
Abstract
Bax inhibitor 1 (BI-1) is a highly conserved protein originally identified as a suppressor of the proapoptotic protein Bax to inhibit cell death in animals and plants. The orthologs of BI-1 are widely distributed in filamentous fungi but their functions remain largely unknown. Herein, we report the identification and characterizations of MrBI-1, an ortholog of BI-1, in the entomopathogenic fungus Metarhizium robertsii. First, we found that MrBI-1 could partially rescue mammalian Bax-induced cell death in yeast. Deletion of MrBI-1 impaired fungal development, virulence and heat tolerance in M. robertsii. We also demonstrated that inactivation of MrBI-1 reduced fungal resistance to farnesol but not to hydrogen peroxide, suggesting that MrBI-1 contributes to antiapoptotic-like cell death via the endoplasmic reticulum stress-signaling pathway rather than the classical mitochondrium-dependent pathway. In particular, we found that unlike the observations in yeasts and plants, expression of mammalian Bax did not lead to a lethal effect in M. robertsii; however, it did aggravate the fungal apoptotic effect of farnesol. The results of this study advance our understanding of BI-1-like protein functions in filamentous fungi.
Similar content being viewed by others


Introduction
Programmed cell death (PCD) is a genetically controlled and highly conserved process occurring in different organisms and include two categories, i.e. apoptosis (type I PCD) and autophagy (type II PCD)1. Both types have been well documented in fungi2. For cellular apoptosis, the mitochondrial (MT) pathways with the typical feature of cytochrome c release have been described in different filamentous fungi such as Aspergillus nidulans3,4,5,6, Neurospora crassa7 and Fusarium graminearum8. The identifications and subsequent characterizations of Bcl-2 family members, e.g. Bcl-2, Bax, Bak, etc., in animals revealed the endoplasmic reticulum (ER) pathways of apoptosis9. Bcl-2 family proteins are not present in fungi; however, exogenous expression of proapoptotic factor Bax or Bak could induce typical apoptotic characteristics, whereas the expression of antiapoptotic factor Bcl-2 could inhibit cell death in both budding and fission yeasts10,11, indicating the presence of conserved apoptotic machineries in fungi.
Bax inhibitor-1 (BI-1) was first identified in mammals but functionally verified in yeast and shown to suppress Bax-induced cell death10. Unlike the Bcl-2 family members, BI-1 is evolutionarily conserved and widely present in eukaryotic species from fungi to plants and animals12. BI-1 proteins of different organisms have either six or seven transmembrane domains and are predominantly localized to the ER membrane10,13. It has been demonstrated that BI-1 can interact with Bcl-2 family members so as to enable the cells to adapt to a wide range of stresses via ER-signaling pathways14,15,16. Homologs of BI-1 have also been cloned and characterized in plants, e.g. AtBI-1 from Arabidopsis17,18,19 and BXI1 from the budding yeast Saccharomyces cerevisiae20. AtBI-1 can interact with different proteins to activate the hypersensitive response with PCD-type plant epidermal cell death against fungal pathogen infections21,22. Yeast BXI1 is an ER-localized protein and the gene deletion mutant has become more sensitive to ER-stress drugs20. Unlike fungi, plants encode a family of ER-residing Bag (Bcl-2 associated anthogene) proteins that can interact with different proteins to regulate the apoptosis-like processes ranging from abiotic stresses to pathogen attacks23. Prior to this study, the distribution and function(s) of BI-1 homologs remained unknown in filamentous fungi.
Ascomycete insect pathogenic fungi such as Metarhizium spp. diverged after the yeasts24 and these species have been studied as model organisms for underpinning the mechanisms of insect-fungus interactions25,26. Various virulence-related genes have been functionally characterized, including the autophagy-related genes27,28. Oxidative stress related MT-signaling pathways have been evident with an apoptosis-like aging in Metarhizium species3,29,30. In the present work, an ortholog of yeast BXI1 gene, designated MrBI-1 (MAA_10304, 30% identity at the protein sequence level), was identified and deleted in M. robertsii. We found that MrBI-1 partially rescued Bax-induced lethal effects in yeasts and that the protein contributed to heat tolerance and antiapoptotic-like cell death in M. robertsii. Deletion of MrBI-1 also impaired fungal virulence.
