
Abstract
In two experiments bimanual movements with various combinations of target directions were studied by means of the timed-response procedure. The findings revealed an adaptive modulation of intermanual interactions during direction specifications depending on particular target directions. For symmetric movements intermanual correlations of movement directions are positive, indicating a symmetric coupling. For parallel movements the positive intermanual correlations, observed at short preparation intervals, turn into negative correlations as the time available for motor preparation increases. Biases of mean directions, that can be observed for movements to targets with different eccentricities, reflect one or the other kind of coupling, symmetrical for symmetric target directions and parallel for parallel target directions. These biases are static, that is, they are present at long preparation times, and they are phasically enhanced at shorter preparation intervals. The task-adaptive modulation of intermanual interactions is superposed on a basic symmetry bias.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bimanual movements cannot always be performed as intended. Concurrent movements of the left and right hand, which differ in a certain characteristic, often become more similar than they should be. This kind of intermanual interaction can be conceived as coupling which results from the cross-talk between signals involved in the control of the movements (cf. Spijkers and Heuer 2004). The present experiments have been designed to examine certain hypotheses on the coupling with respect to movement directions.
There is considerable evidence that aiming movements are specified in terms of amplitude and direction (e.g., Gordon et al. 1994a, b; Krakauer et al. 2004; Rossetti et al. 1995; Vindras and Viviani 2002). While intermanual coupling with respect to amplitudes has been studied in some depth during the last two decades, directional coupling has received rather limited attention. However, there are at least two reasons to expect that the characteristics of directional coupling and its modulation during motor preparation may not be the same as those of amplitude coupling and its modulation.
First, the anatomical substrates of intermanual interactions with respect to various movement characteristics are likely to be different. Direction and amplitude are coded differently at the level of single cells and cell populations (Fu et al. 1993, 1995). There is also evidence of different brain structures being involved in the specification of these two movement characteristics (e.g., Desmurget et al. 2003; Krakauer et al. 2004). Finally, Wenderoth et al. (2005) specifically showed that intermanual amplitude interference activates not only a parietal–premotor network, which is also activated by intermanual direction interference, but also prefrontal areas.
Second, there is a perhaps rather fundamental difference between directions and amplitudes in that “same” amplitudes are clearly defined, but “same” directions are not. Rather “same” directions can be defined in terms of different reference systems, namely with reference to the symmetry of the body or with reference to the external world. Therefore “same” can refer to symmetric or parallel movements of the two hands, respectively. Thus, whereas there are same and different amplitudes, there are symmetric, parallel, and different (neither symmetric nor parallel) directions. This suggests that intermanual interactions with respect to directions might exhibit a more complex pattern behaviorally than intermanual interactions with respect to amplitudes.
The systematic study of intermanual interactions with respect to movement directions has begun only recently. Swinnen et al. (2001, 2002, 2003; Wenderoth et al. 2004) used a task in which lines with vertical orientation were drawn repetitively with the left hand and lines with variable orientations with the right hand. The orientation of the right-hand lines had to be shifted by 45° clockwise every five cycles. The variable orientations of right-hand movements induced systematic biases in the orientation of left-hand movements.
Swinnen and co-workers took some steps to determine the nature of signals that are critical for directional coupling. Intermanual interactions are generally conceived as resulting from the cross-talk between signals involved in the advance specification and the control of left-hand and right-hand movements. Different types of signals have been claimed to be critical, in particular outflow signals (e.g., Cattaert et al. 1999; Preilowski 1975), sensory signals (e.g., Mechsner et al. 2001) or signals involved in the specification of movement characteristics (Heuer 1993; Spijkers and Heuer 1995). When the observed directions were dissociated from the actual movement directions, Swinnen et al. (2003) found the cross-manual directional effects to be determined by the actual movement directions. Thus, sensory signals (at least visual ones) are not critical. Swinnen et al. (2001) varied the forces necessary to produce the movements. The cross-manual directional effects turned out to be essentially independent of the forces. Thus, they are not critically dependent on outflow signals. Taken together, these findings suggest a critical role of cross-talk during the specification of movement directions.
The present experiments have been designed to study the concurrent specifications of the directions of bimanual movements in more detail. In particular we analyze whether directional coupling is modulated in the course of motor preparation and whether the rates of specifying symmetric, parallel, and different directions differ. We shall first describe a method which is suited to answer these questions and then some concepts and preliminary data which suggest certain expectations for the answers.
A methodology which allows the study of the time course of specifying movement characteristics like amplitude and direction is the timed-response procedure (Hening et al. 1988; Ghez et al. 1997). Basically, participants have to initiate movements in synchrony with a predictable pacing signal. At a variable time before the pacing signal cues are presented which indicate certain characteristics of the movements, e.g., their amplitudes. Thus, the time available for movement specification is controlled. With the passage of preparation time, amplitudes or other characteristics of the movements produced gradually change from an instructed or spontaneously chosen default value to the cued target value.
The timed-response procedure allows rather direct tests of the notion that at least certain kinds of intermanual interactions result from the cross-talk during movement specifications (Spijkers and Heuer 1995; cf. Heuer 1990; Spijkers and Heuer 2004, for overviews). Cross-talk during movement specification in principle can be static; in this case different movement characteristics cannot be specified concurrently, but there will be interaction effects no matter how much time is available for motor preparation. However, cross-talk during movement specification can also be phasic or transient; in this case interaction effects will be seen only at short preparation intervals, but not at long ones. Transient cross-talk implies a de-coupling of the concurrent processes of specifying left-hand and right-hand movement characteristics in the course of motor preparation.
Heuer et al. (1998), (2001) used the timed-response procedure to trace the time course of amplitude specifications. Figure 1 shows typical results, taken from Heuer et al. (2001), which illustrate evidence of transient cross-talk and the associated de-coupling in the course of motor preparation. These results are taken from a condition in which the differences between target amplitudes were sufficiently small for the often found assimilation effects at long preparation intervals (e.g., Marteniuk et al. 1984; Sherwood 1990, 1991, 1994a, 1994b; Sherwood and Nishimura, 1992) to be absent. On the abscissae of the two graphs of Fig. 1 the preparation interval is shown, operationally the interval between the presentation of the cues and the mean initiation times of the movements of the two hands. For the case of very short cueing intervals participants were instructed to prepare for intermediate amplitudes. Thus, at the shortest cue-response intervals (CRIs) amplitudes were intermediate and independent of the cues.

a Time course of amplitude specifications for bimanual movements with same and different amplitudes. The CRI is the time from cueing the target amplitudes until the start of the movements. b Time course of intermanual amplitude correlations during the specification of same and different amplitudes (after Heuer et al. 2001)
Figure 1a shows the gradual specification of short and long amplitudes when the CRI becomes longer. More importantly, the rate of specification depends on whether the same or a different amplitude is specified for the other hand concurrently. With different amplitudes specification lags behind initially, so that long-amplitude movements are shorter and short-amplitude movements are longer than with same amplitudes. This is a transient assimilation which can be seen only at short preparation intervals. It disappears in the course of motor preparation even though the difference between the amplitudes actually produced increases.
In Fig. 1b the intermanual amplitude correlations are shown. These are the correlations between the left-hand and right-hand amplitudes computed across a set of trials with a certain range of CRIs. They capture the relation between the random variations of left-hand and right-hand amplitudes. Formally they are independent of the assimilation effects, which relate to the mean amplitudes, but functionally this is unlikely to be the case. At short CRIs the intermanual amplitude correlations are high, and when same amplitudes are specified they remain high at long CRIs. However, when different amplitudes are specified they decline, so that amplitude variation of the left hand becomes almost independent of the amplitude variation of the right hand. This is an indication of de-coupling which is claimed to be responsible for the transient nature of assimilation effects.
The differences in the anatomical substrates of amplitude and direction specifications and of the intermanual cross-talk related to these movement characteristics suggest that the behavioral manifestations of intermanual cross-talk may be different as well. However, they do not suggest specific differences. Given the general symmetry constraints on bimanual movements (see Heuer 1996, for an overview), it is likely that there is a basic symmetric coupling with respect to movement directions. It may be relaxed when different directions are specified concurrently, similar to how amplitude coupling is relaxed in the course of specifying different amplitudes. Of course, this is a hypothesis which is based only on the assumption that intermanual interactions during motor specification are basically of the same kind as far as amplitudes and directions are concerned; even though this is quite a weak basis, the hypothesis can serve as a reference for deviant findings.
What are the expectations for the concurrent specification of parallel directions? Again a general symmetry constraint could be relaxed. However, there is also the possibility that there is a strong modulation of directional coupling during motor preparation in that there is not only a de-coupling, but also a re-coupling which is parallel instead of symmetric. In this case intermanual direction correlations would not only approach zero as the preparation interval increases, but would also change sign. Preliminary observations of such changes have been made by Steglich (2002) for rapid bimanual aiming movements with symmetric and parallel directions.
Experiment 1
Our primary interest in the first experiment was in the intermanual correlations of movement directions and their changes during motor preparation. Modulations of intermanual correlations in the course of direction specifications should be different for symmetric, parallel, and different directions. Assuming an initial symmetric coupling of movement directions, intermanual direction correlations should decline in the course of specifying different directions (comparable to the decline of intermanual amplitude correlations in the course of specifying different amplitudes). More importantly, intermanual direction correlations should switch sign in the course of specifying parallel directions, indicating a switch from symmetric to parallel coupling.
