
Abstract
Phenolic compounds are generated by the secondary metabolism of plants and have been associated with antibacterial properties. Among bacteria affecting human health, Helicobacter pylori has been associated with gastric cancer. There is limited information about the effect of individual or grouped phenolics on H. pylori growth. Previous studies have evaluated the effect of phenolics being part of highly complex food or beverage matrices. The aim of this work was to evaluate the in vitro antibacterial effect of kaempferol and (−)-epicatechin, both individually and combined, on H. pylori in liquid and solid cultures and in co-cultures with AGS human gastric carcinoma cells. Bacterial viability tests were performed in liquid cultures with subsequent CFU/mL counting and in solid cultures by measuring inhibition haloes. Kinetic curves of bacterial growth inhibition in the presence of those phenolics, and the protective effect of (−)-epicatechin on AGS cells against H. pylori infection were characterized. (−)-Epicatechin and kaempferol displayed antibacterial activities, being (−)-epicatechin more effective than kaempferol. After the combined application of both phenols, a synergistic effect of kaempferol plus low but not high doses of (−)-epicatechin was observed. Finally, (−)-epicatechin yielded protection to AGS cells against H. pylori infection. (−)-Epicatechin and kaempferol, both individually and combined, have antibacterial properties and protective effect on H. pylori infection.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Helicobacter pylori is a neutralophilic Gram-negative spiral-shaped bacterium that colonizes the human stomach [1]. Several studies have shown that H. pylori is one of the most prevalent infections throughout the world [2]. Approximately 80 % of the infected population is asymptomatic and only a small but significant fraction of the colonized patients present gastric or duodenal ulcers, mucosa-associated lymphoid tissue lymphoma (MALT lymphoma) or gastric cancer [1–3]. H. pylori infection represents the major risk factor for gastric cancer which accounts annually for at least 738,000 deaths [3, 4]. It has been estimated that H. pylori increases ten times the probability to develop gastric cancer [5]. Eradication therapies for H. pylori involve the use of a proton-pump inhibitor (PPI), tetracycline, amoxicillin, clarithromycin or metronidazole, in a triple therapy [6]. However, the increased prevalence of antibiotic resistance makes H. pylori eradication not always successful. A recent study in Latin American countries showed that antibiotic resistance rates vary significantly by drug and by country, but not by year of sample collection [7]. However, such observation contrasts with another study from outside the Latin America region that showed rising resistance rates to certain antibiotics over time [6]. Metronidazole can reach 100 % resistance [8], whereas 30 and 20 % of all isolates are resistant to tetracycline and clarithromycin, respectively [9]. Considering that eradication therapies based on the use of drugs that may be unavailable in some geographic areas or that may produce undesirable secondary effects can be costly and ineffective, the search for new drugs and the development of alternative therapies has become critical [10]. Plants are attractive sources of polyphenols, a family of compounds showing promising results in the treatment of a variety of diseases. Particularly, several studies have shown that phenolic compounds found in cranberries, green tea, apple and wine, and affect H. pylori viability [11–19]. All those studies were based on the analysis of the effect of polyphenols from complex plant extracts. In that regard, limited information exists about the effect of pure polyphenols on H. pylori [20]. Kaempferol and (−)-epicatechin, two phenolic compounds, are widely distributed among plants [21, 22]. Not much is known about the effect of either pure kaempferol or pure (−)-epicatechin, or about any eventual synergy or interference between them, on H. pylori growth. The aim of this study was to compare the antimicrobial effect of kaempferol and (−)-epicatechin, alone or in combination, against H. pylori viability. Also, an eventual secondary protective effect of (−)-epicatechin on H. pylori-infected epithelial cells was tested.
Materials and methods
Materials
Kaempferol and (−)-epicatechin standards (Sigma-Aldrich, Santiago, Chile), cellulose acetate filters (Asahi Glass, Tokyo, Japan), bacto agar (JT Baker, Mexico), dimethyl sulfoxide (DMSO) (Winkler, Santiago, Chile), horse serum (Thermo Scientific HyClone, Utah, USA), antibiotic supplements Dent and Vitox (Oxoid, Hampshire, England) and pro-analysis grade chemicals (Merck Ltd., Santiago, Chile) were purchased.
