
Abstract
Carbon, nitrogen and phosphorus inputs through atmospheric deposition, surface runoff and point sources were measured in the Ganga River along a gradient of increasing human pressure. Productivity variables (chlorophyll a, gross primary productivity, biogenic silica and autotrophic index) and heterotrophy (respiration, substrate induced respiration, biological oxygen demand and fluorescein diacetate hydrolysis) showed positive relationships with these inputs. Alkaline phosphatase (AP), however, showed an opposite trend. Because AP is negatively influenced by available P, and eutrophy generates a feedback on P fertilization, the study implies that the alkaline phosphatase can be used as a high quality criterion for assessing river health.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
There is substantial evidence that human activities have caused and will continue to lead profound effects, both directly and indirectly, on ecosystem functions. Surface waters, particularly, are more vulnerable to anthropogenic-changes occurring in the watershed. Rivers receive inputs from the watershed and, from within-stream erosion and transport. Understanding the changing links between these domains and associated shifts in microbial and biogeochemical processes in rivers is an important scientific challenge (Castillo et al. 2003; Gibbons et al. 2014). Unlike terrestrial ecosystems, the microbial processes in rivers suffers episodic events driven by urban-industrial flushing (more important during low flow) and rain driven pulsed connection between terrestrial and aquatic components. Nutrient driven increases in autochthonous-C together with enhanced input of allochthonous-C activate microbes and microbial processes (e.g., increased heterotrophy and microbial processing of organic matter). Fluxes of nutrients and organic matter in rivers like Ganga are driven principally by urban-industrial releases, atmospheric deposition (AD) and surface runoff (Pandey et al. 2014). The Ganga River with highest population density and extensive agricultural land (73.44% of total basin area) in its catchment (1,086,000 km2) becomes particularly sensitive to AD- inputs and surface based nutrient delivery. The Ganges basin receives ~3.32 Tg reactive- N (Nr) and ~173 Gg P annually through atmospheric deposition (Pandey et al. 2016). This additional supply together with surface runoff and sewage input can significantly change the autotrophic-heterotrophic linkages in the Ganga River.
During recent years a substantial number of water quality indices have been developed for effectively addressing to what degree the water has been rendered unsuitable for various usages and how much P reduction would be necessary for rejuvenating the system to a healthy ecological condition (Ponader et al. 2008). Total microbial activity and hydrolytic enzymes are often used as a predictor of organic matter turnover in soil, litter and water (Schnurer and Rosswall 1982; Costa et al. 2007). Alkaline phosphatase, an important hydrolytic enzyme, produced by bacteria and phytoplankton, plays a crucial role in P cycling and during P- starvation (Duhamel et al. 2010). About 78% of the AP activity is contributed by the aerobic bacteria attached to the algae and zooplanktons. On the contrary, a group of anaerobic bacteria decreases the AP activity by increasing the orthophosphate concentrations (Mhamdi et al. 2007). The AP activity can be used to relate P- status because AP is negatively influenced by P supply (Sayler et al. 1979). Therefore, quantifying AP would help better understand how human activities impact P loads, river eutrophy and feedbacks in surface waters. In the present study, conducted along a 37 km river stretch, we tested whether microbial activity in the Ganga River varies with changing influence of land use and urban core, and whether alkaline phosphatase activity can be used as a tool for monitoring the water quality. Because P concentration is rapidly rising in the Ganga River and because P is intricately linked with primary productivity and feedbacks at sediment–water interface, data on water quality interaction with alkaline phosphatase will provide important cues for assessing river health.
Materials and Methods
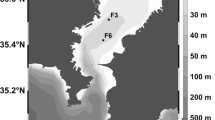
This 3 years study (March 2013–February 2016) was conducted at three sampling stations of Ganga River covering ~37 km representing up- and downstream the Varanasi urban core (25°18′N lat. 83°1′E long.) (Fig. 1). The climate of the area is tropical monsoonal. The year can be divided in a humid monsoon season (July–October), a cold winter (November–February) and a hot-dry summer (March–June). Over 80% of annual rainfall (~1050 mm) occurs during monsoon season. Soil of the region is highly fertile eutric cambisols and shallow luvisols.

