemer 1987; Curran and Keele 1993; Wong et al. 2015). Spatial sequences, whether in the stimulus or response, contribute more to sequence learning, facilitate recall, and have a greater influence on response times (Koch and Hoffmann 2000; Verwey et al. 2010). In our task, the spatial responses of each block within a sequence were the same.
Third, subjects might only benefit from awareness if they were not already performing quickly. Verwey (2015) suggested that the influence of sequence awareness on finger-tapping performance is limited since translation of explicit knowledge into performance parameters takes longer than the execution of already consolidated motor movements. Although awareness could promote movement anticipation, subjects could already anticipate movements with the entire sequence prompt presented simultaneously. The lack of effect of awareness on saturation point also suggests that movement anticipation was not a benefit of awareness.
Factors that did not predict awareness
Factors that have been shown to predict explicit knowledge in stimulus–response tasks did not contribute to the logistic model. Specifically, we predicted younger subjects and subjects with more musical experience would have a greater probability of sequence awareness. It is true that younger subjects and subjects with more years of musical instrument practice tended to tap with greater kinematic consistency, so musical experience may make a contribution toward the probability of awareness. However, temporal and kinematic consistency alone was able to predict awareness.
It is possible that there are relevant factors that predict awareness that we did not explore. Furthermore, we cannot rule out that our method of asking awareness did not identify all subjects that knew the sequence was the same. However, our method of probing awareness assessed the subjective awareness of the motor pattern (Boyd et al. 2009). We wanted to know whether, without incentive or prompt, subjects would discover the underlying pattern. Previous groups have confirmed reports of explicit knowledge in a sequence by having a subject writing it down (Verwey et al. 2010; Yordanova et al. 2015), or identify the sequence of interest in a forced-choice or playback scenario (Boyd et al. 2009). It would be interesting to see whether awareness of the motor pattern is still predicted by consistency when subjects are forced to recognize of reproduce the sequence. We also cannot rule out that subjects without awareness would have become aware with repeated exposure, after a delay (Verleger et al. 2015), or after sleep (Darsaud et al. 2011).
Deciding factor for awareness
It is likely that kinematic or temporal consistency reinforced a sequence representation or memory trace, but engagement of higher-level cognitive resources was required to bring pattern regularities to conscious awareness (Dehaene and Naccache 2001; Frensch et al. 2002; Cleeremans and Sarrazin 2007). This engagement can be prompted by unexpected events or a deliberate attempt to uncover the underlying sequence pattern. (Norman and Shallice 1986; Rünger and Frensch 2008; Verleger et al. 2015; Yordanova et al. 2015).
Not all loci for attention in a finger-tapping task will facilitate awareness. For example, (Boutin et al. 2014) found that subjects who are instructed to subjectively assess their performance tapped faster, but were not more likely to have explicit knowledge of the sequence than subjects who were given no instruction. In other words, attention toward the movements does not facilitate awareness. In fact, some explicit knowledge can be gained after simply watching the stimulus without making movements (Willingham 1999), or watching another person make the movements (Heyes and Foster 2002). Awareness seems to occur only after a subject directs attention to the pattern itself.