Polyphenol solutions
Stock solutions of kaempferol and (−)-epicatechin (50 mg/mL) were prepared by dissolving each polyphenol in 100 % DMSO and 50 % methanol, respectively. Both solutions were sterilized by filtration through cellulose acetate filters (0.2 µm pore size; 25 mm diameter).
Bacterial strains and growth conditions
Two H. pylori strains were used in this study: ATCC 43504 (NCTC 11637), which was first isolated from the gastric antrum of an Australian patient, and ATCC 700392 (26695), which was isolated from a patient with gastritis in the UK. Frozen stocks of H. pylori were recovered and routinely grown for 48 h at 37 °C in 5.5 % CO2 and 70–80 % humidity on trypticase soy agar (TSA) (Becton–Dickinson, Sparks, MD 21152, USA) with 0.4 % H. pylori selective supplement Dent (Oxoid Basingstoke, Hampshire, England), 0.3 % Isovitalex (Oxoid) and 5 % horse serum [23–25]. For growth experiments in liquid cultures, cells were grown in trypticase soy broth (TSB) (Becton–Dickinson) with 5 % horse serum supplemented with Isovitalex and Dent. Bacteria were first grown at pH 7.0 to an optical density of 0.6–1.0 U at 600 nm (OD600) and subsequently diluted to a starting OD600 of 0.05 U. To measure the growth of H. pylori in liquid medium, a serial dilution of the culture medium was prepared, aliquots of the various dilutions were plated on trypticase soy agar (TSA) plates, and the number of colonies (CFU) was determined [26].
Protective effect of phenolics on gastric cells against H. pylori
AGS gastric cancer cells (ATCC CRL 1739) were cultured as reported elsewhere [27]. For infection assays, AGS host cells were infected at a MOI (multiplicity of infection) of 100 (bacteria to gastric cell ratio) as described by Valenzuela et al. [27]. In a parallel assay, gastric cells were treated with (−)-epicatechin prior to infection. AGS cell viability was determined by measuring the mitochondrial reductase activity (MTT assay) according to del Campo et al. [28].
H. pylori growth assay in liquid medium
Helicobacter pylori (3 × 107 cells/mL) was inoculated in 5 mL of TSB supplemented with a range of concentrations (0.0, 0.2, 0.4, 0.6, 0.8 and 1.0 mg/mL) of kaempferol or (−)-epicatechin. After incubation at 37 °C for 48 h with constant shaking at 250 rpm in a controlled atmosphere incubator (5.5 % CO2 and 70 % relative humidity), bacterial growth was determined by turbidimetry at 600 nm or by counting colony forming units on TSA plates [29].
H. pylori viability assay
From each of the experimental culture tubes described in the previous section, 100 µL of aliquots was taken at the end of the incubation period to prepare serial dilutions in PBS. Aliquots of 10 µL from each of these dilutions were plated on TSA and incubated for 48 h at 37 °C [20]. The number of colony forming units per mL (CFU/mL) corresponding to each experimental condition was determined.
Inhibition kinetics of bacterial growth
The assay was conducted basically as described by Romero et al. [30] with minor modifications. Briefly, 3 × 107 CFU/mL were inoculated into Eppendorf tubes containing 1 mL of TSB medium supplemented with 1 mg/mL of kaempferol or (−)-epicatechin and incubated at 37 °C. From this mixture, 100 µL of aliquots was removed at 0, 15, 30, 45, 60 and 90 min. The numbers of viable cells at various time points were determined as described above.
Inhibition halo test on agar plates
The procedure was performed as described by Rodríguez et al. [31]. Three-millimeter-diameter wells were made in TSA plates, and 30 µL of a series of kaempferol or (−)-epicatechin solutions (corresponding to 0–1 mg of the polyphenol) was deposited in individual wells. Then, 100 µL of H. pylori suspension containing 3 × 107 cells/mL was evenly spread over the plates with a metal handle loop. After 48 h of incubation at 37 °C, the diameter of each growth inhibition halo was determined.
