
Abstract
Symmetric, target-directed, bimanual movements take less time to prepare than asymmetric movements (Diedrichsen et al. in Cerebral Cortex 16(12):1729–1738, 2006; Heuer and Klein in Psychol Res 70(4):229–244, 2006b). The preparation savings for symmetric movements may be related to the specification of symmetric amplitudes, target locations, or both. The goals of this study were to determine which symmetric movement parameters facilitate the preparation of bimanual movements and to compare the size of the facilitation for different parameters. Thirty participants performed bimanual reaching movements that varied in terms of the symmetry/asymmetry of starting locations, movement amplitudes, and target locations. Reaction time savings were examined by comparing movements that had one symmetric parameter (and two asymmetric parameters) to movements with all asymmetric parameters. We observed significant savings (~10 ms) for movements with symmetric amplitudes and movements with symmetric target locations. Reaction time costs were examined by comparing movements that had two asymmetric parameters (and one symmetric parameter) to movements with all symmetric parameters. We observed significant reaction time costs (~13 ms) for all movements with asymmetric amplitudes. These results suggest that movement preparation is facilitated when amplitudes or target locations are symmetric and that movement preparation suffers interference when amplitudes are asymmetric. The relative importance of the three parameters to movement preparation, from most to least important, is movement amplitudes, target locations, and then starting locations. Interference with asymmetric amplitudes or target locations may be caused by cross-talk between concurrent processes of parameter specification during response programming.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coordinated movements with the two arms are important for a plethora of activities of daily living. As common as these movements are, it is difficult to move each arm with different spatio-temporal features; for example, think of how much practice is needed to coordinate your arms while cascade juggling. One approach to studying bimanual coordination has been to test bimanual reaching movements where the arms have the same (symmetric) or different (asymmetric) movement amplitudes [reviewed by Wenderoth and Weigelt, (2009)]. Kelso, Southard, and Goodman, (1979) compared the movement times of bimanual symmetric movements with short or long movement amplitudes to bimanual asymmetric movements where one arm had a short movement amplitude and the other has a long amplitude. The finding of interest was that during bimanual asymmetric movements the movement time of a short amplitude movement was lengthened to be comparable to the movement time of a long movement. Kelso et al. argued that this temporal assimilation of movement times suggested that bimanual movements are not simply the sum of two unimanual movements; instead, the two arms of a bimanual movement are prepared as a functional unit.
Subsequent studies probed the complexity of movement preparation for bimanual symmetric and asymmetric reaching movements. It was reasonably assumed that more complex movement preparation requires more processing time. Therefore, increased reaction time (RT) was used as an indicator of increased complexity. Spijkers, Heuer, Kleinsorge & van der Loo, (1997) compared the RTs of bimanual symmetric and asymmetric reversal movements. The movements were cued symbolically; for example, in the first experiment, the German words for short and long were used to indicate the target movement amplitudes for the left and right arms. They found longer RTs for bimanual asymmetric movements compared to symmetric movements. Spijkers et al. argued that intermanual cross-talk during the response programming of asymmetric amplitudes caused more complex movement preparation for bimanual asymmetric movements.
A potential confound was investigated by Diedrichsen, Hazeltine, Kennerley & Ivry (2001). They reasoned that the bimanual asymmetric costs could be caused by symbolic cues and not bimanual interference during response programming. Asymmetric movements were cued with two different symbolic cues and this may place greater processing demands on response selection compared to two identical cues for symmetric movements. Diedrichsen et al. compared the bimanual asymmetric costs for symbolically and directly cued movements, with the assumption that direct cues place minimal processing demands on response selection. They found large bimanual asymmetric costs with symbolic cues (55 ms, Experiment 1) and non-significant asymmetric costs with direct cues (6 ms, Experiment 1). These results were used to argue that there are large costs to translate two different symbolic cues and that there are no costs to prepare bimanual asymmetric movements.
More recently, several studies have shown that there are small processing costs (typically 10–20 ms) to prepare bimanual asymmetric movement, even when movements are directly cued (Blinch et al., 2014; Diedrichsen, Grafton, Albert, Hazeltine & Ivry, 2006; Franz & McCormick, 2010; Heuer & Klein, 2006b; Stelmach, Amrhein & Goggin, 1988; Weigelt & Cardoso de Oliveira, 2003). Collectively, the findings support the position that movement preparation is more complex for bimanual asymmetric movements than symmetric movements. A common feature of these previous studies is that the bimanual symmetric movements had symmetric starting locations, movement amplitudes, and target locations. The bimanual asymmetric movements had the hands starting at symmetric locations but travelling asymmetric amplitudes to asymmetric target locations. This difference in parameters between symmetric and asymmetric movements raises the following question: Do asymmetric movement amplitudes, asymmetric target locations, or both cause the bimanual asymmetric costs? In this study, we examined three kinds of spatial symmetry in bimanual reaching: starting location symmetry, movement amplitude symmetry, and target location symmetry to assess whether these parameters influence movement preparation. Two studies (Heuer & Klein, 2006a; Weigelt, 2007) have tried to isolate the independent effects of these parameters.