In addition to the intermanual correlations we were interested in the second indicator of intermanual directional coupling, namely assimilation effects. Assimilation effects which show up at short preparation intervals might disappear at longer preparation intervals (transient or phasic effects), but they can also remain (static effects). Static effects at long preparation intervals would be expected from the findings of Swinnen et al. (2001). However, the assimilation effects observed in repetitive-movement tasks are not necessarily the same as those observed in discrete-movement tasks (Heuer and Klein 2005).
In the first experiment we studied the specification of symmetric, parallel, and different directions of rapid bimanual reversal movements. Rapid reversal movements had been used in previous studies on the specification of bimanual amplitudes (Heuer et al. 1998, 2001) because for them the measurement of amplitudes is more straightforward than for rapid aiming movements. Thus far there are no reasons to question the tacit assumption that intermanual interactions are essentially the same for rapid reversal movements and rapid aiming movements. Therefore we continued to use rapid reversal movements in the present experiments.
In the studies of directional coupling Swinnen et al. (2001) had used differences between the left-hand and right-hand directions of 45° and multiples thereof. In contrast, we used differences between left-hand and right-hand directions of only 0°, 30°, and 60°. Specifically, at the start of each trial participants were instructed to prepare for the production of forward movements, and the cues asked for forward movements or directions that deviated laterally or medially by only 30°. The reason for choosing such a restricted range of directions is a finding of Ghez et al. (1997). They found a continuous specification of movement directions only when the alternative directions were not too different. Otherwise direction specification was discrete, so that only the one or the other of the alternative directions was specified in the course of motor preparation, but not the intermediate ones. The different modes of direction specification could be associated with different intermanual interactions. The restricted range of movement directions in the present experiments therefore serves to secure a continuous direction specification, comparable to the continuous specification of amplitudes in previous experiments.
Method
Participants
Fourteen participants, 2 females and 12 males, aged 22–35 years (mean: 25.6 years, SD: 3.8 years), took part in the experiment. All participants were right-handed. Three additional participants were unable to follow the instruction to produce straight rapid reversal movements in synchrony with a pacing tone. Participants had given informed consent prior to the start of the experiment. The present study was done with the approval of the institutional ethics committee and in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
Apparatus
Participants sat on a height-adjustable chair facing a table. A 15-inch LCD monitor (EIZO Flex Scan L365) was placed on the table platform with the screen at a distance of about 100 cm from the eyes of the participants. Both their index fingers were strapped to slides of 50×30 mm2 (height: 6 mm) which ran with only little friction on a glass plate. The slides carried sensors of a miniBIRD system (Ascension Technology Corporation, miniBIRD 800), which were vertically oriented and located above the finger nails. The transmitter was located 20 cm below the table top, laterally centered relative to the work space and 10 cm nearer to the participant than the start positions of the bilateral reversal movements. Positions of the finger tips were recorded at 120 Hz (static spatial resolution: 0.5 mm). An occluder 20 cm above the table platform prevented vision of the hands. A loudspeaker was located at the wall directly behind the participants.
Task, experimental conditions, and procedure
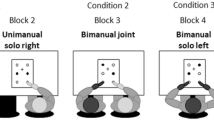
Participants were to produce rapid straight bimanual reversal movements to targets indicated on the monitor, beginning in synchrony with the fourth (and last) of a regular series of tones. On the monitor the start-target configurations for the two hands were shown (Fig. 2). Targets were circles of 7 mm diameter, the center-to-center distances from the start circles to target circles were 71 mm, and the lateral distance between start positions was 120 mm. Overall the start-target configuration was of width 10.5° and height 4.5°. On the table the lateral distance between the start positions was 40 cm (their distance from the table edge facing the participants was 12 cm). Target amplitudes were 15 cm. Targets for each hand were located in the forward direction (0°), laterally displaced (−30°), or medially displaced (+30°).

Display of start-target configurations for the two hands; hand positions were indicated as small red circles (shown as gray in the Figure) as long as the start positions were not yet reached (left graph). Targets were cued by way of filling the appropriate circles (right graph)
For each trial participants were first to prepare bimanual forward movements. At a variable time before the requested movement initiation (cueing interval) one of the nine pairs of targets was marked as the correct one by way of filling the corresponding circles on the screen (cf. Fig. 2, right graph). In total there were 72 different conditions, obtained by crossing three experimental factors: target direction of the left hand (−30°, 0°, +30°), target direction of the right hand (−30°, 0°, +30°), and cueing interval (0, 150, 250, 325, 400, 475, 575, and 700 ms).
At the start of each trial participants had to move the index fingers to the start positions. For that purpose their finger positions were indicated on the screen by two filled red circles of 3 mm diameter (cf. Fig. 2, left graph). As soon as each finger reached its start position, the respective red circle turned green. When both fingers were in the start positions, the filled red circles which marked the finger positions disappeared and the two forward targets were filled for 1 s. This was to remind participants to prepare forward movements initially. After the second had elapsed, a sequence of four tones, 1,000, 1,260, 1,500, and 2,000 Hz, was presented, with inter-stimulus intervals of 500 ms and tone durations of 20 ms. At a variable cueing interval of 700, 575, 475, 400, 325, 250, 150 or 0 ms before the fourth tone the targets for that trial were cued by way of filling the respective circles on the screen. The cues remained on for 1,000 ms after presentation of the fourth tone. Thereafter feedback was presented on the monitor, which defined rather strict performance criteria except for accuracy. These performance criteria were also emphasized in the instruction.
For valid trials "ok" was shown. For valid trials with deviations of movement initiations from the fourth tone in the range of 50–200 ms, these deviations were indicated in addition. For invalid trials the reasons were specified which could be: (1) deviation of movement initiation from the fourth tone longer than 200 ms, (2) asynchrony of the left-hand and right-hand movement initiations longer than 100 ms, (3) movement duration (until reversal) longer than 250 ms, (4) movement amplitude less than 100 mm, (5) very strong curvature. To assess curvature, for each measured finger position its orthogonal deviation from the straight line connecting the start and end position of each movement was determined; movements were defined as invalid when the maximum deviation was larger than 12 mm and the mean deviation larger than 8 mm. The criteria for movement duration and curvature in particular served to prevent a strategy with which movements are started forward, rather independently of the cued direction, and then corrected during execution to reach the indicated target. Such a strategy is difficult with really rapid movements in principle, and it results in curved movements which did not pass the performance criteria.
Each block consisted of 72 experimental trials and two initial warm-up trials. To each of the experimental trials one of the 72 experimental conditions was assigned randomly. In total there were 18 blocks of trials, four on day 1, seven on day 2, and seven on day 3 of the experiment. The experimental blocks on day 1 were preceded by a practice period of about 30 min duration. During this period rapid reversal movements as well as the synchronization of movement initiations with the fourth tone were practiced. Experimental blocks on days 2 and 3 were preceded by 18-trial warm-up blocks.
Data analysis
The time series of x- and y-coordinates of each finger tip were low-pass filtered (fourth order Butterworth, 10 Hz, forward and backward pass). From the distances between successive positions the tangential-velocity signals were derived (two-point central difference algorithm) and low-pass filtered again. For each movement the reversal position was determined as the position with the largest Euclidean distance from the start position. Moving backward from the position of the peak tangential velocity, the start of the movement was identified as the first point at which the distance between successive sampled positions was less than 3 mm. Movement amplitudes and directions were derived from the vectors pointing from the start positions to the reversal positions of the two movements in each trial. In addition to the terminal direction the initial direction of each movement was identified. Initial direction was the direction of the vector pointing from the start position of the movement to the earliest position at which the distance from the start position exceeded 20 mm.
As an illustration of typical individual data, Fig. 3 shows the movement directions of the first participant in symmetric-movement conditions. For each movement with the left or right hand the initial direction is shown as a function of the CRI, which is the time from the presentation of the cues to movement initiation. For CRIs up to about 200 ms initial directions varied randomly around the forward directions (default directions of 0°). For CRIs longer than 400–500 ms the cued directions were reached which were close to the target directions of −30° (lateral) and +30° (medial). For CRIs longer than 200 ms and shorter than 400–500 ms, movement directions gradually shifted from the default direction to the target direction.

Individual data in the symmetric-movement condition. For each movement the initial direction is shown as a function of the interval between the cue presentation and the start of the movement. Filled circles represent movements to medial targets, open circles represent movements to lateral targets
For the statistical analysis the trials were sorted into bins according to the CRI. As compared with an analysis in terms of the cueing intervals, this procedure has the advantage that for each data point the variability of the CRIs is smaller (without omitting trials in which movement initiations deviated too strongly from the fourth tone); in addition the means associated with each bin capture the CRIs more accurately than the cueing intervals (cf. Heuer et al. 1998). For the sorting of trials into bins the CRI was defined for each individual trial (rather than for each individual movement as in Fig. 3). Specifically, the CRI of a trial was the mean of the two CRIs for the left-hand and the right-hand movement in that trial. Table 1 gives the boundaries of the bins as well as the mean number of trials in each bin. In brackets the ranges across the nine combinations of movement directions are given. In the following all conditions are referred to the mean CRI of each bin.