Map of the study area showing sampling Sites 1 Adalpura (Adpr); 2 Assighat (Asht); 3 Rajghat (Rjht); a– f are sewage sampling sites where a and b are associated with Site 1, c with Site 2, and d, e and f with Site 3
Each study station represents three sites and samples for AD, river water and sediment were collected from each site in triplicates on monthly interval. Atmospheric deposition was collected fortnightly in triplicate using bulk samplers made up of 5 L high-density polyethylene buckets connected to a Teflon funnel. Thymol was used as biocide in collection bottles to avoid changes in nutrient concentration. Concentrations were converted to deposition considering time and the area of the bulk samplers. The values were then converted to per hectare and subsequently on sub-watershed scale for Adalpura (8400 ha), Assighat (13,400 ha) and Rajghat (17,500 ha) as described in Pandey et al. (2014). For surface runoff, sites were selected based primarily on land use and the strategy was to collect storm water at single outlet from the area marked for the purpose. Surface runoff water was collected in triplicate on event basis, initiated from the first flush, using pre-sterilized plastic bottles. Sampling details for surface runoff is described elsewhere (Pandey et al. 2014). Runoff volume was determined considering flow velocity and cross area of discharge outlet. Values were converted to sub-watershed scale by multiplying the discharge with mean concentrations. River water samples were collected from each site from directly below the surface (15–25 cm depth) in acid rinsed plastic containers. NO3 − was analysed following Voghel (1971) and NH4 + using Nessler’s reagent. Orthophosphate (dissolved reactive-P; DRP) was measured following ammonium molybdate-stannous chloride method (APHA 1998). The sum of NO3 − and NH4 + represents dissolved inorganic-N (DIN). Dissolved organic carbon (DOC) in water sample was measured following KMnO4 digestion method (Michel 1984). Sewage water was collected from the mouth of sewage outlets a–f (Fig. 1) and analyzed for NO3 −, NH4 +, PO4 3− and DOC as above. The Assi drain, marked as c in Fig. 1 represents the average sewage composition for sub-watershed 2 because it is the only outlet in study area 2 that carries sewage load from many small outlets originating from urban and sub-urban areas. Total nutrients and DOC entering through sewage were calculated by multiplying unit concentrations with annual discharge of the drain.
Biological oxygen demand (BOD) in river water was quantified following standard methods (APHA 1998). Chlorophyll a (Chl a) was measured following acetone extraction procedure (Maiti 2001). Gross primary productivity (GPP), photosynthetic activity and respiration all were measured by light and dark bottle method (APHA 1998). Ash free dry mass (AFDM) was determined by quantifying the weight loss at 525°C. Chlorophyll a and AFDM were used to calculate autotrophic index (AI). Sediment samples (25–30 m reach; 0–10 cm depth) were collected in triplicate using core samplers at monthly intervals. Substrate-induced respiration (SIR) was measured in 20 g of saturated sediment amended with 120 mg of glucose (Wardle et al. 1993). Alkaline phosphatase (AP) was determined following Tabatabai and Bremner (1969) and fluorescein diacetate hydrolytic activity (FDAase) by filtering and incubating the sediment suspension with fluorescein diacetate at 24°C (Schnurer and Rosswall 1982).
Three way analysis of variance (ANOVA) was used to test the effects of site, season and year. Regression analysis was used to test significant relationship between variables. Canonical correlation analysis (CCorA) was used to ordinate sampling locations and environmental variables using XLSTAT 2016. SPSS package (version 16) was used for statistical analysis.