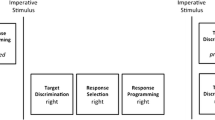
Heuer and Klein, (2006a) investigated the contribution of asymmetric movement amplitudes and target locations to the bimanual asymmetric costs. They tested all possible combinations of bimanual reversal movements with symmetric or asymmetric starting locations, amplitudes, and target locations (Fig. 1). The order of the various movement combinations was randomized for each block of trials. Although starting locations were manipulated, their contribution to the bimanual asymmetric costs was not analysed. Heuer and Klein concluded that asymmetric amplitudes and asymmetric target locations contributed additively to the asymmetric costs.

The 16 movements tested by Heuer and Klein, (2006a). Symmetric and asymmetric starting locations, movement amplitudes, and target locations were tested, but only the effects of amplitudes and target locations on RTs were evaluated. The label above each column indicates the symmetric parameters (lowercase letters) and the asymmetric parameters (uppercase letters). The number at the bottom of each cell indicates the number of movement choices for those starting locations. Adapted from Heuer and Klein, (2006a)
The interaction between amplitudes and target locations could not be fully dissociated because of a confound in the study: the number of movement choices depended on the starting locations. This ranged from 4-choice RT to simple RT. (The number at the bottom of each cell in Fig. 1 indicates the number of movement choices for those starting locations.) This is a problem because RTs can increase with the number of movement choices (Hick, 1952; Hyman, 1953; Favilla, 1996; Wright, Marino, Belovsky & Chubb, 2007).
Weigelt, (2007) re-examined the contributions from movement amplitudes and target locations in bimanual reaches, and he also considered the contribution from starting locations. Another advantage to Weigelt’s design was that it included four movement choices in all of the starting locations. As with the previous study, the order of the various movement combinations was randomized for each block of trials. A symbolic cue (the letter A or B) indicated the movement for each arm. The letter A cued the short distance target and B cued the long target. The sixteen bimanual movements were divided into five types of movements based on the symmetry/asymmetry of the movement parameters. The five movement types were:
-
1.
All parameters symmetric (sat)
-
2.
Symmetric movement amplitudes (asymmetric starting and target locations; SaT)
-
3.
Symmetric starting locations (asymmetric amplitudes and target locations; sAT)
-
4.
Symmetric target locations (asymmetric starting locations and amplitudes; SAt)
-
5.
All parameters asymmetric (SAT).
The bimanual asymmetric costs can also be thought of as RT savings for symmetric movements. Potential RT savings were investigated by comparing the RTs of movements with one symmetric parameter (and two asymmetric parameters; SaT, sAT, SAt) to movements with all asymmetric parameters (SAT). These comparisons yielded three results: (1) RT savings for movements with symmetric target locations [SAt vs. SAT], (2) no RT savings for movements with symmetric amplitudes [SaT vs. SAT], and (3) RT costs for movements with symmetric starting locations [sAT vs. SAT].
The limitation with Weigelt’s, (2007) use of symbolic cues is that RTs are longer when the cues are different for each arm (AB or BA) compared to when they are the same (AA or BB; see Diedrichsen et al., 2001). The types of movements in Weigelt’s, (2007) experiment that had different symbolic cues (SAT, 579 ms; sAT, 600 ms; SaT, 583 ms) had substantially longer RTs than movements with the same cues (SAt, 554 ms; sat, 551 ms). The first comparison was confounded by comparing movements with different symbolic cues to movements with the same cues (SAt vs. SAT). Therefore, it was not surprising that there were RT savings for movements with symmetric target locations (identical symbolic cues) compared to movements with all asymmetric parameters (different symbolic cues). The same symbolic cues, symmetric target locations, or both could have caused these savings.
The goals of this study were to determine which symmetric movement parameters facilitate the preparation of bimanual movements and to compare the size of the facilitation for different parameters. The contributions from starting locations, movement amplitudes, and target locations were considered. We eliminated confounds from previous experiments by equating the number of movement choices in each starting location and by directly illuminating the targets. Movements with one symmetric parameter and two asymmetric parameters were compared to movements with all asymmetric parameters for potential RT savings and to movements with all symmetric parameters for potential RT costs. The effects of the three symmetric and asymmetric parameters on the movement times of bimanual movements were also investigated.
Method
Participants
Thirty volunteer participants were tested from the university community (mean age of 23.6 years, 13 female). All participants reported being right-handed and had normal or corrected-to-normal vision. The research ethics board at the University of British Columbia approved the study, and participants gave informed written consent before participation.
Apparatus
Participants were seated at a table and their midsagittal plane was aligned with the middle of a button box placed on the table. On the surface of the button box was a four-row by two-column array of square pushbuttons (1.4 by 1.4 cm). The distance from the middle of one row of buttons to the next row was 10 cm and the distance between the two columns was 15 cm. The buttons could be either starting locations or target locations of bimanual reaches. Illuminating an LED inside the button served as a direct cue for the starting and target locations.
