
Abstract
Genome instability manifested by multiple reorganization contributes to the acquisition of new properties by tumor cells, resistance to therapy in particular. Structural rearrangements of chromosomes can be induced by damage to DNA, for example, by ionizing radiation. In this study, changes in the karyotype of the pseudodiploid cell line HindIIIG-1 obtained after irradiation of apoptosis-resistant transformed rat cells HindIIIG have been examined. The HindIIIG-1 line is a result of polyploidization associated with the cell cycle block in the G2/M phase and subsequent depolyploidization. The original HindIIIG line, before irradiation, was represented mainly by a pseudodiploid population with the normal number of 42 chromosomes, tetraploid fraction composed of 14%. The cell karyotype had two numerical and one specific structural rearrangement of chromosomes (SRCs), der(14). Irradiation induced polyploidization and multiple fragmentations of chromosomes. In the process of cultivation, a pseudodiploid population of cells with the karyotype similar to the karyotype of nonirradiated cells began to predominate, but the number of clonal and nonclonal SRCs increased. At late passages, the karyotype of HindIIIG-1 cells again became identical to the karyotype of cells before irradiation and did not contain new clonal SRCs. The role of nonclonal and clonal SRCs in the process of survival of irradiated cells and the formation of new cell populations is discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Chromosomal instability is a key characteristic of tumor cells. The population of tumor cells is heterogeneous in terms of clonal and nonclonal numerical and structural rearrangements of chromosomes (SRCs). Nonclonal SRC facilitates cell adaptation to a changing environment, while clonal SRCs have been seen to alter the expression of many genes, including oncogenes and tumor suppressors (Mitelman et al, 2007; Ohgak and Kleihues, 2009).
Chromosomal instability is induced by DNA-damaging agents, such as ionizing radiation, causing single- and DNA double-strand breaks (DSBs ). Disruption of DNA replication or going through mitosis after the replication fork collapse can also lead to a DS-DNA that promotes SRC (Jegg and Löbrich, 2006; Kakarougkas et al., 2014).
Chromosome fragmentation often observed in polyploid tumor cells after irradiation or anticancer therapy alters the expression profile of many genes (Crasta et al., 2012; Zhang et al., 2015; Ly and Cleveland, 2017; Pellestor, 2019; Ye et al., 2019). Mutations of the tumor suppressor p53 (TP53) result in changes or loss of its functions associated with changes in cell viability (Mardin et al., 2015; Zhang et al., 2015).
Earlier, we showed that apoptosis-resistant rat embryonic fibroblasts transformed with the HindIIIG region of human adenovirus 5 overcame the process of cell death through specific mechanisms (Chitikova et al., 2014). Irradiation of HindIIIG cells generated giant polyploid cells due to DNA replication while maintaining the G2/M block of the cell cycle The survival of the irradiated population was ultimately provided by HindIIIG-1 cells with DNA content close to that of untreated HindIIIG cells (Chitikova et al., 2014).
In this work, we analyzed the dynamics of chromosomal rearrangements in HindIIIG-1 cells arose after irradiation and during long-term cultivation resulted in the return of pseudodiploid proliferating cells, which ensured provided the viability of the cell population.
MATERIALS AND METHODS
Cells
The HindIIIG cell line was established from rat embryonic fibroblasts by cotransfection of the HindIIIG early region of human adenovirus type 5 (E1A + E1B 19 kDa) with the pSV 2-neo plasmid carrying the geneticin-resistance gene. HindIIIG cells were cultured in Dalbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum and 0.1 mg/mL gentamicin at 37°C in the presence of 5% CO2.
Irradiation
HindIIIG cells were exposed to X rays at 6 Gy with an Axiom Iconos R200 radiology system (Siemens, Germany). Within 27 days after irradiation, HindIIIG cells were cultured until a monolayer appeared without subculturing. HindIIIG cells that survived after X-ray irradiation were named HindIIIG-1. HindIIIG-2 cells were obtained after subsequent irradiation of HindIIIG-1 cells at the same dose.