Effect of combining kaempferol and (−)-epicatechin on H. pylori growth
Helicobacter pylori strains were grown on TSA plates with a series of wells containing a fixed concentration of kaempferol (0.4 mg/well) plus increasing doses of (−)-epicatechin (0.0, 0.05, 0.1, 0.2, 0.25 and 0.4 mg/well). After 48 h of incubation at 37 °C, the diameter of each growth inhibition halo was measured.
Statistical analysis
Analysis of variance (ANOVA) and Tukey’s tests were used to evaluate differences.
Results
Antibacterial activity of (−)-epicatechin against H. pylori
H. pylori growth in (−)-epicatechin-supplemented liquid medium
Growth of H. pylori strains 43504 and 26695 in the presence of (−)-epicatechin was evaluated in TBS medium supplemented with a range of (−)-epicatechin concentrations over a 48-h incubation period. After this time, CFU/mL of each cell suspension was determined. As shown in Fig. 1, the CFU/mL parameter decreased when the medium was supplemented with a range of concentrations of (−)-epicatechin. Full growth inhibition was observed with (−)-epicatechin concentrations of 0.8 and 1.0 mg/mL for H. pylori 43504 and 26695 strains, respectively. In this assay, the minimum inhibitory concentration (MIC) of (−)-epicatechin to produce 99 % of H. pylori growth inhibition was 0.6 mg/mL.

Concentration-dependent bactericidal effect of (−)-epicatechin on H. pylori strains in liquid medium. Aliquots of culture medium taken after 48 h of culturing H pylori 43504 and 26695 strains in the presence of a range of (−)-epicatechin concentrations are tested by counting CFU. The figure is representative of three independent experiments. Growth inhibition in both H. pylori strains is significant (Tukey’s test, p ≤ 0.05) over all the range of tested (−)-epicatechin concentrations
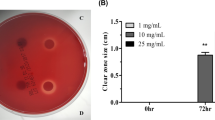
Inhibition halo test for (−)-epicatechin
Bacteria were cultured for 48 h on TSA plates with a series of wells containing a range of fixed amounts of (−)-epicatechin. As shown in Fig. 2, diameters of the growth inhibition zones around the wells increased gradually with the content of (−)-epicatechin in the range from 0 mm for the control condition (only solvent) to 15 mm in the presence of 1.0 mg of (−)-epicatechin. Compared to the control, growth inhibition halos were significantly larger (p < 0.05) with amounts of (−)-epicatechin equal or greater than 0.4 mg/well.

Inhibitory effect of (−)-epicathechin on H. pylori growth in semisolid agar medium. Inhibition halo test on H. pylori strains exposed to (−)-epicatechin over the range 0–1 mg/well showed a progressive inhibitory effect. Values are mean ± SD (n = 3). No inter-strain differences are observed (Tukey’s test, p ≥ 0.05). The ordinate represents diameter of the inhibition zone around well
Kinetics of bacterial growth inhibition by (−)-epicatechin
Cultures of H. pylori 26695 and 43504 strains in TBS medium were supplemented with 1.5 mg/mL of (−)-epicatechin at time zero and at various time points; thereafter (up to 90 min), aliquots of the cell culture were seeded on TSA plates to determine the CFU/mL parameter. As shown in Fig. 3, after 90 min of exposure to (−)-epicatechin the CFU/mL values for H. pylori 43504 decreased from 3 × 108 at time zero to 9.5 × 107, whereas for H. pylori 26695 such parameter fell from 3 × 108 to 3 × 106 after the same treatment. Decreases in viabilities of both H. pylori strains became statistically significant (p < 0.05) after 60 min of incubation in the presence of (−)-epicatechin. Altogether, (−)-epicatechin provoked a dose-dependent decrease in the CFU/mL values and a dose-dependent increase in the diameters of the inhibition halos on agar plate cultures with both bacterial strains. Those effects are also consistent with the observation that the CFU/mL value decreased with increasing times of exposure of the H. pylori strains to such polyphenol. Accordingly, (−)-epicatechin has a significant growth-inhibitory effect and antimicrobial activity on H. pylori, thus having a major influence on the viability of the microorganism.