Results and Discussion
In aquatic ecosystems, natural biogeochemical cycles are often disturbed by human-driven episodic flushing of carbon and nutrients. Such additional supply of, for instance N and P, enhances autochthonous C build-up and provides feedback to C dynamics and microbial processes (Castillo et al. 2003). In this study, a broad range of N and P concentrations were observed with values increasing downstream urban core (Fig. 2), and the differences due to site, season and year were highly significant (F values ranged from 10.50 to 170.30; p < 0.0001; ANOVA). NO3 − concentrations ranged from <154 µg L−1 at Adpr (upstream) to >296 µg L−1 at Rjht (downstream), and PO4 3− from <22 to >124 µg L−1. Thus, PO4 3− concentrations ranged from less than the benchmark of P limitation (30 µg L−1 as SRP) to sizably greater than that benchmark (Stevenson et al. 2012). This trend is similar to those observed in streams draining urban land use (Wahl et al. 1997). Each year, Rjht sub-watershed receives 770.70 tons DIN (NO3 − + NH4 +), 64.75 tons DRP (PO4 3−) and 558.60 tons DOC as AD input; 194.18 tons DIN, 48.55 tons DRP and 1456.35 tons DOC through sewage; and 225.75 tons DIN, 41.13 tons DRP and 2764.13 tons DOC through surface runoff. These values were three to seven fold higher than those received at upstream Adpr (Table 1).

Box-plot with upper and lower quartiles for nutrients, dissolved organic carbon (DOC) and productivity variables in the river from Adpr to Rjht with results of post hoc test. Lines represent the mean differences (MD) between sites at p < 0.05
Productivity variables (Chl a, GPP and photosynthetic rate) and heterotrophic functions (BOD, AI and respiration) all increased downstream (Figs. 2, 3) and showed significant effect of site, season and year (F = 17.09–1420; p < 0.0001; ANOVA). FDAase and SIR in sediment, increased overtime and downstream the urban land use with significant site-wise differences (F = 74.67–1160.00; p < 0.0001; ANOVA). The values were highest in summer and lowest in rainy season. AP did show an opposite trend with values being highest in rainy season (Fig. 4). Respiration and BOD showed positive correlation (R2 = 0.26; p < 0.004 and R2 = 0.38; p < 0.001) with FDAase but negative correlation (R2 = 0.85; p < 0.001 and R2 = 0.82; p < 0.001) with AP. Respiration showed significant correlation with DOC (R2 = 0.28; p < 0.004) and BOD (R2 = 0.82; p < 0.001) (Fig. 5). Further, despite similar trend downstream, FDAase and AI showed weak negative correlation (R2 = 0.23; p < 0.004) due possibly to strong influence of hydrologic flow on GPP. CCorA separated productivity, heterotrophy and biogenic silica (BSi) towards nutrient rich side while AP and DO away from these variables (Fig. 6) indicating possible role of phosphorus and C pool influencing heterotrophy and AP (Castillo et al. 2003; Lau et al. 2009).

Trends in a biological oxygen demand (BOD), b photosynthetic activity, c respiration, and d AI in the river along the study gradient. Values are mean (n = 12) ± 1SD. S summer, R rainy, W winter

Trend in a SIR, b BSi, c FDAase, and d AP activity in river sediment. Values are mean (n = 12) ± 1SD. S summer, R rainy, W winter

Significant relationships between a phosphate and AP, b respiration and alkaline phosphatase, c BOD and alkaline phosphatase, d respiration and DOC, e AI and FDAase, f respiration and FDAase, g BOD and FDAase, and h BOD and respiration

CCorA biplot showing correlation between environmental variables and their canonical axes. Values above 0.5 are significant at p < 0.05; F 1 first canonical axes, F 2 second canonical axes
Sediment microbial communities are responsible for a number of metabolic activities in river ecosystems (Gibbons et al. 2014). Positive relationships among nutrients, organic-C and BOD together with FDAase and SIR acknowledge such links. The ecosystem level metabolism at nutrient-rich downstream sites was dominated by heterotrophs where a high proportion of primary production is expected to be channelled to DOC. The relative importance of heterotrophic bacteria in C-degradation and releasing AP is expected to change as the system shifts from low to high productivity. Low productivity at Adpr, for instance, was associated with substantially low concentrations of N and P. The P starved heterotrophic populations at Adpr presumably carry metabolic attributes that allow them to release AP. These considerations are relevant in human impacted rivers where increased supply of carbon and nutrients enhance heterotrophy. Furthermore, heterotrophic population may enhance dissolution of siliceous cell wall of diatoms shifting the partitioning and sequestration of biogenic silica and carbon in the Ganga River (Bidle and Azam 1999).