Results
Phylogeny and secondary structure analyses
Genome-wide surveys of different ascomycete fungal genomes indicated that as in mammals, plants and yeasts, a single copy of a BI-1-like gene is present in M. robertsii and other ascomycetous, basidiomycetous and zygomycetous fungi. The extensive genome surveys indicated that the BI-1-like proteins are not present in the basal fungal genomes of chytrids and microsporidians. A phylogenetic analysis demonstrated that MrBI-1 is mostly closely related to the homolog (MAC_04368) from M. acridum (a locust-specific pathogen) and that it was then clustered with those from hypocrealean fungi (Fig. 1A). In general, except for the yeast BXI1, the phylogeny of BI-1 proteins is congruent with organism speciation24, an indicator of a highly conserved relationship among these orthologs. Scanning of the transmembrane domain revealed that, in contrast to human BI-1, MrBI-1 contains seven transmembrane domains like the orthologs from yeast (BXI1) and Arabidopsis (AtBI-1) (Fig. 1B).

Phylogenetic analysis and transmembrane domain comparison. (A) Phylogenetic analysis of MrBI-1 (highlighted in bold) with different homologs. The protein sequences were aligned with CLUSTAL X and a Maximum likelihood tree was generated using a Dayhoff substitution model. (B) Schematic comparison of transmembrane domains present in MrBI-1, yeast BXI1, Arabidopsis AtBI-1 and human BI-1.
MrBI-1 partially rescued the Bax-induced growth defect in yeast
The yeast strains harboring either an empty vector or the plasmid containing a galactose-inducible promoter-controlled Bax gene, i.e. the ZD09001 strain, grew equally well on a glucose-containing synthetic drop-out (SD) medium. However, once induced on a galactose medium, ZD09001 cells lost their viability due to the lethal effect of Bax (Fig. 2A). To determine whether MrBI-1 could suppress Bax-induced cell death in yeast, the cDNA of MrBI-1 as well as the positive controls of mammalian Bcl-2 and Arabidopsis AtBI-1 genes were used for yeast transformations. All yeast strains obtained grew equally well on the glucose medium. However, once Bax protein expression was induced by galactose, in contrast to the negative control, the Bcl-2-containing strain showed complete viability; while similar to the AtBI-1 -transformed strain, MrBI-1 partially suppressed Bax-induced cell death in yeast (Fig. 2B).

Functional assay of MrBI-1 in yeasts. (A) Yeast strain BF264-15Dau was transformed with the vector pTS909-Bax and the resultant strain ZD09001 was either streaked on a SD-Trp (Glu) or on an SD-Trp (Gal) plate and incubated at 30 °C for 3 d. Glu, glucose; Gal, galactose. All strains grew equally well on a SD-Trp (Glu) plate while the ZD09001 cells lost their viability when streaked on an SD-Trp (Gal) plate. (B) ZD09001 was transformed by various vectors (pYX112-Bcl2, pYX112-AtBI, and pYX112-MrBI, pYX112 as control). Yeast strains of 5 μl by gradient were spotted on either SD-Trp-Ura (Glu) or SD-Trp-Ura (Gal) plates and incubated at 30 °C for 3 d. The viabilities of the yeast cells containing Bcl-2, MrBI-1 and AtBI-1 genes were increased to varied degrees when compared with the control strain transformed with the empty vector.
Gene deletion and phenotypic characterization
To investigate the potential function of MrBI-1 in M. robertsii, the gene was deleted by homologous recombination. Gene complementation was performed by transformation of ΔMrBI-1 with the MrBI-1 cDNA to obtain the mutant Comp. Genetically stable transformants were verified by PCR (Fig. 3A) and RT-PCR (Fig. 3B). In terms of the growth rate, no obvious difference was observed when growing the wild-type (WT) and mutants on potato dextrose agar (PDA) (Figs. 3C and 4A). As indicated above, the proapoptotic Bax-like factor is not present in fungi or plants31. To further determine the function of MrBI-1, the mammalian Bax gene was made under the control of a constitutive promoter and used to transform the WT and ΔMrBI-1 strains. The successfully obtained transformants were designated WT::Bax and ΔMrBI-1::Bax, respectively. When grown on PDA, no growth rate variations were observed for these mutants when compared to the WT of M. robertsii (Fig. 4A). Our RT-PCR analysis confirmed that the exogenous Bax gene could be similarly expressed by the fungi (Fig. 4B).

Verification of fungal transformation and phenotypic characterization. (A) PCR verification. The plasmid (PL) pBarMrBI was used as a positive control. (B) RT-PCR verification to confirm the loss and recovery of MrBI-1 transcripts. The small subunit ribosomal gene (18 S rRNA) was used as a reference. (C) Growth characteristics of WT, ΔMrBI-1, and Comp on PDA for 15 d. No growth-rate variation was observed among the WT, ΔMrBI-1 and comp.