Kinetics of growth inhibition of H. pylori by (−)-epicatechin. Liquid cultures of H pylori 43504 and 26695 strains are combined with 1 mg/mL (−)-epicatechin, and at the times indicated on the abscissa aliquots are taken and tested by counting CFU. Values are mean ± SD (n = 3). Growth inhibition in both H. pylori strains is directly influenced (Tukey’s test, p ≤ 0.05) by the exposure time to the polyphenol over the range tested
Antibacterial activity of kaempferol against H. pylori
H. pylori growth in kaempferol-supplemented liquid medium
Helicobacter pylori strains 26695 and 43504 were incubated for 48 h in TBS liquid culture medium containing a range of kaempferol concentrations (0 to 1 mg/mL) (Fig. 4). The CFU/mL values ranged from 5.2 × 108 (43504 strain) or 3.5 × 107 (26695 strain) in the control condition to 0 CFU/mL in both cultures at 1.0 mg/mL of kaempferol. Both decreases were statistically significant (p < 0.05).

Concentration-dependent bactericidal effect of kaempferol on H. pylori strains in liquid medium. The figure is representative of three independent experiments. Experimental conditions and results are identical to those described in Fig. 1 caption, except the polyphenol
Inhibition halo test for kaempferol
Bacteria were cultured for 48 h on TSA well plates containing a series of different amounts of kaempferol. As shown in Fig. 5, diameters of the growth inhibition halos around the wells increased gradually with the increasing content of kaempferol in the range from 0 mm for the control condition (only solvent) up to 15 mm for the presence of 1.0 mg of kaempferol. Growth inhibition halos were significantly larger than control (p < 0.05) with amounts of kaempferol equal or greater than 0.6 mg/well.

Inhibitory effect of kaempferol on H. pylori growth in semisolid agar medium. Experimental conditions and results are identical to those described in Fig. 2 caption, except the polyphenol. Values are mean ± SD (n = 3)
Kinetics of bacterial growth inhibition by kaempferol
In a similar experiment to the one described for (−)-epicatechin, the CFU/mL parameter was determined at various times after supplementation of the culture medium with 1.5 mg/mL of kaempferol. With both H. pylori strains, an increased viability was observed after 30 min of exposure to kaempferol. Longer times of exposure resulted in a statistically significant decrease in the CFU/mL value. Thus, after 90 min of exposure to kaempferol the CFU/mL value was reduced from 3 × 109 at time zero to 3 × 107 (Fig. 6). The inhibitory effect of kaempferol on H. pylori growth was found to be similar to the one observed for (−)-epicatechin. Furthermore, no statistically significant differences in the sensitivities of both H. pylori strains were observed. This observation suggests that the antibacterial effects of (−)-epicatechin and kaempferol are not dependent upon the bacterial strain.

Kinetics of growth inhibition of H. pylori by kaempferol. The study is identical to the one described under Fig. 3, excepting the polyphenol. Values are mean ± SD (n = 3). Although in this study growth of both H. pylori strains is influenced by the exposure time to the polyphenol, growth inhibition is found to be significant (Tukey’s test, p ≤ 0.05) only after at least 60 min of exposure. Both strains displayed a similar behavior
Effect of combining (−)-epicatechin and kaempferol on H. pylori growth
Helicobacter pylori 43504 was grown on TSA well plates with a series of either a fixed concentration of (−)-epicatechin (0.25 mg/well) or kaempferol (0.5 mg/mL) plus increasing doses of kaempferol or (−)-epicatechin, respectively. After 48 h of incubation at 37 °C CFU/mL, values were determined. Table 1 summarizes a factorial arrangement of treatments with doses of 0, 50, 100 and 150 µg/mL of each compound. CFU/mL values ranged from 5.4 × 107 (control) to 1.4 × 103 (50 µg/mL (−)-epicatechin plus 150 µg/mL kaempferol). Over the whole range of tested concentrations, increasing concentrations of kaempferol in the presence of a constant dose of (−)-epicatechin resulted in further reduction in bacteria growth. A similar trend was observed with varying doses of (−)-epicatechin and a fixed amount of kaempferol. Comparatively, at similar concentrations of the polyphenol, viability of the bacterium in the presence of kaempferol was somewhat greater than that observed in the presence of (−)-epicatechin. Such effect was amplified with increasing doses of kaempferol.