For each participant, combination of target directions, and CRI bin the intermanual correlations were computed, that is, the correlations between both the initial and terminal directions of left-hand and right-hand movements. For each hand lateral directions were given a negative sign and medial directions a positive sign, so that positive correlations indicate a symmetric relation between directional variations of the two hands and negative correlations a parallel relation. For all further analyses of these correlations Fisher’s z-transforms were used, but the presented results are again correlation coefficients obtained by inverse transformations of the mean z-transforms. In addition the means of initial direction, terminal direction, amplitude, and movement time were determined for each hand. Amplitude and movement time were examined mainly to be able to detect eventual variations which could have affected the findings on directions.
Results
Overall the mean amplitude of the rapid reversal movements produced was 152 mm, and the mean movement time (until reversal) was 198 ms. In reporting the results, we focus on the initial movement directions and do not report the findings for terminal directions. Initial directions are less affected than terminal directions by biomechanical factors and by feedback-based corrections and/or by the continued specification of directions during movement execution. In addition, the findings for the initial and terminal directions were quite similar. There were only two major differences which are not important for the purpose of the present study.
The first difference was that the overall terminal directions were shifted laterally as compared with initial directions, indicating a slight but systematic curvature of the movements. The second difference was that the intermanual correlations of terminal directions were more extreme than the intermanual correlations of initial directions. The first of these differences is likely due to biomechanical factors. The lateral shift of directions during the movements is accompanied by a reduction of inertia which is higher for movements in more medial directions than for movements in more lateral directions. The second of these differences is likely due to the fact that measurement noise resulting from limited spatial and numerical resolution has a stronger effect on initial than on terminal directions.Footnote 1
Intermanual correlations
The various target combinations were grouped into four conditions: forward movements (target combination 0°/0°), symmetric movements (target combinations −30°/−30°, +30°/+30°), parallel movements (target combinations −30°/+30°, +30°/−30°), and different movements (target combinations −30°/0°, +30°/0°, 0°/−30°, 0°/+30°). For each participant the z-transforms were averaged across the respective target combinations, and these averaged z-transforms were subjected to statistical analysis. The mean correlations as a function of the CRI are shown in Fig. 4.

Intermanual correlations of initial directions as a function of the CRI for bimanual movements with parallel, symmetric, forward, and different directions (Experiment 1)
For all conditions the intermanual correlations were positive at the shortest CRIs. Positive correlations indicate a symmetric coupling so that a left-hand movement directed more laterally tends to go along with a right-hand movement directed more laterally as well. For symmetric movements the correlations remained rather stable at the longer CRIs, for forward movements there was a slight decline, and for movements with different directions there was a somewhat stronger decline. In contrast, for parallel movements the correlations at CRIs longer than about 300 ms were negative and indicated a parallel coupling, so that a left-hand movement directed more laterally tended to go along with a right-hand movement directed more medially.
The statistical analysis consisted of sets of planned contrasts based on a two-factorial design with four relations between movement directions (forward, different, symmetric, parallel) and seven CRIs. The contrasts dealt with the initial state of directional coupling, its change during motor preparation, and its final state at long CRIs in turn. The first set of contrasts was concerned only with the shortest CRI, that is, the initial state of directional coupling. The differences between the four relations were not reliable, F<1, and the mean correlation, computed across all four relations between target directions, was significantly different from zero, F(1, 13)=28.4, P<0.01. Thus, initially there was a symmetric coupling that was independent of the relation between target directions.
The second set of contrasts was concerned with the changes of the correlations across CRIs. We considered the condition with forward movements as a baseline condition because the cued directions were identical to the default directions. Thus, during the CRI no change of the specified directions was required. The slight decline of the intermanual direction correlations was not significant, F(6, 78)=1.3, P>0.20. We compared the change in each of the other conditions with the change in the baseline condition. With different directions the change across CRIs was not significantly different from the one observed with forward movements, F(6, 78)=1.4, P>0.20, and with symmetric movements there was an almost significant difference from the baseline change, F(6, 78)=1.9, P<0.10. Only with parallel movements the change across CRIs was clearly different from the one observed in the baseline condition, F(6, 78)=5.8, P<0.01.
The third set of contrasts was concerned with the longest CRI, that is, the final state of directional coupling at the end of motor preparation. First, we tested whether for the baseline condition with forward movements the correlations were significantly different from zero, which they were, F(1, 13)=7.5, P<0.05. Second, for each of the other combinations of target directions we tested the deviation from the baseline condition. This was significant for symmetric movements, F(1, 13)=22.4, P<0.01, and for parallel movements, F(1, 13)=8.6, P<0.05, but not for movements with different directions, F(1, 13)=1.3, P>0.10. (Both for symmetric and parallel movements the correlations were also significantly different from zero, F(1, 13)=39.7, P<0.01, and F(1, 13)=5.1, P<0.05, respectively). Finally we tested the mean of the correlations observed with symmetric and parallel movements against zero. This contrast is equivalent to a comparison of the absolute correlations in these two conditions, and it was significant, F(1, 13)=14.5, P<0.01. Thus, at the end of motor preparation the symmetric coupling observed with symmetric movements was stronger than the parallel coupling observed with parallel movements.
Mean initial directions at long CRIs
The mean initial directions at the longest CRI give evidence of static assimilation effects. The mean initial directions of movements to eccentric targets are shown in Fig. 5. In these analyses all target directions were designated as +30°, so that movements in lateral and medial directions could be meaningfully compared. For each hand and target direction—with the exception of left-hand movements to lateral targets—the movement direction was the least eccentric when a different target direction was assigned to the other hand, that is, when the other hand performed forward movements. The data were subjected to a three-way ANOVA with the within-participant factors Relation (symmetric, parallel, different), Target Direction (lateral, medial), and Hand (right, left). The triple interaction was significant, F(2, 26)=3.5, P<0.05, and so were the Relation×Hand interaction, F(2, 26)=5.7, P<0.01, and the Relation×Target Direction interaction, F(2, 26)=6.0, P<0.01. Finally there was a significant main effect of Target Direction, F(1, 13)=6.4, P<0.05, in that movements to lateral targets were more eccentric than movements to medial targets.

Asymptotic initial directions at the longest CRI of movements of the left and right hand to lateral and medial targets as a function of the relation between the target directions (symmetric, parallel, and different). (Movements in the direction of the target are marked by positive values.)
Mean normalized initial directions
Time courses of direction specifications give evidence of intermanual interactions during motor preparation. For example, they can reveal transient assimilation effects that can be observed only at short preparation intervals, but not at long ones (cf. Fig. 1a). However, the comparison of the time courses can be plagued by differences observed at long preparation intervals. For example, when two conditions are compared in which movement directions are different at long preparation intervals, this difference will gradually develop in the course of motor preparation and result in different time courses which are a consequence of the different asymptotic directions only. To unconfound differences between time courses from differences between asymptotic directions, we analyzed normalized directions. In particular, for each participant, relation between directions, target direction, and hand, the deviation of the direction at each CRI from the direction at the shortest CRI was expressed as a proportion of the deviation of the direction at the longest CRI from the direction at the shortest CRI. Thus, for the shortest CRI normalized directions were 0 by definition, and for the longest CRI they were 1 by definition.
In Fig. 6 the normalized initial directions are shown. Beginning about 200 ms after the presentation of the cues, the initial directions started to shift away from the default (forward) directions to the final directions of about 30° eccentricity. For movements to medial targets the time courses of direction specifications were essentially the same for symmetric, parallel, and different target directions. In contrast, for movements to lateral targets the time courses were different. In particular, symmetric directions were specified at a faster rate than parallel and different directions. More exactly, lateral directions were specified at a faster rate when symmetric (lateral) directions for the other hand were specified concurrently than when parallel (medial) or different (forward) directions for the other hand were specified.

Normalized initial directions of movements to lateral and medial targets as a function of the CRI and the relation between the target directions (symmetric, parallel, and different; Experiment 1)
The normalized initial directions were subjected to a series of planned contrasts (neglecting the first and last CRI for which all normalized directions are 0 and 1 by definition). We chose the condition with symmetric movements as a reference condition with which we compared the conditions with parallel and different movements. With different target directions, normalized initial directions were less eccentric (smaller) overall than with symmetric target directions, F(1, 13)=4.7, P<0.05. The interaction of different vs. symmetric directions with CRI was significant in addition, F(4, 52)=2.8, P<0.05, and so was the triple interaction of different vs. symmetric directions, CRI, and Target Direction, F(4, 52)=3.1, P<0.05. Thus, the time course of direction specifications was different for symmetric and different directions, but only for lateral and not for medial targets. In contrast, the comparison of conditions with symmetric and parallel target directions revealed that the differences which are apparent from Fig. 6 were not statistically significant.
Mean amplitude and movement time
Mean amplitudes and movement times were of interest only with respect to possible variations which could have critically affected the findings on intermanual correlations and mean initial directions. However, there are no obvious confounds of this kind. The mean amplitudes and movement times are shown in Fig. 7. They are plotted in terms of the direction of the other hand (forward, medial, lateral) rather than in terms of the relation between directions (different, symmetric, parallel) because this simplifies the pattern of results.