Karyotypic Analysis
To obtain metaphase plates, the culture medium was supplemented with colchicine at a final concentration of 0.06 μg/mL for 1–1.5 h. Hypotonic treatment was carried out with 0.55% KCl solution (ICN Biomedical Inc., United States) at 37°C for 12–15 min. The cells were then fixed with a mixture of methanol and glacial acetic acid (3 : 1) for 15 min, with it being changed twice. The suspension of fixed cells was dropped on glass slides over a water bath at 52–55°C until complete drying took place. To identify chromosomes, preparations were stained using the G-banding method (GTG-method) (Ozkinay et al., 1979). The karyotype of the new line HindIIIG-1 obtained from cells that survived after irradiation was examined at passages 8, 17, 22, and 34.
To identify regions of metaphase chromosomes carrying ribosomal genes, preparations were stained with 50% silver nitrate (AgNO3, Sigma, United States) (Howell and Black, 1980). They were then treated with 2% gelatin solution for 8–10 min at 56°C, washed in water, dehydrated with ethyl alcohol, and stained using the standard G-banding technique. Chromosomes were identified according to the chromosome ideograms of rat Rattus norvegicus for 300 bands (Satoh et al., 1989). The karyotype was described in accordance with the International System for Human Cytogenetic Nomenclature [(Shaffer et al., 2009).
RESULTS
Karyotype of Transformed HindIIIG Cells
Analysis of the karyotype of HindIIIG cells before irradiation showed that the cell population was mostly pseudodiploid and heterogeneous. The proportion of mitotic polyploid cells in the HindIIIG population before irradiation was 14 and 2% had a ploidy of more than 4n.
The number of chromosome (NC) varied from 40 to 44, with the modal class with NC 42 being observed only in 38% of cells. In all HindIIIG cells, only one SRC, probably a duplication of the q21–q22.1 region of the long arm of chromosome 14 with a deletion of the q22–qter telomere region, was detected. Two cells had karyotype 42,XX,der(14)dup(14)(q21q22.1) del(14)(q22qter),–15,+20 with monosomy of chromosome 15, trisomy of chromosome 20, and rearrangement of chromosome 14 (Fig. 1). In other cells, in addition to these permanent clonal rearrangements of chromosomes, other clonal and nonclonal SRCs or altered NCs were registered.

Karyotype of HindIIIG cells with modal number of chromosome and without additional numerical and structural chromosome rearrangements before irradiation. G-banding; arrow indicates a permanent structural rearrangement of chromosome 14: 42,ХХ,der(14)?dup(14)(q21q22.1)del(14)(q22qter),−15,+20.
In cells with NC 43, two homologues of chromosome 15 were found. In 10% of cells with NC 41, chromosome 3 had an enlarged p12 locus of the short arm and amplification of ribosomal genes (ARG) was observed (Figs. 2a–2c). In addition, these cells had a rearranged chromosome 16 der(16)t(8;16)(q13;p12) and lacked the third normal homologue of chromosome 20.

Amplification of ribosomal genes in the short arm of chromosome 3 in HindIIIG cells before and after irradiation. (a) Karyotype of HindIIIG cells before irradiation with ARG at the p12 locus of chromosome 3, as well as other clonal and nonclonal numerical and structural rearrangements of chromosomes 41,ХХ,der(1)t(1;8)(q43;11.2)der(3)amp(3)(p12)t(3;?20) (p13;?q11),add(4)(q42),+der(8)t(8;17)(q11.2;p12),der(14)?dup(14)(q21q22.1)del(14)(q22qter),−15,der(16)t(8;16)(q13;p12)+6–17. Arrows show rearranged chromosomes. (b) Structural changes in chromosome 3 from three different HindIIIG-1 cells at the 17th passage after irradiation; the left homologue is the normal homologue of chromosome 3, the right homologue has ARG at the p12 locus. (c) ARG at the p12 locus of chromosome 3; arrows point to chromosomes 11, 12, and 3 with functionally active ARG. (a, b) G-banding. (c) Staining with AgNO3.
The karyotype of HindIIIG cells was analyzed after irradiation on days 1, 4, and 27 (before the first passage of the stable line). One day after irradiation, mitotic cells with a ploidy higher than 4n were not detected (Table 1), the proportion of tetraploid cells was 17.7%. On the fourth day after irradiation, almost all cells became polyploid with a ploidy higher than 4n.