(−)-Epicatechin effect on a co-culture of AGS cells and H. pylori
To see how H. pylori infection affects viability of AGS cells, cultures of these cells were infected with different H. pylori MOIs. After incubation for 24 h, the mitochondrial reductase activity (MTT) was determined as an index of AGS cell viability. As shown in Fig. 7a, the higher the MOI the lower the viability of AGS cells so that at a MOI of 500 AGS cell viability was reduced to 40 %. On the other hand, viability of AGS cells in the presence of (−)-epicatechin over the range 100–500 µg/mL was unaffected or just marginally affected (Fig. 7b). To evaluate the protective effect of (−)-epicatechin on H. pylori-infected AGS cells, viability assays were conducted in liquid medium in the presence of either 250 or 500 µg/mL of the flavanol at a MOI of 500. As shown in Fig. 7c, viability of AGS cells decreased to 40 % at a MOI of 500 (compared to 100 % viability of uninfected AGS cells serving as control), while infection at a MOI of 500 in the presence of either 250 or 500 µg/mL (−)-epicatechin reduced AGS cell viability to 75–80 % only. These results are indicative of a protective effect of (−)-epicatechin on H. pylori-infected AGS cells.

Protective effect of (−)-epicatechin on AGS cell viability (MTT assay) during H. pylori infection. a AGS cell survival in DMEM medium over a range of H. pylori MOI. b AGS cell survival over a range of (−)-epicatechin concentrations. c Survival of AGS cells is measured when their infection by H. pylori at a MOI 500 is produced in the presence of (−)-epicatechin at concentrations (250–500 µg/mL) having no detrimental effect on AGS viability. Under both conditions, AGS cell viability (mean ± SD, n = 3) is significantly higher than that observed in the absence of the polyphenol (Tukey’s test, p ≤ 0.05). EC and MOI stand for (−)-epicatechin and multiplicity of infection by H. pylori, respectively
Discussion
Polyphenols, a highly abundant vegetal component of the human diet, have been associated with important sensory, taxonomic, nutritional and pharmacological properties of drinks and foods [32]. Several authors have reported on the bactericidal effect of polyphenol-rich extracts against a number of microorganisms [11, 13, 16, 18, 19, 30, 33]. Such observation has stimulated research on naturally occurring substances as potential alternatives or complements for the conventional treatment of infectious diseases [12–14, 18, 19]. H. pylori, a bacterium that colonizes the human gastric mucosa, has been associated with various pathological conditions, including peptic ulcer, intestinal lymphoma and gastric cancer (1–3). Anti-H. pylori effects have been observed in polyphenol extracts from almond skins [34], muscadine grape skins [35] and blackberry leaves [16], among others. A few studies aimed to identify particularly active anti-H. pylori components of those extracts have pointed recurrently to protocatechuic acid, catechin, epicatechin, ellagic acid, myricetin, quercetin, resveratrol, gallic acid, tannic acid and n-propyl gallate [20, 34, 36, 37]. In the present study, we evaluated quantitatively the individual and combined effects of kaempferol and (−)-epicatechin, two widely distributed components of plant extracts, on H. pylori growth in liquid and semisolid cultures. We have now shown that both polyphenols have a major growth-inhibitory effect on H. pylori.
Epicatechin displayed a concentration-dependent inhibitory effect on H. pylori growth as measured either in liquid culture medium (turbidimetry at 600 nm) or in semisolid agar plates (counting CFU and measuring growth inhibition halos). Such inhibitory effect was found to depend upon the time of exposure of the bacterium to the polyphenol. Using the same approaches, kaempferol, another member of the flavonol family of polyphenols, was also found to have growth-inhibitory effects against H. pylori. All those growth-inhibitory effects were identical in two widely used H. pylori strains (H. pylori 26695 and 43504). Altogether, on a per mole basis, kaempferol and epicatechin displayed similar growth-inhibitory effects on H. pylori.