Mean amplitude and movement time (MT) of the movements to lateral and medial targets as a function of the CRI, shown separately for different directions (lateral, forward, medial) of the other hand
At the three shortest CRIs, when essentially forward movements were produced and intermanual correlations were hardly affected by the different combinations of target directions, mean amplitudes and movement times were also independent of the target direction cued for the other hand. At the longer CRIs both amplitudes and movement times increased, more so for movements to medial targets than for movements to lateral targets. The increase was graded according to the target direction of the other hand, being strongest with a lateral direction of the other hand and weakest with a medial direction. (Only for the movement time of movements to lateral targets was the increase across CRIs quite weak and the gradation according to the target direction for the other hand was absent.) The statistical analysis revealed that all these variations were reliable, but for the sake of brevity and simplicity the details are not reported.
Discussion
The primary purpose of the first experiment was to compare the changes of intermanual direction correlations while different combinations of bimanual movement directions are specified. At short preparation intervals, when forward movements have been prepared as default and are actually produced, intermanual direction correlations are positive, no matter whether symmetric, parallel or different target directions are cued. As compared to a baseline condition in which forward movements are cued, intermanual direction correlations tend to become even more strongly positive when symmetric directions are cued. In contrast, when parallel directions are cued, positive correlations turn into negative ones at CRIs of about 300 ms. The switch from positive to negative correlations indicates a switch from symmetric to parallel coupling and confirms preliminary observations of Steglich (2002). The modulations of directional coupling in the course of specifying symmetric and parallel movements are adaptive with respect to task requirements, in that the specification of symmetric movements is supported by a symmetric coupling and the specification of parallel movements by a parallel coupling.
Even though the modulations of directional coupling during motor preparation are adaptive, there is a basic symmetry bias in addition. This is revealed by two observations. First, for forward movements, which are both symmetric and parallel, the directional coupling is symmetric (positive intermanual direction correlation). Second, at the longest CRI the positive intermanual correlations with symmetric directions are stronger than the negative correlations found with parallel directions. Thus, the adaptive modulation of directional coupling is superposed on a basic symmetric coupling which is present independently of the task requirements.
Whereas directional coupling is adapted to task requirements when symmetric and parallel movements are specified, there seems to be no adaptive modulation when target directions are different. As compared to the baseline condition in which forward movements are cued, with different target directions there is no reliable decline of intermanual direction correlations which would indicate a de-coupling and thus represent an adaptive modulation of intermanual correlations. This finding contrasts sharply with the observations made for the concurrent specification of same and different amplitudes of bimanual reversal movements (Heuer et al. 1998, 2001), and same and different peak forces of bimanual isometric contractions (Steglich et al. 1999). Perhaps the unexpected finding is a chance result or is related to the fact that with different target directions one of them was always the default direction in this experiment, so that after presentation of the cues only one direction had to be re-specified.
Turning to the time courses of direction specifications, the present results reveal static assimilation effects at long CRIs. These are likely to be analogous to the cross-manual directional effects observed by Swinnen et al. (2001, 2002, 2003; Wenderoth et al. 2004) in a repetitive-movement rather than discrete-movement paradigm. Specifically, when target directions are different, the movements to the eccentric targets are less eccentric than when target directions are symmetric or parallel (with the exception of left-hand movements to the lateral target). In contrast, the eccentricity of movement directions does not depend on whether target directions are symmetric or parallel. This observation is consistent with the conclusion based on intermanual correlations that directional coupling is adapted to task requirements. Thus, the specification of parallel directions is not opposed by a symmetric coupling which could have produced a kind of “symmetric assimilation” of parallel movements, resulting in a reduced eccentricity of the movement directions of both hands.
In addition to static assimilation effects, the time courses of direction specifications reveal some differences between bimanual movements with symmetric, parallel, and different directions. Basically, with different and parallel target directions the specification of movement directions is slowed down as compared to symmetric target directions. However, this difference is asymmetric: it is found for movements to lateral targets, but not for movements to medial targets. Even though the statistical status of this pattern of results is ambiguous, it is consistent with preliminary observations of Steglich (2002).
The details of the different time courses of specifying symmetric, parallel, and different movement directions do not seem to conform to the assumption that they result from a transient (or phasic) coupling of movement directions. In the studies of bimanual amplitude specifications (Heuer et al. 1998, 2001) the differences of the time courses of specifying same and different amplitudes were of a certain kind. With different amplitudes the amplitude change with increasing CRI was slowed down at the shortest CRIs and made it up to the amplitudes specified with the same target amplitudes at longer CRIs (cf. Fig. 1a). In this experiment, in contrast, the specification of parallel and different directions was not delayed relative to the specification of symmetric directions from the very start, but the delay showed up at a longer CRI only.
The variations of mean amplitude and mean movement time turned out to be essentially unrelated to the other results and to reflect other influences. For example, movements in the medial direction are somewhat slower than movements in the lateral direction, which conforms to the observation that the differences in inertia between the different movement directions are not fully compensated in general (Gordon et al. 1994; Levin et al. 2001).
Experiment 2
The second experiment was intended to replicate the main findings of Experiment 1, in particular the unexpected ones like the absence of a reliable difference between the intermanual correlations observed for forward directions and different directions and the statistically ambiguous ones like the different time courses of specifications of symmetric and parallel directions. However, there was also a major change in Experiment 2 in that targets with 15° eccentricity were added. This allows an orthogonal variation of symmetric vs. parallel and same vs. different directions. Thus, in addition to forward movements, there were symmetric same, symmetric different, parallel same, and parallel different movements.
The designation of the combinations of target directions in terms of the contrasts “symmetric–parallel” and “same–different” needs some explanation. The contrast “same–different” refers to the same and different eccentricities of symmetric or parallel movements. Of course, one might hesitate to call symmetric different movements “symmetric” and parallel different movements “parallel”. However, here we use the contrast “symmetric–parallel” for categories of directions, namely lateral and medial directions. Thus, symmetric movements have either lateral or medial directions for both hands, whereas parallel movements have a medial direction for one hand and a lateral direction for the other hand. The designation of target directions in terms of categories and graded differences within categories is pragmatic because it provides simple labels for the conditions of this experiment. However, a hierarchical representation of target directions could also be more than pragmatic and actually correspond to the way they are processed (Huttenlocher et al. 1991; Messier and Kalaska 1999).
Method
Participants
Twelve right-handed participants, five females and seven males, aged 18–29 years (mean: 22.8 years, SD: 4.2 years), took part in the experiment. One additional left-handed participant was not included in the analysis, and five further participants did not complete the experiment because they produced too many errors according to the criterion described below. All participants had given informed consent.
Task, experimental conditions, and procedure
The apparatus was the same as in Experiment 1. So were the basic features of the task: participants had to produce rapid bimanual reversal movements to targets indicated on a monitor, beginning in synchrony with the fourth (and last) of a regular series of tones. The major difference was that for each hand there were five targets, one located in the forward direction (0°), two being laterally displaced (−30°, −15°), and two being medially displaced (+15°, +30°). The horizontal distance between the start positions on the screen was 140 mm. Otherwise the measures of the display were as in Experiment 1, and so was their mapping on the table on which the movements were produced.
From the total of 25 combinations of the five targets for each hand, 17 different combinations were used. These were forward movements (0°/0°), four combinations with symmetric same directions (−30°/−30°, −15°/−15°, +15°/+15°, +30°/+30°), four combinations with parallel same directions (−30°/+30°, −15°/+15°, +15°/−15°, +30°/−30°), four combinations with symmetric different directions (−30°/−15°, −15°/−30°, +15/+30°, +30°/+15°), and four combinations with parallel different directions (−30°/+15°, −15°/+30°, +15°/−30°, +30°/−15°). For each of the 17 target combinations there were the same 8 cueing intervals as in Experiment 1 (0, 150, 250, 325, 400, 475, 575, 700 ms), so that there were 136 experimental conditions in total. Across two successive blocks of trials one trial of each of the 136 experimental conditions was presented. Only forward movements (0°/0°) were requested twice for each cueing interval, so that in two successive blocks there were 144 experimental trials in total, split into two sets of 72 trials each. Each set was preceded by two warm-up trials. Participants came for three sessions on consecutive days. On the first day there were four blocks of trials subsequent to a practice period of about 30 min duration; on each of the following 2 days there were eight blocks of trials, preceded by a short warm-up block.
There were two more procedural changes as compared with Experiment 1. Both were related to movement errors. First, an additional type of error was defined by at least one of the two terminal movement directions in a trial deviating more than 15° from the target direction. With the timed-response procedure directional errors occur necessarily, therefore no feedback on them had been provided in Experiment 1. However, directional errors are inherent to the procedure for short cueing intervals only. Therefore in Experiment 2 the directional-error criterion was applied in trials in which the cueing interval was 575 ms or longer. Second, blocks of trials with more than 15 errors were repeated, and only one repetition per session was allowed. Whenever more than 15 errors were produced in a second block of a session, the experiment was aborted. This happened with five participants.