Most of the biomonitoring approaches are based either on time consuming extensive field sampling, or on cost intensive analytical techniques. Furthermore, simultaneous occurrence of multiple factors including those of hydrologic and climatic origin, effect of upstream water quality and anthropogenic influences often limit the use of specific bioassessment tools for monitoring river health (Lafont 2001; Schinegger et al. 2012; Turley et al. 2016; Pearson et al. 2016). AP is one of the most important ectoenzymes in river bed sediments (Sinsabaugh et al. 2009). In the present study, the AP activity showed a declining trend downstream due possibly to the anthropogenically added phosphorus. Negative correlation (R2 = 0.82; p < 0.001) between phosphate concentration and phosphatase activity in river sediment could be expected because high phosphate concentration inhibits phosphatase activity as a feedback (Newman and Reddy 1993). Typically, highest concentration of P coincided with lowest activity of AP indicating a P-induced feedback inhibition (Dyhrman et al. 2006). This effect was more pronounced at Rjht draining urban landuse. Furthermore, the cumulative effect of diffused and point sources of nutrients trigger C build-up and, consequently, enhance oxygen demand by heterotrophs creating low oxygen tension and sediment P release (Mhamdi et al. 2007). Overall, our analyses reveal consistent effects of P loading on alkaline phosphatase activity along the study stretch. The degree of consistency along the study gradient adds confidence to our interpretation that the AP activity in the riverbed sediment provides valuable information on water quality condition of the Ganga River. It may help exploring the likely cause and the magnitude of ecological degradation and deviation from pristine condition.
Our study establishes, in the Ganga River, phosphate-alkaline phosphatase linkages as a high quality criterion for assessing the river health and provides cues useful in river rejuvenation. Because AP activity is related implicitly with P availability and demand, and the latter links eutrophy with sediment P release as a feedback, positive correlation between FDAase (a measure of overall microbial activity) and BOD and negative correlation between AP and BOD if coupled with independently measured physical and chemical parameters with linear or non-linear dependence might be useful for assessing ecosystem resilience of rivers. Field measurements of these linkages provide important cues on which river rejuvenation strategies can be keyed.
References
APHA (1998) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, DC
Bidle KD, Azam F (1999) Accelerated dissolution of diatom silica by marine bacterial assemblages. Nature 397:508–512
Castillo MM, Kling GW, Allan JD (2003) Bottom-up controls on bacterial production in tropical lowland rivers. Limnol Oceanogr 48:1466–1475
Costa AL, Paixão SM, Caçador I, Carolino M (2007) CLPP and EEA profiles of microbial communities in salt marsh sediments. J Soil Sediments 7:418–425
Duhamel S, Dyhrman ST, Karl DM (2010) Alkaline phosphatase activity and regulation in the North Pacific subtropical gyre. Limnol Oceanogr 55(3):1414–1425
Dyhrman ST, Chappell PD, Haley ST, Moffett JW, Orchard ED, Waterbury JB, Webb EA (2006) Phosphonate utilization by the globally important marine diazotroph Trichodesmium. Nature 439:68–71
Gibbons SM, Jones E, Bearquiver A, Blackwolf F et al (2014) Human and environmental impacts on river sediment microbial communities. PLoS ONE 9(5):e97435. doi:10.13771/0097435
Lafont M (2001) A conceptual approach to the biomonitoring of freshwater: the ecological ambience system. J Limnol 60(Suppl. 1):17–24