Design
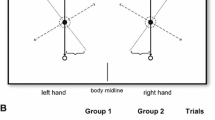
The experiment consisted of testing six blocks of bimanual reaches in 2-choice RT conditions (Fig. 2). The starting locations in each block were held constant, and the left and right arms could be cued to reach for either a short distance (10 cm) or a long distance (20 cm) target. The blocks were also designed to isolate the various parameters of bimanual reaches (starting locations, movement amplitudes, and target locations). There was one block where all parameters of the movements were symmetric (symmetric starting locations, amplitudes, and target locations; Fig. 2, sat). By symmetric starting and target locations, we are referring to buttons in the same row; symmetric amplitudes imply that both arms reached similar distances (either 10 cm or 20 cm). There were two blocks with symmetric amplitudes and asymmetric starting and target locations (Fig. 2, SaT), and one block with symmetric starting locations and asymmetric amplitudes and targets (Fig. 2, sAT). The last two blocks included one movement with symmetric targets and asymmetric starting locations and amplitudes and one movement with all asymmetric parameters (Fig. 2, SAt and SAT). The order of these blocks was counterbalanced with a balanced Latin square design.

The twelve movements tested in this study and their symmetric and asymmetric parameters. The six columns represent the six blocks with two movement choices. The label above each movement indicates the symmetric parameters (lowercase letters) and the asymmetric parameters (uppercase letters)
A block consisted of practice trials followed by test trials. The first block had 32 practice trials (16 of each movement choice) and the remaining blocks had 16 practice trials (8 of each movement choice); there were 40 test trials in every block (20 of each movement choice). The order of the movement choices was randomized in each block, and the same randomisation was used for each participant.
Procedure
At the start of each block the experimenter covered the unused buttons with black construction paper that matched the surface of the button box. Participants were then shown (with gestures) the starting locations and the target locations for the two potential movement choices in that block. Using gestures instead of words prevented the potential bias of using conceptually unified or separate language to describe the bimanual movements (Franz & McCormick, 2010). Trials began with the illumination of the starting locations for the left and right arms. This was the cue for the participants to press and hold these buttons down. Each participant decided at the beginning of the experiment whether they would press the buttons with either their index fingers or their index and middle fingers, based on whichever was more comfortable. This decision was enforced for the entire experiment. When the buttons were depressed, the lights inside the starting locations were turned off and a 1–2 s variable foreperiod began. The imperative stimuli consisted of illuminating the target locations, and participants were instructed to “react and hit the targets as fast as possible.” The target lights were turned off when the target buttons were depressed, and the participants were told to keep the buttons depressed. The starting locations for the next trial were illuminated 2 s after the targets were pressed, at which point the participants could release the target buttons and begin the next trial when ready.
Each trial was labelled as “good” or “bad,” with bad trials being recycled to the end of the block and excluded from subsequent analyses. Types of bad trials were target errors (missing the target button or hitting an incorrect button), anticipation (RT < 100 ms), inattention (RT > 750 or movement time >500 ms) and asynchronous movement initiation (>60 ms RT difference between the arms). If it was a bad trial, then the experimenter explained the reason to the participants, but participants were not told that these trials were recycled. In total, 0.6 % of all the trials were recycled.
A “speed score” was given to participants after every six good trials. This score gave participants feedback on their total response time (RT + movement time) and was intended to motivate them to react and hit the targets as fast as possible. It was calculated by comparing the mean total response time of the last six good trials to the previous six good trials. The percent change was calculated and multiplied by ten; for example, if the total response time decreased from 300 to 270 ms (a 10 % change), then the speed score would be −100 [−100 = 1,000 × (270/300 − 1)]. A multiple of ten was used to make the small changes in total response time (typically less than 5 %) more salient to the participants.
Data acquisition and analysis
The state of each button (open or closed) was sampled with a 12-bit digital input–output card. These states were used to calculate the RT (from imperative stimulus to release of the starting button) and movement time (from release of the starting button to depression of the target button) of the left and right arms. Mean RTs for each participant were calculated for each arm and movement choice in the six blocks. Means were then calculated for the five movement types (sat, SaT, sAT, SAt, SAT), collapsing across hand and movement choice, to isolate the parameters of bimanual reaches. These RTs were analysed with a one-way repeated measures ANOVA with five levels. If there was a significant difference in RTs for the five movement types, then three sets of post hoc comparisons were performed.
First, potential RT savings were examined by comparing the RTs of movements with one symmetric parameter (SaT, sAT, SAt) to movements with all asymmetric parameters (SAT). Second, the RTs of the movements with one symmetric parameter (SaT, sAT, SAt) were contrasted with three comparisons. This allowed us to make inferences about the size of the relative savings for each symmetric parameter.Footnote 1 Third, potential RT costs were examined by comparing the RTs of movements with two asymmetric parameters (SaT, sAT, SAt) to movements will all symmetric parameters (sat). An advantage for evaluating RT savings compared to costs is that a savings can be isolated to the change in one movement parameter; for example, the only difference between the SaT and the SAT movement types is that amplitudes are either asymmetric or symmetric. RT costs are somewhat more complicated because they involve a change in two movement parameters. These three sets of post hoc tests each involved three comparisons, for a total of nine, two-tailed comparisons. The familywise error rate was controlled with the Šidák correction based on nine comparisons.