Data analysis
Data analysis matched the analysis of Experiment 1. For the statistical analyses the trials were sorted into bins according to the CRI. The boundaries of the bins as well as the mean CRIs of all trials in each bin are listed in Table 2. In brackets the ranges of the 17 combinations of movement directions are given. In the following all data are referred to the mean CRIs of the bins. In Table 2 also the mean numbers of trials for each combination of movement directions and bin are given; here the frequencies of forward movements are neglected which are about twice as high as for the other pairs of directions.
Results
Overall the mean amplitude of the rapid reversal movements produced was 158 mm, and the mean movement time (until reversal) was 195 ms. In presenting the results, we focus on the intermanual correlations and the mean initial directions, and do not report the mean amplitudes and mean movement times in detail. For them the findings were essentially the same as in Experiment 1 as far as the effects of the CRI and the medial or lateral directions of the movements and of the movements of the other hand are concerned.
Intermanual correlations
The mean intermanual correlations of initial directions as a function of the CRI are shown in Fig. 8a for the five sets of movement combinations: forward (0°/0°), symmetric same (−30°/−30°, −15°/−15°, +15°/+15°, +30°/+30°), parallel same (−30°/+30°, −15°/+15°, +15°/−15°, +30°/−30°), symmetric different (−30°/−15°, −15°/−30°, +15°/+30°, +30°/−15°), and parallel different (−30°/+15°, −15°/+30°, +15/−30°, +30°/−15°). In Fig. 8b the mean correlations for parallel same and symmetric same movements are shown separately for small eccentricities (−15°/−15°, +15°/+15° and −15°/+15°, +15°/−15°, respectively) and large eccentricities (−30°/−30°, +30°/+30° and −30°/+30°, +30°/−30°, respectively).

Intermanual correlations of initial directions as a function of the CRI for bimanual movements in parallel same, parallel different, symmetric same, symmetric different, and forward directions (a). In b the correlations are shown separately for parallel same and symmetric same directions with low (15°) and high (30°) target eccentricities (Experiment 2)
For forward movements the intermanual correlations of initial directions were positive, indicating a symmetric coupling (cf. Fig. 8a). For symmetric movements the intermanual correlations increased at intermediate CRIs and returned to the short-CRI level at the longest CRIs. The transient increase was more conspicuous than in Experiment 1. For parallel movements, the intermanual correlations again changed sign. At the longest CRI they were negative, but their absolute values were smaller than the positive correlations observed for symmetric movements. At intermediate CRIs the negative correlations were particularly strong. This phasic decrease had been absent in the first experiment. While there was a strong difference between parallel and symmetric movements, the differences between same and different eccentricities for movements of each type were only small. At most there was a slight tendency that intermanual correlations deviated less from zero when directions were different than when they were same.
From Fig. 8b the differences between symmetric and parallel movements with different eccentricities are apparent. Most importantly, the change of sign was not only observed for parallel movements with 30° eccentricity, but also for parallel movements with only 15° eccentricity. In spite of the qualitative similarity of the time courses of the correlations for small and large eccentricities across CRIs, there were quantitative differences. When target eccentricities were large, intermanual correlations tended to be more extreme than when target eccentricities were small, being larger in the positive range and smaller in the negative range.
The statistical analysis of the intermanual direction correlations was carried out in three steps. In the first step the intermanual correlations in conditions with symmetric and parallel target directions of the same eccentricities were subjected to a three-way ANOVA with the within-participant factors Relation of Directions (symmetric, parallel), Eccentricity (small, large), and CRI. These are the data shown in Fig. 8b. The interaction of Relation of Directions with CRI was significant, F(6, 66)=16.5, P<0.01, and so were the main effects of Relation of Directions, F(1, 11)=86.7, P<0.01, and CRI, F(6, 66)=6.9, P<0.01. The changes of the intermanual correlations across the various CRIs were stronger when the eccentricity was high than when it was low, giving rise to a significant triple interaction, F(6, 66)=5.2, P<0.01, as well as to a significant interaction of Relation of Directions and Eccentricity, F(1, 11)=10.7, P<0.01. In addition, at the shortest and longest CRIs, but not at the intermediate ones, intermanual correlations were consistently higher in the high-eccentricity conditions than in the low-eccentricity conditions. The interaction of eccentricity and CRI was significant, F(6, 66)=2.5, P<0.05, and so was the main effect of eccentricity, F(1, 11)=7.9, P<0.05.
In the second step of the statistical analysis conditions with different eccentricities were included. Conditions with small and large same eccentricities were combined so that both the conditions with same and different eccentricities comprised low-eccentricity and high-eccentricity movements (cf. Fig. 8a). A three-way ANOVA with the within-participant factors Relation of Eccentricities (same, different), Relation of Directions (symmetric, parallel), and CRI revealed again a significant Relation of Directions×CRI interaction, F(6, 66)=23.01, P<0.01, as well as significant main effects of Relation of Directions, F(1, 11)=220.2, P<0.01, and CRI, F(6, 66)=8.0, P<0.01. However, the factor Relation of Eccentricities was involved in no significant effect.
In the third step we contrasted the conditions in which symmetric and parallel movement directions were to be specified with the baseline condition in which forward movements had been cued which had also been instructed as the default directions. For the analysis the conditions with same and different eccentricities were collapsed both for symmetric and for parallel movements. The contrast of symmetric and forward movements was significant, F(1, 11)=9.1, P<0.05, and so was its interaction with CRI, F(6, 66)=4.1, P<0.01. Similarly, the contrast of parallel and forward movements was significant, F(1, 11)=105.1, P<0.01, and so was its interaction with CRI, F(6, 66)=7.9, P<0.01.
Mean initial directions at long CRIs
The mean initial directions at the longest CRI are shown in Fig. 9. In general, large eccentricities of movement directions tended to be smaller when the direction of the other hand was less eccentric, and small eccentricities of directions tended to be larger when the direction of the other hand was more eccentric. Thus, at the longest CRI there was a (static) assimilation of directions. It was quite clearly present for movements in the medial direction, but less clearly for movements in the lateral direction. In addition, as in Experiment 1, movements to lateral targets were more eccentric than movements to medial targets when target eccentricities were 30°. However, the difference was reversed when target eccentricities were only 15°.

Asymptotic initial directions at the longest CRI of symmetric and parallel movements to lateral and medial targets as a function of same and different target eccentricities. Long bars indicate directions of movements to more eccentric targets (30°), short bars of movements to less eccentric targets (15°). (Movements in the direction of the target are marked by positive values)
The mean directions at the longest CRI were subjected to a five-way ANOVA with the within-participant factors Relation of Eccentricities (same, different), Relation of Directions (symmetric, parallel), Eccentricity (large, small), Direction (lateral, medial), and Hand (left, right). The assimilation was reflected in a significant Eccentricity×Relation of Eccentricities interaction, F(1, 11)=8.2, P<0.05. This interaction was not reliably modulated by any of the other factors. From Fig. 9 it is apparent that with different eccentricities the reduction of large eccentricities was stronger than the increase of small eccentricities. This resulted in overall smaller eccentricities when target eccentricities were different than when they were the same, F(1, 11)=5.8, P<0.05.
The difference between movements to lateral and medial targets, which was opposite at small and large eccentricities, gave rise to a significant Eccentricity×Direction interaction, F(1, 11)=53.2, P<0.01. This interaction was more pronounced for parallel than for symmetric movements, as reflected in a triple interaction with Relation between Directions as the third factor, F(1, 11)=5.3, P<0.05. Finally, parallel movements were overall more eccentric than symmetric movements, F(1, 11)=10.8, P<0.01; this difference had failed to reach statistical significance in Experiment 1.
Mean normalized initial directions
For the analysis of the time courses of direction specifications with the various combinations of target directions normalized initial directions were used as in Experiment 1. In Fig. 10 the mean normalized initial directions are shown for symmetric and parallel movements as a function of the CRI, separately for movements to lateral and to medial targets. Specification of symmetric directions was faster than specification of parallel directions, but this difference was restricted to movements to lateral targets and not observed for movements to medial targets. In other words, specification of lateral directions for both hands was faster than specification of directions when a medial direction had to be specified for at least one hand. As in Experiment 1, the difference developed at later CRIs only and not at the very start of direction specification. From Fig. 10 it is also evident that the specification of lateral directions was faster than the specification of medial directions.

Normalized initial direction of movements to lateral and medial targets as a function of the CRI and the relation between the target directions (symmetric and parallel; Experiment 2)
The normalized initial directions were subjected to a six-way ANOVA with the within-participant factors Relation of Eccentricities (same, different), Relation of Directions (symmetric, parallel), Eccentricity (large, small), Direction (lateral, medial), Hand (left, right), and CRI. The pattern of results shown in Fig. 10 gave rise to a significant interaction of Relation of Directions, Direction, and CRI, F(4, 44)=3.7, P<0.05, as well as to a significant interaction of Relation of Directions and CRI, F(4, 44)=3.7, P<0.05. The difference in the rate of specifying lateral and medial directions was reflected in a significant Direction×CRI interaction, F(4, 44)=6.0, P<0.01.