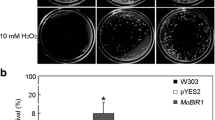
Growth test and spore survival assays against H2O2 challenge. (A) Phenotypes of the WT, ΔMrBI-1, Comp, WT::Bax and ΔMrBI-1::Bax on PDA or PDA supplemented with 20 or 40 mM H2O2 for 10 d. No obvious difference in growth rate was observed between the WT and mutants. (B) Verification of exogenous Bax gene expression. The strains were grown in SDB for 3 d and used for RT-PCR analysis to verify the transcription of Bax gene in WT::Bax and ΔMrBI-1::Bax. The 18 S rRNA gene was used as a reference. (C) Germination rates of different strain conidia in the presence of H2O2. The conidia of the WT, ΔMrBI-1, Comp, WT::Bax and ΔMrBI-1::Bax were suspended in SDB with 1 or 2 mM H2O2 for 12 h.
Deletion of MrBI-1 increased fungal sensitivity to heat shock but not to H2O2
To examine the effect of MrBI-1 on different stress factors, the spores of the WT and mutants were challenged with H2O2 or heat shock. The growth of mutants was not inhibited when the fungi were grown on PDA or PDA supplemented with H2O2 as compared to the WT of M. robertsii (Fig. 4A). During germination assays, the survival rates of the WT and mutant spores were equally reduced after the treatment with H2O2, i.e. the viabilities of mutant spores were not significantly different from those of the WT (Fig. 4C). This is consistent with the report in yeasts that BXI1 is not involved in suppressing H2O2-induced cell death12. Heat shock treatment indicated, however, that significant differences in conidial survival rates were observed between the WT and mutants after exposure to 45 °C for 1 or 2 h (Fig. 5A). For example, the viabilities of ΔMrBI-1 spores were significantly reduced when treated either for 1 h (t-test, P = 0.0311) or 2 h (P = 0.0038) when compared with the WT. In addition, relative to the WT, overexpression of Bax in ΔMrBI-1 but not in the WT further increased fungal sensitivity to heat shock (P = 0.0238 for treatment for 1 h; P = 0.0072 for 2 h). The differences of spore survival were not significant between ΔMrBI-1 and ΔMrBI-1::Bax after exposure to 45 °C for 1 (P = 0.2152) but significant after exposure for 2 h (P = 0.0282) (Fig. 5A). These observations thereby indicated that MrBI-1 was not involved in the oxidative stress response but in heat tolerance in M. robertsii.

Spore survival assays against heat shock and FOH challenges. (A) Spore germinations after heat shock treatment. Conidia of the WT, ΔMrBI-1, Comp, WT::Bax and ΔMrBI-1::Bax were treated at 45 °C in a water bath for different time periods and inoculated on PDA at 25 °C for 24 h. Conidial survival rate of different mutants varied to different degrees when compared with the WT. (B) Spore germinations in the presence of FOH. The conidia of the WT, ΔMrBI-1, Comp, WT::Bax and ΔMrBI-1::Bax were suspended in SDB with 50 μM FOH at 25 °C for 16 h. Conidial survival rate of different mutants varied to different degrees when compared with the WT.
MrBI-1 contributes to antiapoptotic-like cell death in M. robertsii
To determine whether MrBI-1 contributed to the suppression of apoptotic cell death in M. robertsii, fungal cells were treated with the apoptosis-inducing compound farnesol (FOH)32. After exposure to 25 or 50 μM FOH for 4 h, the WT germlings were co-stained with Hoechst and propidium iodide (PI) dyes. The observations revealed the apoptotic features of intense chromatin condensation and marginalization in the cells treated with 25 μM FOH. The exposure to 50 μM FOH even triggered cell necrosis as revealed by PI staining signals (Fig. 6). Based on these results, the contribution of MrBI-1 to the suppression of FOH-induced apoptotic-like cell death was evaluated by inoculation of the WT and different mutant spores in Sabouraud dextrose broth (SDB) in the presence or absence of FOH (50 μM) for 16 h (Fig. 5B). When compared with the WT (91.8%), spore germination rates of ΔMrBI-1 (69.5%, P = 0.0111), WT::Bax (36.9%, P = 0.0007) and ΔMrBI-1::Bax (47.4%, P = 0.0035) were significantly reduced. No significant difference was observed between WT and Comp. The differences between the WT and WT::Bax (P = 0.0007) and between the ΔMrBI-1 and ΔMrBI-1::Bax (P = 0.0285) were also significant; however, there was no significant difference between WT::Bax and ΔMrBI-1::Bax (P = 0.1019) (Fig. 5B).