Mean movement times for each participant were calculated for each arm and movement choice in the six blocks. Means were then calculated for the five movement types (sat, SaT, sAT, SAt, SAT) and the two movement amplitudes (long, short), collapsing across hand and movement choice. These movement times were analysed with a five Movement Type (sat, SaT, sAT, SAt, SAT) by two Movement Amplitude (long, short) repeated measures ANOVA. The significant interaction was examined with simple main effects on Movement Type. The familywise error rate was controlled with the Šidák correction.
When local sphericity was violated (as indicated by Mauchly’s test, p < 0.10), the Huynh–Feldt correction was used when the ε was greater than or equal to 0.75 and the Greenhouse-Geisser correction was used otherwise (Huynh & Feldt, 1976). The uncorrected degrees of freedom and the ε values were reported (Huynh–Feldt εHF, Greenhouse-Geisser εGG). Reported values are means ± the standard errors.
Results
Mean RTs for the five movement types are shown in ascending order in Fig. 3. Movements with all symmetric parameters had the shortest mean RTs (sat, 256 ± 3.9 ms) and movements with all asymmetric parameters had the longest RTs (SAT, 275 ± 4.0 ms). The three movement types with one symmetric parameter (and two asymmetric parameters) fell in between these conditions; in ascending order of mean RTs, they were symmetric movement amplitude (SaT, 262 ± 4.1 ms), symmetric starting locations (sAT, 268 ± 3.9 ms), and symmetric target locations (SAt, 269 ± 4.0 ms). Statistical analysis of the RTs for the five types of movements revealed a significant main effect, F(4, 116) = 14, p < 0.001, εHF = 0.86.

Mean RTs (±the standard errors) in ascending order for the five types of movements. Below each bar is an example of a movement from that condition. *p < 0.05, **p < 0.01
Facilitation for symmetric amplitudes or symmetric target locations
Post hoc comparisons were used to analyse the potential RT savings for movements with one symmetric parameter (and two asymmetric parameters) compared to movements with all asymmetric parameters. There were significant RT savings for movements with symmetric amplitudes (SaT, −13.0 ± 2.8 ms, p < 0.001) and for movements with symmetric target locations (SAt, −5.9 ± 1.8 ms, p = 0.021). There were no significant RT savings for movements with symmetric starting locations (sAT, −7.5 ± 2.9 ms, p = 0.143). These results suggested that movement preparation was facilitated when movement amplitudes or target locations were symmetric.
Relative savings for symmetric starting locations, amplitudes, and target locations
RTs of the three movements with one symmetric parameter were not significantly different from each other. RTs for movements with symmetric amplitudes were comparable to RTs for movements with symmetric starting locations, p = 0.364. RTs for movements with symmetric starting locations were comparable to RTs for symmetric target locations, p = 1.000, and RTs for symmetric amplitudes were comparable to RTs for symmetric target locations, p = 0.106. These results, in isolation, suggested that the relative size of facilitation during movement preparation did not differ for symmetric starting locations, amplitudes, or target locations. However, this must be integrated with the results of the RT savings and costs to appreciate the contributions from each parameter.
Interference for movements with asymmetric amplitudes
There were significant costs for movements with asymmetric amplitudes and target locations (sAT, 11.8 ± 3.5 ms; p = 0.019) and for movements with asymmetric starting locations and amplitudes (SAt, 13.3 ± 2.5 ms; p < 0.001). The one common parameter for these two types of movements was asymmetric amplitudes, so movement amplitude may be the most important parameter for RT costs. In fact, there were no significant RT costs for movements with symmetric amplitudes even when the starting and target locations were asymmetric (SaT, 6.3 ± 2.5 ms, p = 0.176). These results suggested that movement preparation suffers interference when movement amplitudes are asymmetric.
Movement amplitudes determined movement times
Mean movement times for all movements are shown in Fig. 4. One visible difference was that movements with long amplitudes had longer movement times than movements with short amplitudes. This occurred for movements with symmetric amplitudes (Long–Long vs. Short–Short) and to a lesser degree for movements with asymmetric amplitudes (Long movements in Long-Short and Short-Long vs. Short movements in Long-Short and Short-Long). These differences were statistically confirmed with a significant main effect of Movement Amplitude, F(1,29) = 386, p < 0.001, that showed that long amplitude movements had longer movement times (155 ± 6.5 ms) than short amplitude movements (120 ± 5.9 ms).