The normalized initial directions of movements with same and different target eccentricities are shown in Fig. 11 as a function of the CRI, separately for small and large eccentricities and symmetric and parallel directions. There were transient assimilation effects at intermediate CRIs: when target eccentricities were different, the normalized initial directions were too large for movements to the less eccentric targets, and they were too small for movements to the more eccentric targets. This pattern was clearly present for symmetric movements, but with parallel movements the transient assimilation was absent for movements to the more eccentric targets.

Normalized initial direction of symmetric and parallel movements to targets of high and low eccentricities as a function of the CRI, shown separately for same and different eccentricities of the targets for the two hands
In addition to the assimilation effects there were slight differences between low-eccentricity and high-eccentricity movements. These reflected the fact that the time course of specifying high-eccentricity directions was not simply an up-scaled version of the time course of specifying low-eccentricity directions. If that had been the case, the ratios of the specified high-eccentricity and low-eccentricity directions at the various CRIs should have been constant at about 2.0. However, they increased across CRIs, being (0.40, 0.43) 1.32, 1.54, 1.65, 1.74, and 1.73 (the first two ratios are in brackets because the actual directions specified deviated from the forward directions by less than 1°). The associated differences between the specified high-eccentricity and low-eccentricity directions were 0.1, 0.1, 0.4, 4.1, 9.6, 12.5, and 12.8°. Thus, at the first three CRIs (up to about 300 ms) the time courses of direction specifications were essentially the same for low-eccentricity and high-eccentricity target directions.
The assimilation shown in Fig. 11 gave rise to a three-way interaction of Relation between Eccentricities, Eccentricity, and CRI, F(4 ,44)=13.4, P<0.01, as well as to a Relation between Eccentricities×CRI interaction, F(4, 44)=3.4, P<0.05. Whereas the assimilation was clearly present in symmetric movements, it was less distinct in parallel movements, giving rise to a four-way interaction of Relation between Eccentricities, Eccentricity, CRI, and Relation between Directions, F(4, 44)=3.1, P<0.05. Similarly (though not shown in Fig. 11) the basic assimilation pattern was stronger for movements to lateral targets and weaker for movements to medial targets; the four-way interaction of Relation of Eccentricities, Eccentricity, CRI, and Direction was significant, F(4, 44)=4.3, P<0.01.
Discussion
The purpose of Experiment 2 was to replicate the major findings of Experiment 1, in particular the unexpected and statistically ambiguous ones, and to shed more light on the adaptive modulation of directional coupling during motor preparation. In particular we explored again an eventual adaptive modulation when target directions were different, and in addition we explored the role of target eccentricity for the changes of directional coupling when symmetric and parallel directions are specified.
Beyond replicating the findings of the first experiment, Experiment 2 revealed that the divergence of intermanual direction correlations for symmetric and parallel movements with increasing CRI is not restricted to target eccentricities of about 30°, but can also be seen with smaller target eccentricities of only about 15°. There are some quantitative differences, but the switch from positive to negative correlations in the course of specifying parallel directions is clearly present with both eccentricities at preparation intervals of about 300 ms. The quantitative differences between the correlations found with smaller and larger eccentricities do not necessarily reflect differences in the strength of the intermanual coupling. Instead they could also reflect differences in the hand-specific random variation of directions associated with different target eccentricities.
In Experiment 2 a nonmonotonic component of the modulation of intermanual direction correlations in the course of motor preparation was more marked than in Experiment 1. In the range of CRIs of 350–450 ms both positive (with symmetric targets) and negative (with parallel targets) intermanual correlations were particularly strong. The nonmonotonic changes of correlations can result from a simple artifact which has been noted in research on the development of the intercorrelations of tests of intelligence during childhood (Merz and Kalveram 1965). Basically, in particular for CRIs in the range of 350–450 ms, there is not only the trial-to-trial covariation, but also superposed on it is the change of the mean directions across the CRIs within each bin. The common trend of the directions of both hands with symmetric movements within each CRI bin produces additional positive covariation, whereas the divergent trend of the directions of both hands with parallel movements produces additional negative covariation. Of course, the additional covariation is absent at the longest and shortest CRIs at which the mean directions do not change. Therefore the nonmonotonic components of the modulation of the intermanual correlations of initial directions, which are present in the range of CRIs where mean directions actually change, could result from these additional sources of covariation which are not related to intermanual interactions.
To test the role of the potential artifact for the observed modulations of intermanual correlations, we computed partial correlations for each bin. The joint influence of the CRIs within each bin on the initial directions of left-hand and right-hand movements was partialed out. The mean partial correlations are shown in Table 3, averaged for the two early CRI bins, the two latest ones, and the two intermediate ones. Whereas in the early and late bins the differences between simple and partial intermanual direction correlations were only small and unsystematic, they were larger and systematic in the intermediate bins. In particular the partial correlations were less extreme than the simple ones. In spite of this reduction the nonmonotonic modulation was still clearly present even in the partial correlations.
Whereas there is a strong divergence of intermanual direction correlations in the course of specifying symmetric and parallel movement directions, there is essentially no divergence when same and different eccentricities are specified. The differences between symmetric movements with same and different eccentricities as well as between parallel movements with same and different eccentricities are only small and statistically not significant. This confirms the result of Experiment 1 that the specification of different directions (or different eccentricities) is not associated with a de-coupling as indicated by a decline of intermanual direction correlations, and it contrasts sharply with the findings for movements with same and different amplitudes (Heuer et al. 1998, 2001; Sherwood, 1991) and isometric contractions with same and different peak forces (Rinkenauer et al. 2001; Steglich et al. 1999).
Intermanual direction correlations are only one kind of data which allow inferences about intermanual directional coupling. The other kind of data are assimilation effects, and inferences from both kinds of data should be consistent. This, in fact, is what they are. Experiment 2 confirms the finding of static assimilation effects at long CRIs when the eccentricities of movement directions are different. Extending the results of Experiment 1, Experiment 2 reveals that the static assimilation effects have a symmetric or parallel pattern depending on the target directions. For symmetric movements with different eccentricities, assimilation effects produce a bias toward symmetric movements with same eccentricities. For parallel movements with different eccentricities, assimilation effects produce a bias toward parallel movements with same eccentricities. The different patterns of the static assimilation effects match the findings on intermanual direction correlations at the long CRIs which also indicate a symmetric and parallel coupling for symmetric and parallel directions, respectively. In addition the static assimilation effects are consistent with a lack of de-coupling when directions with different eccentricities are specified concurrently. If de-coupling had been present, static assimilation effects should have been absent.
The phasic assimilation effects observed in movements to targets with different eccentricities are quite similar to the static effects. They remain after normalization of the amplitudes. Normalization removes static effects, also those static effects which evolve gradually in the course of direction specifications. Phasic effects of the same kind as static effects thus indicate that the assimilation is proportionally stronger at intermediate CRIs than at long ones. The symmetric and parallel patterns of the observed phasic assimilation effects match the finding of particularly strong positive and negative intermanual direction correlations at intermediate CRIs. At the shortest three CRIs (up to about 300 ms) assimilation effects are lacking for the simple reason that at these early times in the process of direction specification the actually specified eccentricities do not yet differ depending on the eccentricities of the target directions.
Consistent with the somewhat inconclusive observations of Steglich (2002) and of Experiment 1, in Experiment 2 there was clear evidence that the directions of parallel movements are specified more slowly than the directions of symmetric movements. This is true, however, only for lateral directions, but not for medial directions. Thus, the combination lateral–lateral is specified at a faster rate than the other three combinations of directions, lateral–medial, medial–lateral, and medial–medial. It seems unlikely that these differences result from the intermanual coupling of directions. For those CRIs, at which the different rates are observed, intermanual coupling is already strongly influenced by the combination of target directions so that it is symmetric or parallel, respectively.
General Discussion
The purpose of the present experiments was to determine the modulation of intermanual interactions during preparation of bimanual movements with different combinations of directions. There is considerable evidence that direction, in addition to amplitude, is an important parameter in the control of movements aimed at a target (Gordon et al. 1994; Krakauer et al. 2004; Rossetti et al. 1995; Vindras and Viviani 2002). Whereas intermanual interactions with respect to amplitudes have received a good deal of research effort, intermanual interactions with respect to movement directions have been largely neglected until recently. Recent studies have used a reaction-time paradigm (e.g., Diedrichsen et al. 2001; Weigelt 2004) or a periodic-movement paradigm (Swinnen et al. 2001, 2002, 2003; Wenderoth et al. 2004). Their results strongly suggest the need to study intermanual interactions during the specification of movement directions in more detail.
The findings of the present experiments reveal that during the specification of the directions of bimanual movements intermanual interactions are modulated depending on the task requirements. A task-dependent modulation has also been found during the specification of the amplitudes of bimanual movements (Heuer et al. 1998, 2001), and the peak forces of bimanual isometric contractions (Steglich et al. 1999). However, the pattern of modulation during direction specification is quite different and more complex. The peculiarities are perhaps related to the fact that only for the direction continuum there is a symmetry relation, but not for the amplitude and peak-force continua.
Task-dependent modulations of intermanual interactions are less than perfect. Even though task constraints seem to shape intermanual coupling to a considerable degree, there are structural constraints on which task-dependent modulations are superposed (cf. Heuer 1996). We shall discuss both the modulation, that is, the effects of task constraints, and the limits of modulation, that is, the effects of structural constraints, in turn.