Lau DCP, Leung KMY, Dudgeon D (2009) What does stable isotope analysis reveal about trophic relationships and the relative importance of allochthonous and autochthonous resources in tropical streams? a synthetic study from Hong Kong. Freshw Biol 54:127–141
Maiti SK (2001) Handbook of methods in environmental studies. Water and wastewater, vol 1. ABD, Jaipur
Mhamdi BA, Azzouzi A, Elloumi J, Ayadi H, Mhamdi MA, Aleya L (2007) Exchange potentials of phosphorus between sediments and water coupled to alkaline phosphatase activity and environmental factors in an oligo-mesotrophic reservoir. Compte Reus Biol 330: 419–428
Michel P (1984) Ecological methods for field and laboratory investigation. Tata McGraw-Hill Publishing Company, New Delhi
Newman S, Reddy KR (1993) Alkaline phosphatase activity in the sediment-water column of a hypereutrophic lake. J Environ Qual 22:832–838
Pandey J, Pandey U, Singh AV (2014) Impact of changing atmospheric deposition chemistry on carbon and nutrient loading to Ganga River: Integrating land–atmosphere–water components to uncover cross-domain carbon linkages. Biogeochemistry 14:179–198
Pandey J, Pandey U, Singh AV, Tripathi S, Mishra V (2016) Atmospheric N and P deposition in the Ganges basin. Curr Sci 110:974–976
Pearson CE, Ormerod SJ, Symondson WO, Vaughan IP (2016) Resolving large-scale pressures on species and ecosystems: propensity modelling identifies agricultural effects on streams. J Appl Ecol 53(2):408–417
Ponader KC, Charles DF, Belton TJ, Winter DM (2008) Total phosphorus inference models and indices for coastal plain streams based on benthic diatom assemblage from artificial substrates. Hydrobiologia 610:139–152
Sayler GS, Puziss M, Silver M (1979) Alkaline phosphatase assay for freshwater sediments: application to perturbed sediment systems. Appl Environ Microbiol 38(5):922–927
Schinegger R, Trautwein C, Melcher A, Schmutz S (2012) Multiple human pressures and their spatial patterns in European running waters. Water Environ J 26(2):261–273
Schnurer J, Rosswall T (1982) Fluorescein diacetate hydrolysis as a measure of total microbial activity in the soil and litter. Appl Environ Microbiol 43:1256–1261
Sinsabaugh RL, Hill BH, Shah JJF (2009) Ecoenzymatic stoichiometry of microbial organic nutrient acquisition in soil and sediment. Nature 462:795–798
Stevenson RJ, Bennett BJ, Jordan DN, French RD (2012) Phosphorus regulates stream injury by filamentous green algae, DO, and pH with thresholds in responses. Hydrobiologia 695:25–42
Tabatabai MA, Bremner JM (1969) Use of p- nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol Biochem 1:301–307
Turley MD, Bilotta GS, Chadd RP, Extence CA, Brazier RE, Burnside NG, Pickwell AGG (2016) A sediment-specific family-level biomonitoring tool to identify the impacts of fine sediment in temperate rivers and streams. Ecol Indic 70:151–165
Voghel AI (1971) A text book of quantitative inorganic analysis, 4th edn. The English Language Book Society, Longman, Essex
Wahl MH, Mckellar HM, Williams TM (1997) Patterns of nutrient loading in forested and urbanized coastal streams. J Exp Mar Biol Ecol 213:111–131
Wardle DA, Yeates GW, Watson RN, Nicholson KS (1993) Response of soil microbial biomass and plant litter decomposition in maize and Asparagus cropping systems. Soil Biol Biochem 25:857–868
Acknowledgements
The authors are thankful to the Coordinator, Centre of Advanced Study in Botany, Banaras Hindu University for facilities and to Banaras Hindu University-University Grants Commission, New Delhi for funding support as fellowship to A.Y.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Yadav, A., Pandey, J. Water Quality Interaction with Alkaline Phosphatase in the Ganga River: Implications for River Health. Bull Environ Contam Toxicol 99, 75–82 (2017). https://doi.org/10.1007/s00128-017-2108-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00128-017-2108-4