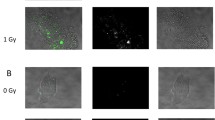
On the fourth day after irradiation, nuclear fragmentation of giant polyploid cells and accumulation of micronuclei around them (Fig. 3), as well as metaphases with chromosomes having one- and two-chromatid breaks (Fig. 4), were observed. In addition, on the fourth day after irradiation, large ploidy metaphases plats were found in 9.3%, which simultaneously contained multiple chromosomal fragments with whole metaphase chromosomes. (Fig. 5). At later stages of cultivation, the number of such metaphase plates decreased. It should be noted that the number of metaphases with chromosome fragmentation declined, which coincided in time with a decrease in the number of metaphases with a ploidy more than 4n. At later stages after irradiation, tetraploid and pseudodiploid metaphase plates with fragmentation of one or several chromosomes were observed (Fig. 6).

(a, b) Numerous micronuclei produced by fragmentation of the giant polyploid nucleus in HindIIIG cells 4 days after irradiation

Metaphase plate of HindIIIG cells with multiple (a) one- and (b) two-chromatid breaks (arrows) and incomplete condensation of chromatin 4 days after irradiation.

Polyploid metaphases with chromosome fragmentation and intact chromosomes in HindIIIG cells at the fourth passage after irradiation. (a) Strong fragmentation. (b) Less expressed fragmentation with an adjacent micronucleus.

Metaphase plate and karyotype of HindIIIG-1 cells with fragmentation in several chromosomes. Passage 22; mar, marker chromosome; arrows point to rearranged chromosomes and chromosome fragmentation.
Analysis of the karyotype of HindIIIG cells on day 27 after irradiation revealed only 2% of tetraploid mitotic cells. However, the heterogeneity of the cell population increased in comparison with the HindIIIG cells before irradiation. The number of cells with 41 chromosomes, as well as the number of nonclonal SRCs (Tables 1, 2), was found in 12 out of 13 cells (92%). The number of chromosomes involved in nonclonal SRC in the karyotype more than doubled compared with nonirradiated cells. The changes affect different chromosomes, but more often chromosomes 6 and 17 (Fig. 7). Rearranged chromosome 14 was detected in all HindIIIG cells before and after irradiation. Monosomy of chromosome 15 was observed in 9 out of 13 cells as before irradiation. Chromosome 3 with amplification at the p12 locus was not found (Table 2).

Breakpoints of structurally rearranged chromosomes in HindIIIG cells before and after irradiation in the new line HindIIIG-1 at different times after irradiation and the new line HindGIII-2. Colors show the points of chromosome breaks. Red, in nonirradiated HindIIIG cells; blue, 27 days after irradiation of HindIIIG cells; green, lilac, and brown, after irradiation of HindIIIG-1 cells at passages 17, 22, and 34, respectively; dark lilac, cells after repeated irradiation of HindIIIG-1 (HindIIIG-2 cells), passage 12. Dots: circles, chromosome breaks in clonal SRCs; colored squares, breaks in clonal SRCs, which are less than 10%; colored triangles, chromosome breaks in nonclonal SRCs; white squares, breakage points in SRCs that became nonclonal at later stages of cultivation; white triangles, breaks in which the presence of structural rearrangements is questionable; square brackets, areas of chromosome breaks that cannot be accurately identified.
HindIIIG-1 Cell Karyotype
The karyotype of the HindIIIG-1 cell line obtained from cells that survived after irradiation was studied at passages 8, 17, 22, and 34. The number of tetraploid cells during subculturing decreased compared to that of nonirradiated HindIIIG cells, and, at late passages, less than 1% were cells with a ploidy higher than 4n (Table 1). Almost the entire population of HindIIIG-1 cells became pseudodiploid.
Analysis of HindIIIG-1 cells at the 17th passage showed a further increase in the heterogeneity of the HindIIIG-1 cell population compared to the composition of HindIIIG population 27 days after irradiation. The modal class of cells in terms of the number of chromosomes was not evident and two types of cells with NC 41 and 42 prevailed (Table 1). In 18 of 20 cells (90%), clonal, non-clonal, numerical, and structural chromosome rearrangements were found. Monosomy of chromosome 15 was revealed in all cells. The number of novel nonclonal SRC more than doubled in comparison with the karyotype of HindIIIG cells on the 27th day after irradiation (Table 2). The cell number with ARG in the short arm of chromosome 3 was three times greater than in HindIIIG cells before irradiation. In addition, in the HindIIIG-1 karyotype, about 17% have a new SRC, a derivative of chromosome 5-der(5)t(1;5;8)(q22?q34;q36;q22), which involved chromosomes 1 and 8. In these cells, apart from der(5), other chromosomes with altered structure were revealed (Fig. 8a).