Task constraints and the adaptive modulation of directional coupling
Both the intermanual direction correlations and the assimilation effects indicate that intermanual interactions are modulated during the specification of bimanual movement directions. Specifically, for symmetric movements the symmetric coupling is maintained, which can also be observed for forward movements at short preparation intervals. In contrast, for parallel movements an initial symmetric coupling is turned into a parallel coupling after a preparation period of about 300 ms. The switch from symmetric to parallel coupling largely precedes the specification of target directions. Thus, the changes of the intermanual correlations are observed at CRIs at which movement directions hardly deviate or do not at all deviate from the (default) forward directions. When symmetric or parallel movement directions are finally specified, there are symmetric or parallel assimilation effects, respectively, when target eccentricities are different.
Even though intermanual interactions are modulated in an adaptive manner for the specification of symmetric and parallel directions, there is no adaptive modulation for the specification of different directions. This contrasts sharply with the adaptive de-coupling found during the specification of different amplitudes of bimanual movements (Heuer et al. 1998, 2001) or different peak forces of bimanual isometric contractions (Steglich et al. 1999). Therefore, the assimilation effects found with different eccentricities are static and not only transient. They persist at long preparation intervals and do not vanish (or become asymmetric) as is the case for assimilations of amplitudes. Nevertheless, the basically static effects can be transiently enhanced. This can happen during the time periods in which symmetric and parallel directions are actually specified. During these time periods symmetric and parallel coupling, respectively, can be particularly tight, as indicated by phasically enhanced intermanual correlations.
The observation that specifications of parallel and symmetric directions are associated with different time courses of directional coupling, whereas specifications of directions with same and different eccentricities are not, is consistent with the notion of a hierarchical coding of movement directions (cf. Huttenlocher et al. 1991; Messier and Kalaska 1999). At one level the coding is categorical, e.g., in terms of quadrants of a field, in terms of left–right or in terms of lateral–medial. In the case of bimanual movements these categorical codes form an even higher level with the categories “symmetric” and “parallel”. At the lowest level there is a metrical coding; here the actual eccentricities are represented.
Evidence for hierarchical coding of spatial locations (which includes the coding of directions) is primarily from studies of the reproduction of the locations of visually presented dots (Huttenlocher et al. 1991) or of the accuracy of pointing to memorized target locations (Messier and Kalaska, 1999). In both types of studies levels of coding are revealed by error distributions. Reproductions or movement endpoints are attracted by the prototypes of the categories, in particular when the interval between the presentation of the target and reproduction (or pointing) is not too short. In the present experiments only the categorical coding of target directions is related to the adaptive modulation of intermanual coupling. In contrast, metrical coding is not used for the modulation, but only for the specification of movement directions. A certain analogy to the use of only categorical representations for a certain purpose has been reported for the Simon effect. For this effect it matters whether (irrelevant) stimulus locations are left or right and responses are left or right, but the degree of “leftness” or “rightness” does not (e.g., Logan 2003).
The different patterns of the adaptive modulation of amplitude coupling and directional coupling in the course of specifying various combinations of amplitudes or directions, respectively, are consistent with the observations that the neural coding of amplitudes and directions is different (e.g., Fu et al. 1993, 1995) and that different brain structures are involved in their specifications (e.g., Desmurget et al. 2003; Krakauer et al. 2004) and their intermanual interactions (Wenderoth et al. 2005). However, the differences in neural coding and neural substrates do not imply particular differences with respect to the adaptive modulation. For them there should be more specific reasons. One might suspect, for example, that certain advantages are inherent to the de-coupling of amplitudes when they are different, but the maintained coupling rather than de-coupling of different directions. In general, coupling should be associated with constant errors, namely assimilation effects, but de-coupling with variable errors (cf. Heuer et al. 2001). Perhaps the modulations of coupling during motor preparation are related to the relative costs of coupled and de-coupled specifications in terms of systematic and random errors, respectively.
Structural constraints and the limits of adaptive modulations
One of the most pervasive structural constraints on bimanual coordination is the symmetry constraint. In the present experiments the adaptive modulation of intermanual interactions was superposed on it. Its continued operation is revealed by at least two observations. First, intermanual direction correlations indicate a symmetric coupling for forward movements. These movements are both symmetric and parallel, and thus do not constrain the appropriate kind of coupling. Second, at long preparation intervals symmetric coupling is stronger than parallel coupling, again indicated by the intermanual direction correlations.
Intermanual interactions, as can be observed in overt behavior, result from neural interactions at various levels of the central nervous system. As candidate structures involved in bimanual coordination, Donchin et al. 2004 list several cortical areas including not only the primary motor cortex and the supplementary motor area, but also the parietal cortex and, of course, the corpus callosum. In addition intermanual interactions can originate in the cerebellum, the basal ganglia, and the spinal cord. It is likely that structural constraints on bimanual coordination and the adaptive shaping of intermanual interactions by the task constraints have different neural substrates. Perhaps structural constraints are adapted to fundamental movement tasks like standing and locomotion and originate from lower-level structures, whereas modulations are primarily of cortical origin. Possibly higher-level structures do not only add to the intermanual interactions established by lower-level structures, but also modulate the strength of lower-level interactions to some extent.
In spite of the evidence for a basic symmetry constraint seen in intermanual correlations, there are no corresponding assimilation effects. The most likely reason is that with parallel target directions symmetric coupling is already switched to parallel coupling when the specified directions start to deviate from the forward default directions. Nevertheless, at longer preparation intervals symmetric directions are specified at a faster rate than parallel directions. More precisely, only lateral symmetric directions are specified at a faster rate than combinations of movement directions which include at least one medial direction. In addition, medial directions are specified at a slower rate than lateral directions. Thus, the findings suggest a kind of constraint similar to one observed in rapid aiming movements (e.g., Kelso et al. 1979) in that with concurrent processes the normally faster one is slowed down, but the normally slower one is not speeded up.
It is tempting to speculate about the reasons of the rate differences in specifying lateral and medial directions. There is an inertial anisotropy (cf. Gordon et al. 1994) in that the mass moved in lateral directions is smaller than the mass moved in medial directions. Perhaps more important, however, are differences in the sets of muscles involved and their required coordination. Specification of movement directions ultimately implies the specification of certain patterns of muscle activations (e.g., Thoroughman and Shadmehr 1999). Thus, there should be a transformation from the spatial coding of directions to coding in terms of patterns of muscle activity. Kakei et al. (2003) found corresponding types of neurons. In ventral premotor cortex extrinsic-space neurons were the majority among directionally tuned ones, whereas in primary motor cortex muscle-related neurons were of about the same frequency. The transformation of direction in extrinsic space into the appropriate pattern of muscle activity could be slower for movements to medial targets than for movements to lateral targets because a more complex pattern of muscle activity has to be specified.
Adaptive parametric coupling as a component of motor preparation
Intermanual interactions have been ascribed to the cross-talk between different kinds of signals involved in the control of left-hand and right-hand movements. The findings reported by Swinnen et al. (2001, 2003) suggest that for interactions with respect to movement directions neither outflow signals nor (visual) feedback signals are critical. Instead parametric coupling might be critical, that is, cross-talk between signals involved in the specification of movement characteristics (or parameters) (Heuer 1993; Spijkers and Heuer 1995). The present results are consistent with this claim. In addition, and this is perhaps more important, they strengthen the view that motor preparation not only embraces the specification of movement characteristics, but also the appropriate adjustments of interlimb interactions. In the present experiments the adjustments of intermanual interactions largely preceded the actual specification of symmetric or parallel directions, similar to how postural adjustments precede voluntary movements (Massion 1992, for an overview).
From a somewhat broader perspective, adaptive modulations of intermanual interactions can be conceived as configurations of task sets. Thus far the notion of a task set (cf. Rogers and Monsell 1995) has generally been used to refer to the functional correlates of the intention to process stimuli in a certain way. However, with a broader notion of a task set as a task-control structure, it is evident that there is not only a “cognitive structure” related to processing of input information but also a “motor structure” related to the control of the movements required by the task (cf. Kleinsorge and Heuer 1999). The setting of task-appropriate intermanual interactions is a basic ingredient of task sets for tasks which involve multi-effector movements. The established couplings then form the background on which the particular movement parameters like directions and amplitudes are specified.
Notes
The different effects of measurement noise on the intermanual correlations of terminal and initial directions result from the different ranges of coordinates and can easily be illustrated by way of simulation. We generated two sets of correlated normally distributed directions (μ=30, σ2=5 ρ=0.8). For each direction we determined the x- and y-coordinates for vector lengths of 20 and 150 mm. These coordinates were rounded to units of 0.5 mm, corresponding to the coordinates used in the determination of initial and terminal directions in the experiment. For each length the directions were re-computed from the rounded coordinates. For a typical sample of 1000 cases the means were 29.98° and 30.02° for the left and right hand, respectively, the standard deviations were 2.22 and 2.25°, and the intermanual correlation was 0.8048. Means (and standard deviations) for the re-computed terminal directions were 29.98° (2.22°) and 30.02° (2.25°) with a correlation of 0.8050; means (and standard deviations) for the re-computed initial directions were 29.96° (2.25°) and 30.03° (2.30°) with a correlation of 0.7751. The differences in standard deviations and correlations are consistent across samples, but not the difference in the means.