Mean movement times (±the standard errors) for each movement type and amplitude
There was also a significant main effect of Movement Type, F(4,116) = 16, p < 0.001, ε HF = 0.81, and a significant Movement Type by Movement Amplitude interaction, F(4,116) = 22, p < 0.001, ε GG = 0.56. The interaction was analysed with simple main effects on Movement Type (Fig. 5). Long (149 ± 6.3 ms) and short (106 ± 5.0 ms) amplitude movements with all symmetric parameters were the baseline movement times for comparisons to the four conditions with asymmetric parameters (SaT, sAT, SAt, SAT).

Mean movement times (±the standard errors) for the long and short movement amplitudes of the five movement types
First, we compared the movement times of long amplitude movements. Movements with symmetric amplitudes and movements with symmetric starting locations had long amplitude movement times (156 ± 7.3, 155 ± 6.4 ms) that were comparable to movements with all symmetric parameters, p = 0.306, p = 0.341. Long amplitude movement times of movements with symmetric target locations (157 ± 6.4 ms) and movements with all asymmetric parameters (160 ± 7.3 ms) were significantly longer than movements with all symmetric parameters, p = 0.030, p = 0.028, but they were comparable to movements with symmetric amplitudes and movements with symmetric starting locations, ps > 0.617.
Next, we compared the movement times of short amplitude movements. Movements with symmetric amplitudes had short amplitude movement times (109 ± 5.3 ms) that were comparable to movements with all symmetric parameters, p = 0.620. Short amplitude movement times of movements with symmetric starting locations (126 ± 6.7 ms), symmetric target locations (125 ± 6.4 ms), and all parameters asymmetric (133 ± 7.6 ms) were significantly longer than movements with all parameters symmetric and movements with symmetric amplitudes, ps > 0.001. In other words, when paired with long amplitude movements, short amplitude movements showed the typical pattern of partial temporal assimilation to the long amplitude movement times (Kelso et al., 1979; Marteniuk, MacKenzie & Baba, 1984). Overall, these results suggested that the movement times were determined by the movement amplitudes and not by the symmetry/asymmetry of the starting or target locations.
Discussion
The goals of this study were to determine which symmetric movement parameters facilitate the preparation of bimanual movements and to compare the size of the facilitation for different parameters. There were significant RT savings for movements with symmetric amplitudes and movements with symmetric target locations compared to movements with all asymmetric parameters. This pattern of savings suggests that movement preparation is facilitated when movement amplitudes or target locations are symmetric. The analyses of RT costs showed significant costs for all movements with asymmetric amplitudes compared to movements with all symmetric parameters. These costs suggest that movement preparation suffers interference when movement amplitudes are asymmetric.
How do these results compare to those from previous studies? Heuer and Klein, (2006a) could not fully analyse the interaction between movement amplitudes and target locations due to a confound in the number of movement choices. However, they did compare conditions with a balanced number of movement choices in an attempt to partially separate the effects of amplitudes and target locations. In one of these comparisons, they found significantly longer RTs for a group of movements that included movements with all asymmetric parameters (SAT) and movements with asymmetric amplitudes and target locations (sAT) compared to movements with all symmetric parameters (sat). Movements with all asymmetric parameters were tested in simple reaction conditions, movements with asymmetric amplitudes and target locations were tested in 4-choice RT conditions, and movements with all symmetric parameters were tested in either simple or 4-choice conditions. We have previously shown comparable RTs for all bimanual movements in simple RT conditions, regardless of the symmetry or asymmetry of the movement parameters (Blinch et al., 2014). Therefore, the significant difference found by Heuer and Klein, (2006a) was likely caused by RT differences in the 4-choice conditions. If we omit movements in simple RT conditions from the comparison by Heuer and Klein, then their result becomes longer RTs for movements with asymmetric amplitudes and target locations than movements with all symmetric parameters. In other words, our interpretation of Heuer and Klein’s result is RT costs for movements with asymmetric amplitudes and target locations. The present study also found RT costs for the same movements. Therefore, both studies suggest that movement preparation suffers interference when movement amplitudes and target locations are asymmetric.
The results by Weigelt, (2007) can also be compared to our results, as both studies compared movements with one symmetric parameter to movements with all asymmetric parameters (RT savings). A common finding was RT savings for movements with symmetric target locations (SAt). This comparison in Weigelt’s study was confounded by the use of same or different symbolic cues, as RTs are shorter when two of the same symbolic cues are used compared to two different cues (Diedrichsen et al., 2001). Movements with symmetric targets had two of the same symbolic cues and movements with all asymmetric parameters had two different cues. Thus, it is not surprising that movements with symmetric targets (two identical symbolic cues) had RTs that were 25 ms shorter than movements with all asymmetric parameters (two different symbolic cues). This confound was eliminated in our study using direct cues. The RT savings for symmetric targets were much smaller in this study (−5.9 ± 1.8 ms) than in Weigelt’s study, suggesting that the cost to translate two different symbolic cues produced most of the difference between movement with symmetric targets and movements with all asymmetric parameters. However, the small but significant RT savings for symmetric targets that we found in the present study suggest that the spatial alignment of the targets does matter. The larger savings in Weigelt’s study were likely caused by a small savings for symmetric targets and a large savings for two identical symbolic cues.