Apoptosis induction in M. robertsii. Germlings of the WT were treated with or without FOH (25 or 50 μM) at 25 °C for 4 h and double stained with Hoechst 33342/PI dyes. Bright field (BF) optics showed a dose-dependent inhibition of hyphal growth. The cells treated with 25 μM FOH displayed the apoptotic characteristic of chromatin condensation and marginalization characteristic of apoptosis, whereas the germlings treated with 50 μM FOH were infiltrated by PI, indicative of cell necrosis. Bar = 10 μm.
MrBI-1 is required for full virulence in M. robertsii
To investigate the effects of MrBI-1 deletion on fungal virulence, insect bioassays were conducted against silkworm larvae. The median lethal time (LT50) values were estimated and compared among the WT, ΔMrBI-1 and Comp. The results indicated that the differences between the WT (LT50 = 2.5 ± 0.093 days) and ΔMrBI-1 (LT50 = 3.0 ± 0.284 days) and between ΔMrBI-1and Comp (LT50 = 2.5 ± 0.186 days) were significant (P < 0.05); but that this was not the case between the WT and Comp. The results thereby indicated that deletion of MrBI-1 impaired full virulence in M. robertsii.
Discussion
In the present study, we characterized a bona fide homolog of BI-1, MrBI-1, in the insect pathogenic fungus M. robertsii. Not surprisingly, MrBI-1 was able to suppress Bax-induced cell death in yeast to some extent. Deletion of MrBI-1 revealed that MrBI-1 is required for fungal heat tolerance, full virulence and contributes to ER-stress inducer FOH, but not to the MT-sensitive factor H2O2. Human BI-1 was first identified as being able to rescue yeast cells containing the proapoptotic factor Bax10. Those experiments demonstrated that the endogenous BI-1 gene (i.e. BXI1) of yeast could not counteract the apoptotic effect caused by mammalian Bax. In support of this, our experiments indicated that unlike Bcl-2, both MrBI-1 and AtBI-1 could only partially rescue Bax-transformed yeast cells, even when integrated with BXI1’s function (Fig. 2B). Given that the orthologs of BI-1 are highly conserved (Fig. 1), the results of protein interaction assays revealed that the Arabidopsis AtBI-1 interacted with cytochrome b5 in plants, but interacted with a fatty acid hydroxylase in yeast to mediate the hydroxylation of fatty acids33. This would explain much of why neither AtBI-1 nor MrBI-1 failed to completely inhibit the apoptotic effect of Bax, as occurs with Bcl-2 in yeast.
FOH is a 15-carbon isoprenoid alcohol that is widely distributed in nature as an odoriferous compound and acts as a precursor in the isoprenoid/sterol biosynthetic pathway34. FOH has been reported to induce apoptosis-like PCD in different fungal species, e.g. S. cerevisiae, A. nidulans, and Candida albicans10. In the present study, we established that FOH triggered cellular apoptosis and even necrosis at a higher concentration in M. robertsii. Observations in mammalian cells indicated that deletion of BI-1 did not increase cell sensitivity to MT stress, but did against ER-stress agents35,36. Consistent with this, our results showed that MrBI-1 was not involved in H2O2-induced growth defects or cell death in M. robertsii (Fig. 4). Thus, we assert that MrBI-1 contributes to antiapoptotic effects in M. robertsii via an ER but not MT stress-response pathway. However, the exact mechanism remains to be elucidated.