Karyotype of new pseudodiploid cell lines HindIIIG-1 and HindIIIG-2. (a) HindIIIG-1 karyotype at passage 17 after irradiation with novel numerical and structural rearrangements of chromosomes: 41,Х,−Х,−4,der(5)t(1;5;8)(q22;?q34q36; q22),der(7)t(X;7)(q12;q32),+8,der(8)(q22)add(q22;?),der(14) ?dup(14)(q21q22.1)del(14)(q22qter),−15,+20,+mar(?;7)(?cen; q32); mar, marker chromosome; (b) karyotype of HindIIIG-2 cells (after repeated irradiation of HindIIIG-1 cells) at the 12th passage: 41,Х,−Х,−4,der(5)t(1;5;8)(q22?;q34q36; q22),der(7) t(X;7)(q12;q32),+8,der(8)(q22)add(q22;?),der(14)?dup(14) (q21q22.1)del(14)(q22qter),−15,+20,+mar(?;7)(?cen;q32). Numbers by der(5) indicate loci of chromosome breaks, arrows show rearranged chromosomes.
At the 22nd passage of cultivation, HindIIIG-1 cells had the most stable karyotype with clearly pronounced modal class, represented by cells with the number of chromosomes equal to 42 and composed 66% of the population Karyotype 42,XX,der(14) dup(14)(q21q22.1)del(14)(q22qter),−15,+20 without non-clonal SRC was observed in 13 cells from 17 (76.5% of cells), and only in 4 cells, except for der(14), new non-clonal SRC were identified. Clonal rearrangements of chromosomes 3 and 5 were detected only in 1% of cells (Table 2). Karyotypic analysis of HindIIIG-1 cells at passage 34 revealed a slight increase in karyotype instability. Proportion of tetraploid cells, as well number of cells with non-clonal SRC increased and no new clonal SRC were found. Nevertheless, the modal class of cells with NC 42 was clearly expressed (Tables 1 and 2).
Rearrangement of chromosome 14 in HindIIIG-1 cells was detected at all periods of observation after irradiation (as in the original HindIIIG cells). It is a permanent specific structural rearrangement in these lines. Monosomy of chromosome 15 was found in all cells after irradiation, in contrast to nonirradiated HindIIIG cells. Trisomy of chromosome 20, was also typical for HindIIIG-1 cells.
All chromosomes in the karyotype underwent structural clonal or non-clonal changes. More often than others, were chromosomes X, 1, 2, 3, 4, 6, 8, 10 (6 or more breaks per chromosome). Chromosome breaks more often occurred at loci 1q32, 1q34, 4q42, 5q36, 11q22, and 20p12 (5 and more breaks per locus) (Fig. 7).
Thus, analysis of the karyotype of the pseudodiploid cell line HindIIIG-1 (obtained by long-term cultivation of irradiated HindIIIG cells) showed that cells at late passages had a the distinct modal karyotype NC 42, and the proportion of polyploid cells decreased by more than twice compared to with nonirradiated HindIIIG cells. Despite the multiple SRC observed at the 17th passage after irradiation, new clonal numerical and SRC in the karyotype of HindIIIG-1 cells were not identified at next passages. The number of non-clonal SRCs on the 22 passage after irradiation became equal to their number before irradiation in the Hind IIIG line.
At passages 22 and 34, cells with normally condensed metaphase chromosomes as well with prophase under-condensed chromosomes were observed (Fig. 9). This probably occurs in binucleated cells, whose nuclei do not enter mitosis synchronously, and premature chromatin condensation happens in the retarded nucleus.

Tetraploid metaphase plate in HindIIIG-1 cells, having metaphase and simultaneously prophase undercondensed chromosomes. Passage 34.
Repeated irradiation of HindIIIG-1 cells (HindIIIG-2 cells). To understand the regularities of the diploidy arrangement in apoptosis-resistant HindIIIG cells and specificity of karyotype changes after irradiation, HindIIIG-1 cells were irradiated at the 8th passage. HindIIIG-1 cells survived after repeated irradiation was named HindIIIG-2. These cells also contain metaphases (0.6%) with increased ploidy and the presence of fragmented chromosomes along with intact metaphase chromosomes.