Considering that both epicatechin and kaempferol showed marked inhibitory effects against H. pylori growth and that both polyphenols are usually part of the complex composition of foods, we additionally evaluated the effect of combining epicatechin and kaempferol on the bacterium growth in liquid culture medium. Thus, mixtures of low doses of epicatechin and kaempferol displayed additive antibacterial activity. Under these experimental conditions, such additive effect was partial because the effect of both polyphenols used in combination was usually less than the sum of the individual effects. Interestingly, at higher doses of epicatechin and kaempferol neither additive nor synergistic activity was observed. These observations contradict Yanagawa et al. [17] who claim that polyphenols in plant extracts are currently less effective antibacterial compounds than the corresponding pure compounds. Anyhow, the partially additive effect of kaempferol and epicatechin on the inhibition of H. pylori growth needs to be further studied. In this same regard, we have recently described that other two polyphenol compounds, catechin and gallic acid, display growth-inhibitory effects in H. pylori and that those effects are also partially additive [20].
According to the present observations, H. pylori seems to display a significantly higher sensitivity (tenfold) to both epicatechin and kaempferol when compared with E. coli, a common gut bacteria (data not shown). Thus, these observations would lend additional support to the hypothesis that these polyphenol compounds may have differential effects on different microorganisms [20, 30]. The fact that the size of the H. pylori genome is a third of the one of E. coli may also explain on a genetic ground its lower ability to counteract the antibacterial effects of epicatechin and kaempferol and probably other polyphenols.
On the other hand, several lines of evidence indicate that H. pylori promotes gastric carcinogenesis depending on the direct contact of live bacteria with human cells [1–3, 27], which would affect the viability of the latter ones. Considering that (−)-epicatechin and kaempferol have been found to display significant anti-H. pylori effects, in this study we designed an (−)-epicatechin protection assay using AGS human gastric carcinoma cells subjected to H. pylori infection. As shown in this study, AGS cell viability was markedly reduced when cultured in the presence of H. pylori. By contrast, when the assay was carried out in the presence of 250–500 μg/mL (−)-epicatechin, AGS cell viability was just marginally affected by H. pylori infection, thus showing indirectly the anti-H. pylori effect of the polyphenol.
Altogether, the whole set of in vitro observations suggest that both kaempferol and (−)-epicatechin display growth-inhibitory effects on H. pylori. It remains to be clarified whether these observations have an in vivo correlate considering that both small-sized polyphenols are efficiently absorbed by the jejunum and ileum, that they may be chemically modified in the small intestine and that they usually represent only a minor fraction of the total polyphenol content of the human diet in contrast with the abundance of non-absorbable more complex polyphenols [38, 39]. Anyhow, antibacterial activities of kaempferol and (−)-epicatechin against H. pylori may well become part of the growing list of beneficial effects of common polyphenol constituents of the human diet.
Conclusion
Under the conditions of this study, both (−)-epicatechin and kaempferol presented a significant inhibitory effect on H. pylori growth in a concentration-dependent manner. Both polyphenols affected growth of H. pylori 26695 and 43504 strains. At identical doses, the effect of (−)-epicatechin was higher than that of kaempferol. In liquid culture medium, at low doses both compounds showed additive antibacterial activity, while at higher doses neither additive nor synergistic activity was observed. Finally, (−)-epicatechin showed a dose-dependent protective role on AGS cell cultures against H. pylori infection.