Conclusions
We have shown that awareness of underlying movement patterns in a finger-tapping task relies on temporal and kinematic consistency. These factors are different than those which predict explicit sequence knowledge in the commonly studied SRT task, in which the pattern exists at the level of the stimulus. Since consistency was not sufficient for awareness, further research is needed to address the factors that promote awareness in subjects tapping with temporal and/or kinematic consistency. Since awareness of underlying patterns in motor sequences is known to be beneficial in motor performance, these findings predict benefit for movement strategies that limit temporal and kinematic variability during motor learning.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723
Balasubramaniam R, Wing AM, Daffertshofer A (2004) Keeping with the beat: movement trajectories contribute to movement timing. Exp Brain Res 159:129–134
Bapi RS, Doya K, Harner AM (2000) Evidence for effector independent and dependent representations and their differential time course of acquisition during motor sequence learning. Exp Brain Res 132:149–162
Boutin A, Blandin Y, Massen C et al (2014) Conscious awareness of action potentiates sensorimotor learning. Cognition 133:1–9
Boyd LA, Edwards JD, Siengsukon CS et al (2009) Motor sequence chunking is impaired by basal ganglia stroke. Neurobiol Learn Mem 92:35–44
Brooks V, Hilperath F, Brooks M (1995) Learning “what” and “how” in a human motor task. Learn Mem 2:225–242
Chaffin R, Lemieux A, Chen C (2007) “It Is Different Each Time I Play”: variability in highly prepared musical performance. Music Percept 24:455–472
Cleeremans A, Sarrazin JC (2007) Time, action, and consciousness. Hum Mov Sci 26:180–202
Cohen A, Ivry RI, Keele SW (1990) Attention and structure in sequence learning. J Exp Psychol Learn Mem Cogn 16:17–30
Curran T, Keele SW (1993) Attentional and nonattentional forms of sequence learning. J Exp Psychol Learn Mem Cogn 19:189–202
Darsaud A, Wagner U, Balteau E et al (2011) Neural precursors of delayed insight. J Cogn Neurosci 23:1900–1910
Dehaene S, Naccache L (2001) Towards a cognitive neuroscience of consciousness: basic evidence and a workspace framework. Cognition 79:1–37
Doyon J, Gaudreau D, Laforce R et al (1997) Role of the striatum, cerebellum, and frontal lobes in the learning of a visuomotor sequence. Brain Cogn 34:218–245
Dryden I, Mardia K (1998) Statistical Shape Analysis, 4th edn. Wiley, Chichester
Dryden IL (2015) Shapes: statistical shape analysis. R package version 1.1-11. http://CRAN.R-project.org/package=shapes
Fendrich DW, Arengo R (2004) The influence of string length and repetition on chunking of digit strings. Psychol Res 68:216–223
Frensch PA, Haider H, Ruenger D et al (2002) The route from implicit learning to verbal expression of what has been learned: verbal report of incidentally experienced environmental regularity. John Benjamins, New York
Ghilardi MF, Moisello C, Silvestri G et al (2009) Learning of a sequential motor skill comprises explicit and implicit components that consolidate differently. J Neurophysiol 101:2218–2229
Hager-Ross C, Schieber MH (2000) Quantifying the independence of human finger movements: comparisons of digits, hands, and movement frequencies. J Neurosci 20:8542–8550
Heyes CM, Foster CL (2002) Motor learning by observation: evidence from a serial reaction time task. Q J Exp Psychol A 55:593–607
Hikosaka O, Nakamura K, Sakai K, Nakahara H (2002) Central mechanisms of motor skill learning. Curr Opin Neurobiol 12:217–222
Honda M, Deiber MP, Ibáñez V et al (1998) Dynamic cortical involvement in implicit and explicit motor sequence learning. A PET study. Brain 121:2159–2173
Hosmer DW, Lemeshow S (2000) Applied logistic regression. Wiley, Hoboken, NJ, USA
Howard D, Howard J (2001) When it does hurt to try: adult age differences in the effects of instructions on implicit pattern learning. Psychon Bull Rev 8:798–805
Hund-Georgiadis M, Yves Von Cramon D (1999) Motor-learning-related changes in piano players and non-musicians revealed by functional magnetic-resonance signals. Exp Brain Res 125:417–425
Keele SW, Jennings PJ (1992) Attention in the representation of sequence: experiment and theory. Hum Mov Sci 11:125–138
Kendall MG, Smith BB (1939) The problem of m rankings. Ann Math Stat 10:275–287