References
Cattaert D, Semjen A, Summers JJ (1999) Simulating a neural cross-talk model for between-hand interference during bimanual circle drawing. Biol Cybern 81:343–358
Desmurget M, Grafton S, Vindras P, Grea H, Turner RS (2003) Basal ganglia network mediates the control of movement amplitude. Exp Brain Res 153:197–209
Diedrichsen J, Hazeltine E, Kennerley S, Ivry RB (2001) Moving to directly cued locations abolishes spatial interference during bimanual actions. Psychol Sci 12:493–498
Donchin O, Cardoso de Oliveira S (2004) Electrophysiological approaches to bimanual coordination in primates. In: Swinnen SP, Duysens J (eds) Neuro-behavioral determinants of interlimb coordination. Kluwer, Norwell, Mass, pp 131–153
Fu Q-G, Suarez JI, Ebner TJ (1993) Neuronal specification of direction and distance during reaching movements in the superior precentral motor area and primary motor cortex of monkeys. J Neurophysiol 70:2097–2116
Fu Q-G, Flament D, Coltz JD, Ebner TJ (1995) Temporal encoding of movement kinematics in the discharge of primate motor and premotor neurons. J Neurophysiol 73:836–854
Ghez C, Favilla M, Ghilardi MF, Gordon J, Bermejo R, Pullman S (1997) Discrete and continuous planning of hand movements and isometric force trajectories. Exp Brain Res 115:217–233
Gordon J, Ghilardi MF, Cooper SE, Ghez C (1994) Accuracy of planar reaching movements. II. Systematic extent errors resulting from inertial anisotropy. Exp Brain Res 99:112–130
Gordon J, Ghilardi MF, Ghez C (1994) Accuracy of planar reaching movements. I. Independence of direction and extent variability. Exp Brain Res 99:97–111
Hening W, Favilla M, Ghez C (1988) Trajectory control in targeted force impulses. V. Gradual specification of response amplitude. Exp Brain Res 71:116–128
Heuer H (1990) Rapid responses with the left or right hand: response–response compatibility effects due to intermanual interactions. In: Proctor RW, Reeve TG (eds) Stimulus-response compatibility: An integrated perspective. North-Holland, Amsterdam, pp 311–342
Heuer H (1993) Structural constraints on bimanual movements. Psychol Res 55:83–98
Heuer H (1996) Coordination. In: Heuer H, Keele SW (eds) Handbook of Perception and Action. Vol. 2: Motor skills. Academic Press, London, pp 121–180
Heuer H, Klein W (2005) Intermanual interactions in discrete and periodic bimanual movements with same and different amplitudes. Exp Brain Res
Heuer H, Spijkers W, Kleinsorge T, van der Loo H, Steglich C (1998) The time course of cross-talk during the simultaneous specification of bimanual movement amplitudes. Exp Brain Res 118:381–392
Heuer H, Kleinsorge T, Spijkers W, Steglich C (2001) Static and phasic cross-talk effects in discrete bimanual reversal movements. J Mot Behav 33:67–85
Huttenlocher J, Hedges LV, Duncan S (1991) Categories and particulars: prototype effects in estimating spatial location. Psychol Rev 98:352–376
Kakei S, Hoffman DS, Strick PL (2003) Sensorimotor transformations in cortical motor areas. Neurosci Res 46:1–10
Kelso JAS, Southard DL, Goodman D (1979) On the coordination of two-handed movements. J Exp Psychol Hum Percept Perform 5:229–238
Kleinsorge T, Heuer H (1999) Hierarchical switching in a multi-dimensional task space. Psychol Res 62:300–312
Krakauer JW, Ghilardi MF, Mentis M, Barnes A, Veytsman M, Eidelberg D, Ghez C (2004) Differential cortical and subcortical activations in learning rotations and gains for reaching: a PET study. J Neurophysiol 91:924–933
Levin O, Ouamer M, Steyvers M, Swinnen SP (2001) Directional tuning effects during cyclical two-joint arm movements in the horizontal plane. Exp Brain R 141:471–484
Logan GD (2003) Simon-type effects: Chronometric evidence for keypress schemata in typewriting. J Exp Psychol Hum Percept Perform 29:741–757
Marteniuk RG, MacKenzie CL, Baba DM (1984) Bimanual movement control: Information processing and interaction effects. Q J Exp Psychol 36A:335–365
Massion J (1992) Movement, posture, and equilibrium: Interaction and coordination. Prog Neurobiol 38:35–56
Mechsner F, Kerzel D, Knoblich G, Prinz W (2001) Perceptual basis of coordination. Nature 414:69–73
Merz F, Kalveram K-Th (1965) Kritik der Differenzierungshypothese der Intelligenz. Arch Gesamte Psychol 117:287–295
Messier J, Kalaska JF (1999) Comparison of variability of initial kinematics and endpoints of reaching movements. Exp Brain Res 125:139–152
Preilowski B (1975) Bilateral motor interaction: Perceptual-motor performance of partial and complete “split-brain” patients. In: Zülch KJ, Creutzfeldt O, Galbraith GG (eds) Cerebral localization. Springer, Berlin Heidelberg New York pp 115–132
Rinkenauer G, Ulrich R, Wing AM (2001) Brief bimanual force pulses: correlations between the hand in force and time. J Exp Psychol Hum Percept Perform 27:1485–1497
Rogers RD, Monsell S (1995) The cost of a predictable switch between simple cognitive tasks. J Exp Psychol Gen 124:207–231
Rossetti Y, Desmurget M, Prablanc C (1995) Vector coding of movement: vision, proprioception, or both? J Neurophysiol 74:457–463
Sherwood DE (1990) Practice and assimilation effects in a multilimb aiming task. J Mot Behav 22:267–291
Sherwood DE (1991) Distance and location assimilation in rapid bimanual movement. Res Q Exerc Sport 62:302–308
Sherwood DE (1994a) Interlimb amplitude differences, spatial assimilations, and the temporal structure of rapid bimanual movements. Hum Mov Sci 13:841–860
Sherwood DE (1994b) Hand preference, practice order, and spatial assimilations in rapid bimanual movement. J Mot Behav 26:123–134
Sherwood DE, Nishimura KM (1992) EMG amplitude and spatial assimilation effects in rapid bimanual movement. Res Q Exerc Sport 63:284–291
Spijkers W, Heuer H (2004) Behavioral principles of interlimb coordination. In: Swinnen SP, Duysens J (eds) Neuro-behavioral determinants of interlimb coordination. Kluwer, Norwell, Mass, pp 223–258
Spijkers W, Heuer H (1995) Structural constraints on the performance of symmetrical bimanual movements with different amplitudes. Q J Exp Psychol 48A:716–740
Steglich C (2002) Experimentelle Untersuchungen zur bimanuellen Koordination als transiente Kopplung bei der Richtungsspezifikation. Unpublished Dissertation, RWTH Aachen
Steglich C, Heuer H, Spijkers W, Kleinsorge T (1999) Bimanual coupling during the specification of isometric forces. Exp Brain Res 129:302–316
Swinnen SP, Dounskaia N, Levin O, Duysens J (2001) Constraints during bimanual coordination: the role of direction in relation to amplitude and force requirements. Behav Brain Res 123:201–218
Swinnen SP, Dounskaia N, Duysens J (2002) Patterns of bimanual interference reveal movement encoding within a radial egocentric reference frame. J Cogn Neurosci 14:463–471
Swinnen SP, Puttemans V, Vangheluwe S, Wenderoth N, Levin O, Dounskaia N (2003) Directional interference during bimanual coordination: is interlimb coupling mediated by afferent or efferent processes. Behav Brain Res 139:177–195
Thoroughman KA, Shadmehr R (1999) Electromyographic correlates of learning internal models of reaching movements. J Neurosci 19:8573–8588
Vindras P, Viviani P (2002) Altering the visuomotor gain. Evidence that motor plans deal with vector quantities. Exp Brain Res 147:280–295
Weigelt M (2004) Target-related coupling in bimanual coordination. Unpublished Dissertation. LMU Munich
Wenderoth N, Debaere F, Sunaert S, van Hecke P, Swinnen SP (2004) Parieto-premotor areas mediate directional interference during bimanual movements. Cereb Cortex 14:1153–1163
Wenderoth N, Debaere F, Sunaert S, Swinnen SP (2005) Spatial interference during bimanual coordination: differential brain networks associated with control of movement amplitude and direction. Hum Brain Mapp 26:286–300
Acknowledgements
The research reported in this paper was supported by grant HE 1187/14-1 of the Deutsche Forschungsgemeinschaft. We thank Barbara Herbst, Holger Küper, Kevin Schepers, Henning Stracke, and Petra Wallmeyer for their support in running the experiments, and Thomas Kleinsorge, Stephan Swinnen, and two anonymous reviewers for helpful comments on earlier versions of this paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Heuer, H., Klein, W. The modulation of intermanual interactions during the specification of the directions of bimanual movements. Exp Brain Res 169, 162–181 (2006). https://doi.org/10.1007/s00221-005-0135-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-005-0135-6