Analysis of the karyotype of HindIIIG-1 cells at the 8th passage before irradiation reveal a clear modal class with NC 41 (52% of cells), while cells with NC 42 accounted for only 25%. The karyotype of HindIIIG-2 cells was analyzed at the 12th passage after irradiation. Modal class with NC 42 and two subpopulations of cells with NC 41 and 43 were identified (Table 1). Karyotype 42,XX,der(14)dup(14)(q21q22.1)del (14)(q22qter),–15,+20 without new SRC in HindIIIG-2 cells was found only in 4 cells. In HindIIIG-1 cells at the 8th passage it was registered in 2 cells. Other HindIIIG-2 cells, along with der(14), had new clonal and non-clonal SRC. Two clones, accounting for more than 10% of the population, had the same SRC as HindIIIG-1 cells at passage 17. The first clone had ARG at the p12 locus of chromosome 3 and t(8;16)(8q13;16p12) translocation (Fig. 2a). The karyotype of the second clone contained der(5)t(1;5;8)(q22q?34;q36;q22), der(7), and der(8) (Fig. 8b). The number of cells with these rearrangements differed insignificantly from HindIIIG-1 cells irradiated once at the 17th passage (Table 2). The total number of non-clonal SRCs and chromosome breaks in HindIIIG-2 cells was lower than in HindIIIG-1 cells at the 17th passage after the first irradiation.
Thus, it has been shown that X-ray irradiation of HindIIIG cells at a dose of 6 Gy induced population polyploidization and reorganization of the entire genome: fragmentation of giant polyploid nuclei, formation of many micronuclei, and fragmentation of chromosomes in polyploid cells. The appearance of a new population of pseudodiploid cells was accompanied by the appearance of a large number of non-clonal SRCs and the reappearance of SRCs der(5) and der(7), as well as an increase in the number of cells with SRC amp(3) and der(8).
DISCUSSION
We have previously shown that the cell cycle checkpoint at the G2/M boundary to be disrupted in HindIIIG cells. This resulted in sustained DNA replication; in the absence of cytokinesis, the amount of DNA in cells reaches 110 or more (Chitikova et al., 2014). Four days after irradiation of HindIIIG cells, metaphase plates of very high ploidy with multiple chromosomal fragments, as well as intact metaphase chromosomes, were observed. Fragmentation of chromosomes leads to a genomic chaos in cells and karyotypic heterogeneity. From all cell variants, cells with an updated genome and the most adapted to new conditions are selected (Liu et al., 2014; Ye et al., 2019). In our study, HindIIIG cells did not die after irradiation at a dose of 6 Gy and, moreover, became pseudodiploid and proliferating cells. In some cells, the expression of Nanog and Oct 3/4 stem cell markers was found (Chitikova et al., 2014).
The karyotype of HindIIIG-1 cells formed after irradiation at all passages retained clonal SRCs typical for the original HindIIIG cells: rearranged chromosome 14—der(14)dup(14)(q21q22.1)del(14)(q22qter), presumably, with partial duplication of the q21-q22.1 region and monosomy of chromosome 15, as well as trisomy of chromosome 20. As a result of monosomy of chromosome 15, these cells contain only one copy of the Rb1 gene, which is localized at the q12 locus of its long arm. The RB1 protein controls the G1/S phase transition preventing cells from entering the cycle with abnormal chromosomes. As a result of trisomy of chromosome 20, which contains the genes Itgb2, Notch4, Cdk1, Cdkn1b, Srpk1, Vegfa, Rab11b, Tnf, Hsp70-1, Ras3, and Rps18 (according to the rat genome database (RGD), http://rgd/ mcv/edu); in the genome of HindIIIG and HindIIIG-1 cells, these genes are in three copies.
On the 17th passage of the HindIIIG-1 line, a clone of cells with a rearranged chromosome 5, der(5)t(1;5;8)(q22?q34;q36;q22) consisting of fragments of three chromosomes was detected (according to the resolution of the GTG method)
Chromosome 5 has been broken at the q36 locus, which is often involved in rearrangements in HindIIIG-1 cells and is not accidental. In this locus, according to the RGD rat genome database, genes associated with the development of cancer and the maintenance of the tumor phenotype have been identified: Casp9, Kif1b, Pax7, Pik3r3, Prkaα2, Rragc, Runx3, and Wnt4.