Koch I (2007) Anticipatory response control in motor sequence learning: evidence from stimulus–response compatibility. Hum Mov Sci 26:257–274
Koch I, Hoffmann J (2000) The role of stimulus-based and response-based spatial information in sequence learning. J Exp Psychol Learn Mem Cogn 26:863–882
Loehr JD, Palmer C (2007) Cognitive and biomechanical influences in pianists’ finger tapping. Exp Brain Res 178:518–528
Loehr J, Palmer C (2009) Sequential and biomechanical factors constrain timing and motion in tapping. J Mot Behav 41:128–136
Moisello C, Avanzino L, Tacchino A et al (2011) Motor sequence learning: acquisition of explicit knowledge is concomitant to changes in motor strategy of finger opposition movements. Brain Res Bull 85:104–108
Nissen MJ, Bullemer P (1987) Attentional requirements of learning: evidence from performance measures. Cogn Psychol 19:1–32
Norman DA, Shallice T (1986) Attention to action. In: Consciousness and self-regulation. Springer, US, Boston, MA, pp 1–18
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113
Ollen J (2006) A criterion-related validity test of selected indicators of musical sophistication using expert ratings. The Ohio State University, Ohio
Parsons LM, Sergent J, Hodges DA, Fox PT (2005) The brain basis of piano performance. Neuropsychologia 43:199–215
Pascual-Leone A, Grafman J, Hallett M (1994) Modulation of cortical motor output maps during development of implicit and explicit knowledge. Science 263:1287–1289
Povel DJ, Collard R (1982) Structural factors in patterned finger tapping. Acta Psychol (Amst) 52:107–123
Rosenbaum D, Kenny S, Derr M (1983) Hierarchical control of rapid movement sequences. J Exp Psychol Hum Percept Perform 9:86–102
Rünger D, Frensch P (2008) How incidental sequence learning creates reportable knowledge: the role of unexpected events. J Exp Psychol Learn Mem Cogn 34:1011–1026
Sakai K, Kitaguchi K, Hikosaka O (2003) Chunking during human visuomotor sequence learning. Exp Brain Res 152:229–242
Singer R, Lidor R, Cauraugh J (1993) To be aware or not aware? What to think about while learning and performing a motor skill. Sport Psychol 7:19–30
Verleger R, Seitz A, Yordanova J, Kolev V (2015) Is insight a godsend? Explicit knowledge in the serial response-time task has precursors in EEG potentials already at task onset. Neurobiol Learn Mem 125:24–35
Verwey WB (2003) Processing modes and parallel processors in producing familiar keying sequences. Psychol Res 67:106–122
Verwey WB (2015) Contributions from associative and explicit sequence knowledge to the execution of discrete keying sequences. Acta Psychol (Amst) 157:122–130
Verwey WB, Abrahamse EL, de Kleine E (2010) Cognitive processing in new and practiced discrete keying sequences. Front Psychol 1:1–13
Verwey WB, Abrahamse EL, Ruitenberg MFL et al (2011) Motor skill learning in the middle-aged: limited development of motor chunks and explicit sequence knowledge. Psychol Res 75:406–422
Verwey WB, Shea CH, Wright DL (2014) A cognitive framework for explaining serial processing and sequence execution strategies. Psychon Bull Rev 22:54–77
Willingham DB (1999) Implicit motor sequence learning is not purely perceptual. Mem Cognit 27:561–572
Willingham DDB, Nissen MJ, Bullemer P (1989) On the development of procedural knowledge. J Exp Psychol Learn Mem Cogn 15:1047–1060
Wong AL, Lindquist MA, Haith AM, Krakauer JW (2015) Explicit knowledge enhances motor vigor and performance: motivation versus practice in sequence tasks. J Neurophysiol 114:219–232
Wulf G, Prinz W (2001) Directing attention to movement effects enhances learning: a review. Psychon Bull Rev 8:648–660
Yordanova J, Kirov R, Kolev V (2015) Increased performance variability as a marker of implicit/explicit interactions in knowledge awareness. Front Psychol 6:1–11
Acknowledgments
Funding was supported by F31NS087835 to MJ, University of Rochester Provost Multidisciplinary Award, Geoffrey Waasdorp Pediatric Neurology Fund, and the University of Rochester CTSI. We thank Gil Rivlis, PhD for computer programming assistance and Martin Gira for electronic shop assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jaynes, M.J., Schieber, M.H. & Mink, J.W. Temporal and kinematic consistency predict sequence awareness. Exp Brain Res 234, 3025–3036 (2016). https://doi.org/10.1007/s00221-016-4704-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-016-4704-7