Weigelt, (2007) also compared movements with symmetric amplitudes (SaT) and movements with symmetric starting locations (sAT) to movements with all asymmetric parameters (SAT). These comparisons were not confounded by the use of identical or different symbolic cues because all these movements had two different symbolic cues. For movements with symmetric amplitudes, Weigelt found a non-significant difference in RTs, while we found significant savings. The presence of this significant effect in our study, but not in Weigelt’s study, may be due to increased statistical power afforded by our study’s larger sample size (30 participants vs. 10). Another explanation is that symbolic cues cause longer RTs than direct cues (Diedrichsen et al., 2001), and small differences in RTs between conditions could be masked by longer RTs.
In contrast to our present results, Weigelt, (2007) found unexpected RT costs for movements with symmetric starting locations. He argued that the same starting locations might facilitate assimilation between the arms and increase interference during preparation. The present study found non-significant RT savings, and not costs, for movements with symmetric starting locations.
In a recent study (Blinch et al., 2014), we used the subtraction method (Donders, 1969) to isolate the costs for movements with symmetric starting locations and asymmetric amplitudes and target locations (sAT) to processes that are unique to choice RT. These processes include target discrimination, response selection, and response programming, and evidence from that study favoured response programming as the source of the interference during movement preparation. If we assume that bimanual interference occurs primarily during response programming, then the interference may be caused by concurrently programming two unimanual movements with asymmetric parameters (a bimanual asymmetric movement). In support of this hypothesis for interference during concurrent programming, we found that RT costs were eliminated when programming of the two arms was temporally separated (Blinch, Franks & Chua, 2013).
Our previous studies (Blinch et al., 2013, 2014) suggest that RT costs for directly cued asymmetric bimanual movements are caused by interference during response programming. So what do the current results suggest about the response programming of bimanual movements? The RT savings suggest that response programming considers the movement amplitudes and target locations. Movement preparation was facilitated when either amplitudes or target locations were symmetric. Starting locations, in contrast, did not consistently facilitate movement preparation. Starting locations may have had minimal impact on movement preparation because they were always known in advance of the imperative stimulus and could potentially be accounted for ahead of the upcoming movement. We have previously shown that the RT savings for bimanual movements with symmetric amplitudes and target locations are eliminated when those movement parameters are known in advance (i.e. in simple RT conditions; Blinch et al., 2014). In the present study, knowing the starting locations before the imperative stimuli may have reduced or eliminated any RT savings for symmetric starting locations.
From our results, we can also examine if the RT savings for movement amplitudes, starting locations, and target locations are additive. Recall that the RT savings were −13.0 ± 2.8 ms for symmetric amplitudes, −7.5 ± 2.9 for symmetric starting locations, and −5.9 ± 1.8 ms for symmetric target locations. Adding these savings together gives a summed RT savings of −26.4 ± 6.0 ms. In comparison, the RT savings for movements with all symmetric parameters compared to movements with all asymmetric parameters (sat–SAT) were –19.2 ± 2.8 ms, which was not significantly different from the summed savings, t(29) = 1.4, p = 0.164, Cohen’s d = 0.28 (calculated with the pooled standard deviation). As the individual savings for amplitudes and targets summed to be similar to the savings for all symmetric parameters, the individual savings may be additive. The additivity of these savings suggests that the savings for symmetric movement amplitudes and target locations occur during separate preparation processes (Sternberg, 1969).
The RT costs suggest that response programming considers the movement amplitudes, as movement preparation suffered interference with asymmetric amplitudes. This occurred for movements with asymmetric amplitudes and either asymmetric target locations or asymmetric starting locations (sAT or SAt). When the amplitudes were symmetric, the RT costs were not significant even when starting and target locations were asymmetric (SaT). Movement amplitude is also the primary determinant of the duration of movement execution; specifically, movement times were determined by the movement amplitudes and not by the symmetry/asymmetry of the starting or target locations.
Taken together, the RT costs and RT savings analyses suggest that movement amplitudes and target locations appear to be more important parameters for movement preparation than starting locations. It is interesting that many of the studies that have shown facilitation for bimanual symmetric movements (sat) compared to bimanual asymmetric movements (sAT) had symmetric starting locations for both types of movements (Blinch et al., 2014; Diedrichsen et al., 2006; Heuer & Klein, 2006b; Stelmach et al., 1988; Weigelt & Cardoso de Oliveira, 2003). These studies manipulated what we now believe are the more important parameters: movement amplitudes and target locations.