In addition, in the HindIIIG-1 line at passage 17, the proportion of cells with ARG at the p12 locus of chromosome 3 increased to 30%, as in HindIIIG-2 cells (32%), compared to HindIIIG cells before irradiation. It was reported (Chitikova et al., 2014) that these cells resumed proliferation, which is likely associated with an increased number of cells with ARG. The copy number and activity of ribosomal genes are important for the processes of proliferation, differentiation, aging, and apoptosis, as well as the maintenance of cell homeostasis (Grummt, 2013; Orsolic et al., 2015). During tumor transformation of cell and tumor progression, there is a change in both the copy number of ribosomal genes, and their movement within same chromosome or to other chromosomes (Mamaeva, 1996, 2000).
The appearance of cells with der(5) and the increased number of cells with ARG on chromosome 3 in the HindIIIG-1 and Hind IIIG-2 cell populations at the 17th and 12th passages, respectively, probably contribute to the survival of the entire population of these cells after irradiation.
At the 17th passage of HindIIIG-1 cells, in almost all cells (25 of 30 cells), the karyotype contained a large number of clonal and nonclonal SRCa involved different chromosomes (up to six rearrangements in one cell). Chromosomes 1, 2, 4, 5, 8, and 11 underwent changes more often than other chromosomes. All in all, 75 chromosome breaks were found, with the 11q22 locus of chromosome 11 being the most often subjected to breaks. Such number of cells with SRC and their preservation for a long time after irradiation can be a result of a delayed repair of -DNA DSB (Chitikova et al., 2014). The absence or impairment of -DNA repair leads to the appearance of chromosome rearrangements.
We observed rearranged chromosomes in metaphase plates from the first to fourth days after irradiation. Such cells should age or undergo apoptosis, but HindIIIG cells with -DNA DSB survive as a result of inactivation of the apoptosis program blocked by a high level of expression of the antiapoptotic gene E1B19kD (Han et al., 1995).
At the 22nd passage, HindIIIG-1 cells did not acquire a new clonal SRC and did not retain the nonclonal SRC detected at the 17th passage. The absence of HindIIIG-1 cells with a nonclonal SRC encountered at the 17th passage at later passages is possibly due to the inability of abnormal chromosomes to properly segregate into daughter cells as a result of an inactive or defective centromere, which can then undergo fragmentation and autophagy. At passage 22, we observed metaphase plates with small chromosomal fragments, probably from one or several rearranged chromosomes (Fig. 6). Cells can acquire a second chromosome homologue to replace the lost one due to the uniparental disomy mechanism, resulting in two chromosome homologues from the same ancestor (Tuna et al., 2009). It is possible that these chromosomes have genetic alterations that determine the new properties of HindIIIG-1 cells.
Currently, much attention is paid to nonclonal SRCs, which create the evolutionary potential of a population, and the number of which mirrors the genome instability. Nonclonal SRC increase the heterogeneity and plasticity of the cell population. This allows cells to adapt to altered living conditions and survive under genotoxic stress (Liu et al., 2014; Heng et al., 2016, 2018; Ye et al., 2018; Frias et al., 2019).
For example, transformed rat LRec-1 cells transferred to a serum-free medium acquired four novel clonal SRCs in their karyotype, which helped the cells to adapt to the changed environmental conditions (Yartseva et al., 2007). At the same time, the cell populations seem to have a stable karyotypic structure and a certain probability of returning to the initial stable state, as was shown for the populations of clones and subclones of the Chinese hamster CHO-K1 cell line (Polyanskaya et al., 1981). An increase in non-clonal and clonal SRCs in the population of HindIIIG-1 and HindIIIG-2 cells is necessary only for the survival of cells during the stress induced by irradiation. At later passages after irradiation, the karyotypic heterogeneity decreases. HindIIIG cells have a necessary, sufficient, and, at the same time, plastic genome, which allows cells to survive after the damage produced by radiation and create a new population of pseudodiploid cells resembling stem cells (Chitikova, 2014).
The emergence of a new population of rapidly proliferating cells is determined not by new clonal SRCs, but, apparently, by epigenomic changes responsible for the altered cell phenotype after irradiation.