Experiments in yeasts11,33 and plant cells37 indicated that heterologous expression of mammalian Bax could result in cell death even with the presence of endogenous BI-1 genes. This was also evident in the filamentous plant pathogenic fungus Colletotrichum gloeosporioides38. In contrast (and unexpectedly), expression of Bax did not induce cell death in either the WT or ΔMrBI-1 of M. robertsii (Fig. 4A). In animal cells, Bax forms a homodimer, multidimer or heterodimer with Bcl-2, while BI-1 can interact with Bcl-2 but not Bax or Bak10,39. Co-expression of Bcl-2 and Bax in C. gloeosporioides did not lead to cell death38. The lack of Bax and Bcl-2 in fungal and plant cells suggests that the heterodimer of Bax and Bcl-2 could not be formed in these heterologous systems. Thus, the disparate non-lethal effect of Bax in Metarhizium suggests that much, if not all, of the aggregation status of Bax varied and thereby played distinct roles in different filamentous fungi, yeasts and plant cells. Consistent with a non-lethal effect, the heat-tolerant abilities of WT::Bax and ΔMrBI-1::Bax mutants were not reduced when compared with either the WT or ΔMrBI-1 (Fig. 5A). However, relative to the WT and ΔMrBI-1, the germination rates of WT::Bax and ΔMrBI-1::Bax spores were significantly reduced when exposed to FOH (Fig. 5B), indicating that Bax aggravated the apoptotic effect in M. robertsii under the stress agent. Different from the yeast assay, the erratic results between the heat shock and FOH induction would suggest that MrBI-1 might not fully function via a Bax-inhibitor mechanism on the basis that Bax is evolutionarily missing in M. robertsii. For example, the spores of ΔMrBI-1::Bax had a higher (P = 0.0282) survival rate than those of ΔMrBI-1 under heat shock for 2 h, while the spore survival rates between the WT::Bax and ΔMrBI-1::Bax were significantly different when the fungi were exposed to heat shock for 2 h (P = 0.0095) but not to FOH (P = 0.1019) (Fig. 5). The exact mechanism(s) involved remains to be elucidated in terms of the disparity of the Bax effects on cell viability between M. robertsii and yeasts or plants.
Pathogenic infections can trigger a hypersensitive response (HR) in plants, including the generation of an oxidative burst that is toxic to both pathogen and plant cells and thereby the rapid cell death in the region surrounding an infection40. In Arabidopsis, AtBI-1 has been functionally implicated in the regulation of cell death in response to fungal infection22. These pathogens, however, secrete avirulence effector(s) and other pathogenicity-related proteins to counteract plant PCD-like immunities so as to allow for successful infection41. An HR-like response of melanization and cell death also occurs in insects when infected by M. robertsii during cuticle penetration42. After reaching the insect hemocoel, fungal pathogens such as Metarhizium species have to adapt to osmotic stress in hemolymph43 and counteract antifungal effects imposed by host cellular and humoral immune responses26. Thus, the attenuated virulence of ΔMrBI-1 would then be expected to be due to its reduced resistance to cell death triggered by insect immune reactions when compared to the WT.
In conclusion, we herein report the function of MrBI-1 in an insect pathogenic fungus M. robertsii. Heterologous expression of MrBI-1 partially suppressed the lethal effect of mammalian Bax in yeast. In contrast to the observations in yeast and plant cells, Bax could not induce cell death in M. robertsii. Our data indicate for the first time that MrBI-1 is involved in ER-related apoptosis-like PCD, which contributes to fungal development and virulence.
Methods
Fungal strains and growth conditions
The WT strain and transformants of M. robertsii ARSEF 2575 (previously classified as M. anisopliae) were routinely cultured on PDA (Difco) at 25 °C. For liquid incubation, fungi were grown in SDB (Difco) at 25 °C in a rotatory shaker. Conidium suspensions were prepared in 0.05% (v/v) Tween-80 and filtered through four layers of sterile lens-cleaning tissues to remove hyphal fragments. Yeast strains used in this study were cultured on different media including yeast extract peptone dextrose (YPD, 1% yeast extract, 2% peptone and 2% glucose), YPDA (YPD plus 0.008% adenine) and synthetic drop-out medium (0.17% yeast nitrogen base, amino acids and different carbon sources, with or without urea).
Phylogenetic and transmembrane domain analyses
To establish the phylogenetic relationship between MrBI-1 and its orthologs, orthologous protein sequences from representative fungal species and those of mouse, human and A. thaliana were aligned using the program CLUSTAL X44. A Maximum likelihood tree was generated using the program MEGA (ver. 6.0)45 with a Dayhoff substitution model, a Nearest-Neighbor-Interchange heuristic method for tree inference, 1,000 bootstrap replications for phylogeny test and a partial deletion for gaps/missing data. Transmembrane domains of MrBI-1, yeast BXI1, Arabidopsis AtBI-1 and human BI-1 were analyzed on the TMHMM server v. 2.0 ( http://www.cbs.dtu.dk/services/TMHMM/).