Previous research has argued that response programming of bimanual movements is facilitated when movement amplitudes are symmetric (Heuer, 1986; Stelmach et al., 1988). When movement amplitudes are asymmetric there may be cross-talk between concurrent processes of amplitude specification that causes transient interference (Heuer, Spijkers, Kleinsorge, van der Loo, & Steglich, 1998). This cross-talk is overcome and asymmetric amplitudes are specified at the cost of greater processing demands on response programming. It is possible that the contribution of target locations to response programming is similar to the contribution from movement amplitudes. When the target locations are symmetric, the same location can be coded for both arms and response programming is facilitated. When the target locations are asymmetric, there may be cross-talk between concurrent processes of target location specification (Heuer, 1993).
A fascinating aspect of the costs for bimanual asymmetric movements is that they can be affected by how the task is conceptualised by the participants [reviewed by Swinnen and Wenderoth, (2004)]. Various forms of bimanual interference have been reduced, or even eliminated, when a difficult bimanual movement is conceptualised as a single unified movement. Examples of unified bimanual movements that decrease interference are when a single goal is shared by the two arms (Franz, Zelaznik, Swinnen, & Walter, 2001), when a single stimulus represents that position of both arms (Kovacs, Buchanan, & Shea, 2009; Swinnen, Lee, Verschueren, Serrien, & Bogaerds, 1997), or both (White & Diedrichsen, 2010).
It was recently shown that the costs for bimanual reaching movements with asymmetric movement amplitudes and target locations, movements that were similar to the present experiment, were eliminated by two manipulations that encourage bimanual unification (Franz & McCormick, 2010). The first manipulation was a change to the target stimuli; connecting the two, circular targets with a line (resembling a dumbbell) virtually abolished the RT costs for bimanual asymmetric movements. RT costs were also abolished when the instructions before each trial were changed from encouraging separate representations of each arm (move one hand and move the other hand to their respective targets) to a unified representation (move both hands to their respective targets).In the present experiment, the bimanual movements were likely conceptualised as two separate unimanual movements as the targets were visually disconnected and the instructions did not use unified language (the targets were illustrated during the instructions by gesturing). Our results of bimanual interference for asymmetric movement amplitude or target locations remain valid, but it is important to appreciate that the magnitude of any form of bimanual interference will likely depend on how the task is conceptualised.
In summary, we investigated the influence of movement amplitudes, target locations, and starting locations on the preparation of bimanual reaching movements. Our findings suggest that movement preparation, or more specifically, response programming, is facilitated when movement amplitudes or target locations are symmetric. Conversely, response programming suffers interference when the movement amplitudes are asymmetric. The relative importance of these parameters to response programming, from most to least important, is movement amplitudes, target locations, and then starting locations. Interference with asymmetric amplitudes or target locations may be caused by cross-talk between concurrent processes of parameter specification. There are, of course, other parameters we did not investigate that may also have an impact on movement preparation, such as movement direction (Heuer, 2006; Heuer & Klein 2006c). Future experiment could investigate these parameters and determine where they fit in the apparent hierarchy of parameters that influence the programming of bimanual movements.
Notes
Statistically, it does not make a difference whether we analyse the RT savings [(SaT, sAT, SAt)—SAT] or the RTs (SaT, sAT, SAt). Subtracting SAT from the RTs to calculate the RT savings affects the participant variance but neither the treatment variance nor the error variance. The results of these analyses are identical because repeated measures ANOVAs disregard the participant variance. The variances that are used are the treatment and error variances and these are identical in both calculations. Therefore, analysis of the RTs allowed us to make inferences about the relative RT savings.
References
Blinch, J., Cameron, B. D., Cressman, E. K., Franks, I. M., Carpenter, M. G., & Chua, R. (2014). Comparing movement preparation of unimanual, bimanual symmetric, and bimanual asymmetric movements. Experimental Brain Research, 232(3), 947–955.
Blinch, J., Franks, I. M., Chua, R. (2013). Eliminating the preparation cost for bimanual asymmetric movements. Poster presented at the biannual meeting of Progress in Motor Control, Montreal, QC.
Diedrichsen, J., Grafton, S., Albert, N., Hazeltine, E., & Ivry, R. B. (2006). Goal-selection and movement-related conflict during bimanual reaching movements. Cerebral Cortex, 16(12), 1729–1738.
Diedrichsen, J., Hazeltine, E., Kennerley, S., & Ivry, R. B. (2001). Moving to directly cued locations abolishes spatial interference during bimanual actions. Psychological Science, 12(6), 493–498.
Donders, F. C. (1969). On the speed of mental processes. Acta Psychologica, 30, 412–431.
Favilla, M. (1996). Reaching movements: programming time course is independent of choice number. NeuroReport, 7(15–17), 2629–2634.
Franz, E. A., & McCormick, R. (2010). Conceptual unifying constraints override sensorimotor interference during anticipatory control of bimanual actions. Experimental Brain Research, 205(2), 273–282.
Franz, E. A., Zelaznik, H. N., Swinnen, S. S., & Walter, C. (2001). Spatial conceptual influences on the coordination of bimanual actions: when a dual task becomes a single task. Journal of Motor Behavior, 33(1), 103–112.