REFERENCES
An International System for Human Cytogenetic Nomenclature, Shaffer, Slovak, M.L. and Cambell, L.G., Eds., Basel: S. Karger, 2009.
Chitikova, Z.V., Gordeev, S.A., Bykova, T.V., Zubova, S.G., Pospelov, V.A., and Pospelova, T.V., Sustained activation of DNA damage response in irradiated apoptosis-resistant cells induces reversible senescence associated with mTOR downregulation and expression of stem cell markers, Cell Cycle, 2014, vol. 13, p. 1424. https://doi.org/10.4161/cc.28402
Crasta, K., Ganem, N.J., Dagher, R., Lantermann, A.B., Ivanova, E.V., Pan, Y., Nezi, L., Protopopov, A., Chowdhury, D., and Pellma, B., DNA breaks and chromosome pulverization from errors in mitosis, Nature, 2012, vol. 482, p. 53. https://doi.org/10.1038/nature10802
Frias, S., Ramos, S., Salas, C., Molina, B., and Rivera-Luna, S.S.R., Nonclonal chromosome aberrations and genome chaos in somatic and germ cells from patients and survivors of Hodgkin lymphoma, Genes, 2019, vol. 10, p. 1. https://doi.org/10.3390/genes10010037
Grummt, I., The Nucleolus—guardian of cellular homeostasis and genome integrity, Chromosoma, 2013, vol. 122, p. 487.
Han, J., Sabbatini, P., Perez, D., Rao, L., Modha, D., and White, E., The E1B 19K protein blocks apoptosis by interacting with and inhibiting the P53-inducible and death-promoting Bax protein, Genes Dev., 1996, vol. 10, p. 461. https://doi.org/10.1101/gad.10.4.461
Heng, H.H.Q., Regan, S, M., Liu, G., and Ye, C.J., Why it is crucial to analyze non-clonal chromosome aberrations or NCCAs?, Mol. Cytogenet., 2016, vol. 9, p. 15.
Heng, H.H., Horne, S.D., Chaudhry, S., Regan, S.M., Liu, G., Abdallah, B.Y., and Ye, C.J., A postgenomic perspective on molecular cytogenetics, Curr. Genomics, 2018, vol. 19, p. 227.
Howell, W. and Black, D.A., Controller silver staining of nucleolus organizer regions with protective colloidal developer: a one-step method, Experientia, 1980, vol. 36, p. 1014.
Jeggo, P.A. and Löbrich, M., Radiation-induced DNA damage responses, Radiat. Prot. Dosimetry, 2006, vol. 122, p. 124.
Kakarougkas, A., Ismail, A., Chambers, A.L., Riballo, E., Herbert, A.D., Künzel, J., Löbrich, M., Jeggo, P.A., and Downs, J.A., Requirement for PBAF in transcriptional repression and repair at DNA breaks in actively transcribed regions of chromatin, Mol. Cell, 2014, vol. 55, p. 723.
Liu, G., Stevens, J.B., Horne, S.D., Abdallah, B.Y., Ye, K.J., Bremer, S.W., Ye, C, J., Chen, D.J., and Heng, H.H., Genome chaos, survival strategy during crisis, Cell Cycle, 2014, vol. 13, p. 528. https://doi.org/10.4161/cc.27378
Ly, P. and Cleveland, D.W., Rebuilding chromosomes after catastrophe: emerging mechanisms of chromothripsis, Trends Cell Biol., 2017, vol. 27, p. 917. https://doi.org/10.1016/j.tcb.2017.08.005
Mamaeva, S.E., Regularities of cell karyotypic evolution in culture, Tsitologiia, 1996, vol. 38, p. 787.
Mamaeva, S.E., Chromosomes of human permanent cell lines, in Atlas khromosom postoyannykh kletochnykh liny cheloveka i zhivotnykh (Atlas of Chromosomes of Human and Animal Cell Lines), Moscow: Nauchnyi Mir, 2002, p. 36.
Mardin, B.R., Drainas, A.P., Waszak, S.M., Weischenfeldt, J., Isokane, M., Stütz, A, M., Raeder, B., Efthymiopoulos, T., Buccitelli, C., Segura-Wang, M., Northcott, P., Pfister, S.M., Lichter, P., Ellenberg, J., and Korbel1, J.O., A cell-based model system links chromothripsis with hyperploidy, Mol. Syst. Biol., 2015, vol. 11, p. 828. https://doi.org/10.15252/msb.20156505
Mitelman, F., Recurrent chromosome aberrations in cancer, Mutat. Res., 2000, vol. 462, p. 247.