Gene function assays in yeasts
The coding region of the mammalian apoptotic gene Bax from the rat Rattus norvegicus was introduced into a HindIII/SalI–digested yeast expression vector pTS909 under the control of the GAL1 promoter33. The resultant vector pTS909-Bax was used to transform the yeast strain BF264-15Dau using a lithium acetate method46. The resultant strain, named ZD09001, was maintained in a SD medium lacking tryptophan (SD-Trp). The cDNA of MrBI-1 was cloned into the yeast vector pYX112 under the control of the triosephosphate isomerase gene (TPI) promoter to produce pYX112-MrBI. The coding regions of the Bax inhibitor gene Bcl-2 from R. norvegicus47 and AtBI-1 from the plant A. thaliana13 were cloned and used to generate the plasmids pYX112-Bcl-2 and pYX112-AtBI, respectively. These three plasmids (pYX112-MrBI, pYX112-Bcl-2 and pYX112-AtBI1) and the control vector pYX112 were transformed into the yeast strain ZD09001. Transformants were spotted either on an SD-glucose plate or on an SD-galactose plate and incubated at 30 °C for 3 d.
Gene deletion and complementation
Targeted gene deletion of MrBI-1 gene was performed by homologous recombination as we described previously48. Briefly, the two primer pairs MrBI1U/MrBI1L and MrBI2U/MrBI2L (Table 1) were used to amplify the 5′- and 3′-flanking regions of the target gene MrBI-1, respectively. The PCR products were digested with the restriction enzymes BamHI and XbaI, respectively and then inserted into the corresponding sites of the binary vector pDHt-bar (conferring resistance against ammonium glufosinate) to generate the plasmid pBarMrBI for Agrobacterium-mediated fungal transformation (ATMT). For gene complementation, the binary vector pBenMrBI was constructed by inserting the full cDNA sequence of MrBI-1 into the plasmid pDHt-ben-gpdA (conferring resistance against Benomyl) under control of the Aspergillus nidulans GPDA promoter (gpdA)27 and transformed into the MrBI-1 null mutant by ATMT to generate the complementary mutant (Comp). For ectopic expression of Bax in M. robertsii, the full-length cDNA of murine Bax gene was amplified by PCR with the primers BaxF and BaxR using the plasmid pTS909-Bax DNA as a template. The purified fragment was cloned into the EcoRI site of the pGPDBen vector to yield the pGPDBen-Bax expression vector, which was then used for ATMT transformation of the WT and ΔMrBI-1 to generate the mutants WT::Bax and ΔMrBI-1::Bax, respectively. To confirm successful gene deletion and complementation, PCR and RT-PCR were performed using the primers MrBIF/MrBIR. To verify the successful expression of Bax, the fungi were grown in SDB for 3 d and used for RT-PCR analysis with primers BaxRT1 and BaxRT1 (Table 1). The 18S rRNA gene was amplified using primers 18SF and 18SR (Table 1) and used as an internal positive control49.
Heat shock assay
Conidia of the WT, ΔMrBI-1, Comp, WT::Bax and ΔMrBI-1::Bax were collected in 0.05% (v/v) Tween-80 and the spore suspensions (1 ml, 2 × 107 conidia/ml) were transferred into Eppendorf tubes for treatment at 45 °C in a water bath for different time periods. The aliquots of suspensions (10 μl) were inoculated onto PDA medium (3 ml in plastic Petri dishes, 6 cm diameter) and the plates were incubated at 25 °C for 24 h for examination of conidium survivals. There were three replicated plates per strain and the experiment was repeated twice.
Conidial stress challenges and survival assays
For cell death analysis, WT germlings were treated with or without the apoptosis-inducing compound farnesol (FOH, Sigma, prepared in dimethyl sulfoxide to final concentrations of 25 or 50 μM) at 25 °C for 4 h34. The dyes Hoechst 33342 and PI (Beyotime Ltd.) were used to stain cell nuclei and necrotic cells, respectively3. Observations using fluorescence microscopy were made with an Olympus microscope (BX51-33P, Olympus). The effect of FOH on cell viability was assayed by inoculation of the spores of WT and different mutants in SDB with or without (control) 50 μM FOH for 16 h or H2O2 (1 or 2 mM) for 12 h at 25 °C to determine spore germination rates. There were three replicated plates per strain and the experiments were repeated twice.
Insect bioassays
To investigate the effect of MrBI-1 on fungal virulence, insect bioassays were conducted against the newly emerged fifth instar silk worm, Bombyx mori. Conidia of the WT, ΔMrBI-1 and Comp harvested from the PDA plates were applied topically by immersing the larvae for 30 seconds in an aqueous suspension containing 2 × 107 conidia/ml. Each treatment had three replicates with 15 insects each and the experiments were repeated twice. Mortality was recorded every 12 h. The values of median lethal time (LT50) were calculated for each strain by Kaplan-Meier analysis50.