Heuer, H. (1986). Intermanual interactions during programming of finger movements: Transient effects of ‘homologous coupling’. In H. Heuer & C. Fromm (Eds.), Generation and modulation of action patterns (pp. 87–101). Berlin: Springer.
Heuer, H. (1993). Structural constraints on bimanual movements. Psychological Research, 55(2), 83–98.
Heuer, H. (2006). Simultaneous specification of amplitudes and directions of bimanual reversal movements. Journal of Motor Behaviour, 38(4), 285–298.
Heuer, H., & Klein, W. (2006a). Intermanual interactions related to movement amplitudes and endpoint locations. Journal of Motor Behavior, 38(2), 126–138.
Heuer, H., & Klein, W. (2006b). The influence of movement cues on intermanual interactions. Psychological Research, 70(4), 229–244.
Heuer, H., & Klein, W. (2006c). The modulation of intermanual interactions during the specification of the directions of bimanual movements. Experimental Brain Research, 169(2), 162–181.
Heuer, H., Spijkers, W., Kleinsorge, T., van der Loo, H., & Steglich, C. (1998). The time course of cross-talk during the simultaneous specification of movement amplitudes. Experimental Brain Research, 118(3), 381–392.
Hick, W. E. (1952). On the rate of gain of information. Quarterly Journal of Experimental Psychology, 4(1), 11–26.
Huynh, H., & Feldt, L. S. (1976). Estimation of the box correction for degrees of freedom from sample data in randomized block and split-plot designs. Journal of Educational Statistics, 1(1), 69–82.
Hyman, R. (1953). Stimulus information as a determinant of reaction time. Journal of Experimental Psychology, 45(3), 188–196.
Kelso, J. A., Southard, D. L., & Goodman, D. (1979). On the coordination of two-handed movements. Journal of Experimental Psychology: Human Perception and Performance, 5(2), 229–238.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2009). Bimanual 1:1 with 90 degrees continuous relative phase: difficult or easy! Experimental Brain Research, 193(1), 129–136.
Marteniuk, R. G., MacKenzie, C. L., & Baba, D. M. (1984). Bimanual movement control: Information processing and interaction effects. Quarterly Journal of Experimental Psychology, 36(A), 335–365.
Spijkers, W., Heuer, H., Kleinsorge, T., & van der Loo, H. (1997). Preparation of bimanual movements with same and different amplitudes: specification interference as revealed by reaction time. Acta Psychologica, 96(3), 207–227.
Stelmach, G. E., Amrhein, P. C., & Goggin, N. L. (1988). Age differences in bimanual coordination. Journal of Gerontology, 43(1), 18–23.
Sternberg, S. (1969). The discovery of processing stages: Extension of Donders’ method. In W. G. Koster (Ed.), Attention and Performance II (pp. 276-315). Amsterdam: North-Holland.
Swinnen, S. P., Lee, T. D., Verschueren, S., Serrien, D. J., & Bogaerds, H. (1997). Interlimb coordination: learning and transfer under different feedback conditions. Human Movement Science, 16(6), 749–785.
Swinnen, S. P., & Wenderoth, N. (2004). Two hands, one brain: cognitive neuroscience of bimanual skill. Trends in Cognitive Sciences, 8(1), 18–25.
Weigelt, M. (2007). Re-examining structural constraints on the initiation of bimanual movements: the roles of starting locations, movement amplitudes, and target locations. Human Movement Science, 26(2), 212–225.
Weigelt, C., & Cardoso de Oliveira, S. (2003). Visuomotor transformations affect bimanual coupling. Experimental Brain Research, 148(4), 439–450.
Wenderoth, N., & Weigelt, M. (2009). Visual cues influence motor coordination: behavioral results and potential neural mechanisms mediating perception-action coupling and response selection. Progress in Brain Research, 174, 179–188.
White, O., & Diedrichsen, J. (2010). Responsibility assignment in redundant systems. Current Biology, 20(14), 1290–1295.
Wright, C. E., Marino, V. F., Belovsky, S. A., & Chubb, C. (2007). Visually guided, aimed movements can be unaffected by stimulus-response uncertainty. Experimental Brain Research, 179(3), 475–496.
Acknowledgements
We would like to thank Associate Editor Peter Q Pfordresher and two anonymous reviewers for their insightful critiques. We would also like to thank Brandon Rasman, Kathryn Clark, and Curtis May for their help with participant recruitment and data collection. The Natural Sciences and Engineering Research Council of Canada supported this research with a postgraduate scholarship awarded to Jarrod Blinch and a discovery grant awarded to Romeo Chua. Brendan Cameron was supported by a Juan de la Cierva postdoctoral fellowship (JCI-2011-09664) and a Marie Curie CIG (618407). The views expressed here reflect those of the authors only and not those of the funders.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Blinch, J., Cameron, B.D., Franks, I.M. et al. Facilitation and interference during the preparation of bimanual movements: contributions from starting locations, movement amplitudes, and target locations. Psychological Research 79, 978–988 (2015). https://doi.org/10.1007/s00426-014-0624-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-014-0624-y