Mitelman, F., Johansson, B., and Mertens, F., The impact of translocations and gene fusions on cancer causation, Nat. Rev. Cancer, 2007, vol. 7, p. 233.
Ohgaki, H. and Kleihues, P., Genetic alterations and signaling pathways in the evolution of gliomas, Cancer Sci., 2009, vol. 100, p. 2235.
Orsolic, I., Jurada, D., Pullen, N., Oren, M., Eliopoulos, A.G., and Volarevic, S., The Relationship between the nucleolus and cancer: current evidence and emerging paradigms, Semin. Cancer Biol., 2015. pii: S1044-579X(15)30004-3. https://doi.org/10.1016/j.semcancer.2015.12.004
Ozkinay, C. and Mitelman, F., A simple trypsin–Giemsa technique producing simultaneous G- and C-banding in human chromosomes, Hereditas, 1979, vol. 90, p. 1.
Pellestor, F.C., Chromoanagenesis: cataclysms behind complex chromosomal rearrangements, Mol. Cytogenet., 2019, vol. 12, p. 6. https://doi.org/10.1186/s13039-019-0415-7
Polyanskaya, G.G., Abramyaqn, D.S., and Glebov, O.K., The karyotypic structure of clonal population of Chinese hamster cells during a prolonged cultivation, Tsitologiia, 1981, vol. 23, p. 818.
Satoh, H., Yoshida, M.S., and Sasaki, M., Resolution chromosome banding in the Norway rat, Rattus norvegicus, Cytogenet. Cell Genet., 1989, vol. 50, p. 151.
Tuna, M., Knuutila, S., and Mills, G.B., Uniparental disomy in cancer, Trends Mol. Med., 2009, vol. 15, p. 120.
Yartseva, N.M., Fedortseva, R.F., and Artsybasheva, I.V., Chromosomal rearrangements and their effects in spontaneous immortalization and transformation of rat embryo cells in vivo, Tsitologiia, 2007, vol. 49, p. 311.
Yartseva, N.M., Bykova, T.V., Zubova, S.G., Pospelov, V.A., and Pospelova, T.V., Chromosomal instability and evolution of transformed phenotypein cell lines selected from senescent rat embryonic fibroblasts with rapamycin, Cell Tissue Biol., 2019, vol. 13, p. 18. https://doi.org/10.1134/S1990519X19010103
Ye, C.J., Regan, S., Liu, G., Alemara, S., and H.Heng, H., Understanding aneuploidy in cancer through the lens of system inheritance, fuzzy inheritance and emergence of new genome systems, Mol. Cytogenet., 2018, vol. 11, p. 31. https://doi.org/10.1186/s13039-018-0376-2
Ye, C.J., Sharpe, Z., Alemara, S., Mackenzie, S., Liu, G., Abdallah, B., Horne, S., Regan, S., and Heng, H.H., Micronuclei and genome chaos: changing the system inheritance, Genes, 2019, vol. 10, p. 366. https://doi.org/10.3390/genes10050366
Zhang, C.Z., Spektor, A., Cornils, H., Francis, J.M., Jackson, E, K., Liu, S., Meyerson, M., and Pellman, D., Chromothripsis from DNA damage in micronuclei, Nature, 2015, vol. 522, p. 179. https://doi.org/10.1038/nature14493
Funding
This work was carried out thanks to budgetary funding on a planned theme of the Institute of Cytology, Russian Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflict of interest.
This study did not performed experiments with animals or human beings.
Additional information
Abbreviations: ARG—amplification of ribosomal genes DSBs—double-strand breaks; SRC—structural rearrangement of chromosomes; NC—number of chromosome.
Rights and permissions
About this article

Cite this article
Yartseva, N.M., Shitikova, Z.V., Bykova, T.V. et al. Karyotypic Changes of Apoptosis-Resistant Rat Cells HindIIIG during Prolonged Cultivation after Exposure to Ionizing Irradiation. Cell Tiss. Biol. 15, 248–259 (2021). https://doi.org/10.1134/S1990519X21030135
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1990519X21030135