Additional Information
How to cite this article: Chen, Y. et al. The Bax inhibitor MrBI-1 regulates heat tolerance, apoptotic-like cell death and virulence in Metarhizium robertsii. Sci. Rep. 5, 10625; doi: 10.1038/srep10625 (2015).
References
Hamann, A., Brust, D. & Osiewacz, H. D. Apoptosis pathways in fungal growth, development and ageing. Trends Microbiol. 16, 276–283 (2008).
Sharon, A., Finkelstein, A., Shlezinger, N. & Hatam, I. Fungal apoptosis: function, genes and gene function. FEMS Microbiol. Rev. 33, 833–854 (2009).
Li, L. et al. Linkage of oxidative stress and mitochondrial dysfunctions to spontaneous culture degeneration in Aspergillus nidulans. Mol. Cell Proteomics 13, 449–461 (2014).
Cheng, J., Park, T. S., Chio, L. C., Fischl, A. S. & Ye, X. S. Induction of apoptosis by sphingoid long-chain bases in Aspergillus nidulans. Mol. Cell Biol. 23, 163–177 (2003).
Leiter, E. et al. Antifungal protein PAF severely affects the integrity of the plasma membrane of Aspergillus nidulans and induces an apoptosis-like phenotype. Antimicrob Agents Chemother 49, 2445–2453 (2005).
Semighini, C. P., Hornby, J. M., Dumitru, R., Nickerson, K. W. & Harris, S. D. Farnesol-induced apoptosis in Aspergillus nidulans reveals a possible mechanism for antagonistic interactions between fungi. Mol. Microbiol. 59, 753–764 (2006).
Marek, S. M., Wu, J., Louise Glass, N., Gilchrist, D. G. & Bostock, R. M. Nuclear DNA degradation during heterokaryon incompatibility in Neurospora crassa. Fungal Genet. Biol. 40, 126–137 (2003).
Semighini, C. P., Murray, N. & Harris, S. D. Inhibition of Fusarium graminearum growth and development by farnesol. FEMS Microbiol. Lett. 279, 259–264 (2008).
Breckenridge, D. G., Germain, M., Mathai, J. P., Nguyen, M. & Shore, G. C. Regulation of apoptosis by endoplasmic reticulum pathways. Oncogene 22, 8608–8618 (2003).
Xu, Q. & Reed, J. C. Bax inhibitor-1, a mammalian apoptosis suppressor identified by functional screening in yeast. Mol. Cell 1, 337–346 (1998).
Jürgensmeier, J. M. et al. Bax- and Bak-induced cell death in the fission yeast Schizosaccharomyces pombe. Mol. Biol. Cell 8, 325–339 (1997).
Chae, H. J. et al. Evolutionarily conserved cytoprotection provided by Bax Inhibitor-1 homologs from animals, plants and yeast. Gene 323, 101–113 (2003).
Kawai, M., Pan, L., Reed, J. C. & Uchimiya, H. Evolutionally conserved plant homologue of the Bax inhibitor-1 (BI-1) gene capable of suppressing Bax-induced cell death in yeast. FEBS Lett. 464, 143–147 (1999).
Ishikawa, T., Watanabe, N., Nagano, M., Kawai-Yamada, M. & Lam, E. Bax inhibitor-1: a highly conserved endoplasmic reticulum-resident cell death suppressor. Cell Death Differ. 18, 1271–1278 (2011).
Robinson, K. S., Clements, A., Williams, A. C., Berger, C. N. & Frankel, G. Bax inhibitor 1 in apoptosis and disease. Oncogene 30, 2391–2400 (2011).
Lisbona, F. et al. BAX inhibitor-1 is a negative regulator of the ER stress sensor IRE1alpha. Mol. Cell 33, 679–691 (2009).
Yue, H., Li, Z. & Xing, D. Roles of Arabidopsis bax inhibitor-1 in delaying methyl jasmonate-induced leaf senescence. Plant Signal Behav. 7, 1488–1489 (2012).
Wang, X. et al. Wheat BAX inhibitor-1 contributes to wheat resistance to Puccinia striiformis. J. Exp. Bot. 63, 4571–4584 (2012).