References
Dunn B, Cohen H, Blaser M (1997) Helicobacter pylori. Clin Microbiol Rev 10:720–741
Eusebi LH, Zagari RM, Bazzoli F (2014) Epidemiology of Helicobacter pylori infection. Helicobacter 19:1–5
Nagani S (2012) Carcinoma of the stomach: a review of epidemiology, pathogenesis, molecular genetics and chemoprevention. World J Gastrointest Oncol 4:156–169
Parkin D (2000) Global cancer statistics in the year. Lancet Oncol 2:533–543
Yamaoka Y (2008) Helicobacter pylori—molecular genetics and cellular biology. Horizon Scientific Press, UK, p 261
O’Connor A, Vaira D, Gisbert JP, O’Morain C (2014) Treatment of Helicobacter pylori infection. Helicobacter 19:38–45
Camargo MC, García A, Riquelme A, Otero W, Camargo CA, Hernandez-García T, Candia R, Bruce MG, Rabkin CS (2014) The problem of Helicobacter pylori resistance to antibiotics: a systematic review in Latin America. Am J Gastroenterol 109(4):485–495
Aboderin OA, Abdu AR, Odetoyin B, Okeke IN, Lawal OO, Ndububa DA, Agbakwuru AE, Lamikanra A (2007) Antibiotic resistance of Helicobacter pylori from patients in Ile-Ife, South-west, Nigeria. Afr Health Sci 7(3):143–147
Vallejos C, Garrido L, Cáceres D, Madrid AM, Defilippi C, Defilippi CC, Toledo H (2007) Prevalence of metronidazole, clarithromycin and tetracycline resistance in Helicobacter pylori isolated from Chilean patients. Rev Med Chile 135:287–293
Bonacorsi C, Raddi MS, Carlos IZ, Sannomiya M, Vilegas W (2009) Anti-Helicobacter pylori activity and immunostimulatory effect of extracts from Byrsonima crassa Nied. (Malpighiaceae). BMC Complement Altern Med 9:2
Tombola F, Campello S, De Luca L, Ruggiero P, Del Giudice G, Papini E, Zoratti M (2003) Plant polyphenols inhibit VacA, a toxin secreted by the gastric pathogen Helicobacter pylori. FEBS Lett 543:184–189
Mabe K, Yamada M, Oguni I, Takahashi T (1999) In vitro and in vivo activities of tea catechins against Helicobacter pylori. Antimicrob Agents Chemother 43(7):1788–1791
Mahady G, Pendland S, Chadwick L (2003) Resveratrol and red wine extracts inhibit the growth of CagAþ strains of Helicobacter pylori in vitro. Am J Gastroenterol 98:1440–1441
Pastene P, Speisky H, García A, Moreno J, Troncoso M, Figueroa G (2010) In vitro and in vivo effects of apple peel polyphenols against Helicobacter pylori. J Agric Food Chem 58:7172–7179
Brown J, Huang G, Haley-Zitlin V, Jiang X (2009) Antibacterial effects of grape extracts on Helicobacter pylori. Appl Environ Microbiol 75:848–852
Martini S, D’Addario C, Colacevich A, Focardi S, Borghini F, Santucci A (2009) Figura N, Rossi C. Antimicrobial activity against Helicobacter pylori strains and antioxidant properties of blackberry leaves (Rubus ulmifolius) and isolated compounds. Int J Antimicrob Agents 34:50–59
Yanagawa Y, Yamamoto Y, Hara Y, Shimamura T (2003) A combination effect of epigallocatechin gallate, a major compound of green tea catechins, with antibiotics on Helicobacter pylori growth in vitro. Current Microbiol 47:244–249
Vattem D, Lin Y, Ghaedian R, Shetty K (2005) Cranberry synergies for dietary management of Helicobacter pylori infections. Process Biochem 40:1583–1592
Yahiro K, Shirasaka D, Tagashira M, Wada A, Morinaga N, Kuroda F, Choi O, Inoue M, Aoyama N, Ikeda M, Hirayama T, Moss J, Noda M (2005) Inhibitory effects of polyphenols on gastric injury by Helicobacter pylori VacA toxin. Helicobacter 10:231–239
Díaz-Gómez R, López-Solís R, Obreque-Slier E, Toledo-Araya H (2013) Comparative antibacterial effect of gallic acid and catechin against Helicobacter pylori. LWT Food Sci Technol 54:331–335
Obreque-Slier E, Peña-Neira A, López-Solís R, Zamora-Marín F, Ricardo da Silva J, Laureano O (2010) Comparative study of the phenolic composition of seeds and skins from Carménère and Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. J Agric Food Chemi 58:3591–3599
Monagas M, Bartolomé B, Gómez-Cordovés C (2005) Update knowledge about the presence of phenolic compounds in wine. Crit Rev Food Sci Nutr 42:85–118
Cerda O, Rivas A, Toledo H (2003) Helicobacter pylori strain ATCC 700392 encodes a methyl-accepting chemotaxis receptor protein (MCP) for arginine and sodium bicarbonate. FEMS Microbiol Letters 224:175–181
Toledo H, Valenzuela M, Rivas A, Jerez C (2002) Acid stress response in Helicobacter pylori. FEMS Microbiol Letters 213:67–72
Almarza O, Nuñez D, Toledo H (2014) The DNA-binding protein HU has a regulatory role in the acid stress response mechanism in Helicobacter pylori. Helicobacter 20:29–40
Toledo H, López-Solís R (2009) Tetracycline resistance in Chilean clinical isolates of Helicobacter pylori. J Antimicrob Chem 65:470–473
Valenzuela M, Perez-Perez G, Corvalán AH, Carrasco G, Urra H, Bravo D, Toledo H, Quest AFG (2010) Helicobacter pylori induced loss of the inhibitor-of-apoptosis protein survivin is linked to gastritis and death of human gastric cells. J Infect Dis 202:1021–1030
del Campo M, Toledo H, Lagos N (2013) Okadaic acid toxin at sublethal dose produced cell proliferation in gastric and colon epithelial cell lines. Mar Drugs 11:4751–4760
Stevens K, Sheldon B, Klapes A, Klaenhammer T (1991) Nisin treatment for inactivation of Salmonella species and other gram-negative bacteria. Appl Environ Microbiol 57:3613–3615
Romero C, Medina E, Vargas J, Brenes M, De Castro A (2007) In vitro activity of olive oil polyphenols against Helicobacter pylori. J Agric Food Chem 55:680–686
Rodríguez M, Alberto M, Manca de Nadra M (2007) Antibacterial effect of phenolic compounds from different wines. Food Control 18:93–101
Monagas M, Bartolomé B, Gómez-Cordovés C (2005) Updated knowledge about the presence of phenolic compounds in wine. Crit Rev Food Sci Nutr 45:85–118
Brown JC, Huang G, Haley-Zitlin V, Jiang X (2009) Antibacterial effects of grape extracts on Helicobacter pylori. Appl Environ Microbiol 75:848–852
Bisignano C, Filocamo A, La Camera E, Zummo S, Fera MT, Mandalari G (2013) Antibacterial activities of almond skins on cagA-positive and-negative clinical isolates of Helicobacter pylori. BMC Microbiol 13:103–109
Brown JC, Jiang X (2013) Activities of muscadine grape skin and polyphenolic constituents against Helicobacter pylori. J Appl Microbiol 114:982–991
Ruggiero P, Tombola F, Rossi G, Pancotto L, Lauretti L, Del Giudice G, Zoratti M (2006) Polyphenols reduce gastritis induced by Helicobacter pylori infection or VacA toxin administration in mice. Antimicrob Agents Chemother 50:2550–2552
Díaz-Gómez R, Toledo-Araya H, López-Solís R, Obreque-Slier E (2014) Combined effect of gallic acid and catechin against Escherichia coli. LWT Food Sci Technol 59:896–900
Spencer JPE (2003) Metabolism of tea flavonoids in the gastrointestinal tract. J Nutr 133(10):3255S–3261S
Obreque-Slier E, Peña-Neira A, López-Solís R, Zamora-Marín F, Ricardo da Silva J, Laureano JO (2010) Comparative study of the phenolic composition of seeds and skins from Carmenere and Cabernet Sauvignon grape varieties (Vitis vinifera L.) during ripening. J Agric Food Chem 58:3591–3599
Acknowledgments
This study was partially funded by Program U-INICIA VID 2011 (EO) and by grants University of Chile 11/05 (EO) and Fondecyt-Chile 1120126 (HT). We would also want to thank Mr. Nicanor Villaroel for his technical support.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to disclose.
Compliance with ethics requirements
The study was conducted according to the recommendations of the Declaration of Helsinki and approved by the Ethics Committee of the Faculty of Medicine, University of Chile.
Rights and permissions
About this article

Cite this article
Escandón, R.A., del Campo, M., López-Solis, R. et al. Antibacterial effect of kaempferol and (−)-epicatechin on Helicobacter pylori . Eur Food Res Technol 242, 1495–1502 (2016). https://doi.org/10.1007/s00217-016-2650-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00217-016-2650-z