Kawai-Yamada, M., Jin, L., Yoshinaga, K., Hirata, A. & Uchimiya, H. Mammalian Bax-induced plant cell death can be down-regulated by overexpression of Arabidopsis Bax Inhibitor-1 (AtBI-1). Proc. Natl. Acad. Sci. USA 98, 12295–12300 (2001).
Cebulski, J., Malouin, J., Pinches, N., Cascio, V. & Austriaco, N. Yeast Bax inhibitor, Bxi1p, is an ER-localized protein that links the unfolded protein response and programmed cell death in Saccharomyces cerevisiae. PloS One 6, e20882 (2011).
Weis, C. et al. Co-immunoprecipitation-based identification of putative BAX INHIBITOR-1-interacting proteins involved in cell death regulation and plant-powdery mildew interactions. Mol. Plant Pathol. 14, 791–802 (2013).
Weis, C. et al. CYP83A1 is required for metabolic compatibility of Arabidopsis with the adapted powdery mildew fungus Erysiphe cruciferarum. The New phytologist 202, 1310–1319, doi:10.1111/nph.12759 (2014).
Doukhanina, E. V. et al. Identification and functional characterization of the BAG protein family in Arabidopsis thaliana. J. Biol. Chem. 281, 18793–18801 (2006).
Hu, X. et al. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 111, 16796–16801 (2014).
St Leger, R. J. & Wang, C. Genetic engineering of fungal biocontrol agents to achieve greater efficacy against insect pests. Appl. Microbiol. Biotechnol. 85, 901–907 (2010).
Wang, C. & Feng, M.-G. Advances in fundamental and applied studies in China of fungal biocontrol agents for use against arthropod pests. Biological Control 68, 129–135 (2014).
Duan, Z. et al. Linkage of autophagy to fungal development, lipid storage and virulence in Metarhizium robertsii. Autophagy 9, 538–549 (2013).
Zhang, L. et al. The autophagy gene BbATG5, involved in the formation of the autophagosome, contributes to cell differentiation and growth but is dispensable for pathogenesis in the entomopathogenic fungus Beauveria bassiana. Microbiology 159, 243–252 (2013).
Wang, C., Butt, T. M. & St Leger, R. J. Colony sectorization of Metarhizium anisopliae is a sign of ageing. Microbiology 151, 3223–3236 (2005).
Li, L., Pischetsrieder, M., St Leger, R. J. & Wang, C. Associated links among mtDNA glycation, oxidative stress and colony sectorization in Metarhizium anisopliae. Fungal Genet Biol. 45, 1300–1306 (2008).
Watanabe, N. & Lam, E. Bax Inhibitor-1, a conserved cell death suppressor, is a key molecular switch downstream from a variety of biotic and abiotic stress signals in plants. Int. J. Mol. Sci. 10, 3149–3167 (2009).
Liu, P. et al. Farnesol induces apoptosis and oxidative stress in the fungal pathogen Penicillium expansum. Mycologia 102, 311–318 (2010).
Nagano, M. et al. Functional association of cell death suppressor, Arabidopsis Bax inhibitor-1, with fatty acid 2-hydroxylation through cytochrome b5 . Plant J 58, 122–134 (2009).
Edwards, P. A. & Ericsson, J. Sterols and isoprenoids: signaling molecules derived from the cholesterol biosynthetic pathway. Annu. Rev. Biochem. 68, 157–185 (1999).
Chae, H. J. et al. BI-1 regulates an apoptosis pathway linked to endoplasmic reticulum stress. Mol. Cell. 15, 355–366 (2004).
Joo, J. H. & Jetten, A. M. Molecular mechanisms involved in farnesol-induced apoptosis. Cancer Lett. 287, 123–135 (2010).
Yoshinaga, K. et al. Mammalian Bax initiates plant cell death through organelle destruction. Plant Cell Rep. 24, 408–417 (2005).
Barhoom, S. & Sharon, A. Bcl-2 proteins link programmed cell death with growth and morphogenetic adaptations in the fungal plant pathogen Colletotrichum gloeosporioides. Fungal Genet Biol. 44, 32–43 (2007).
Ghibelli, L. & Diederich, M. Multistep and multitask Bax activation. Mitochondrion 10 (2010).
Lam, E., Kato, N. & Lawton, M. Programmed cell death, mitochondria and the plant hypersensitive response. Nature 411, 848–853 (2001).
Govrin, E. M. & Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr Biol. 10, 751–757 (2000).
Huang, W., Shang, Y., Chen, P., Gao, Q. & Wang, C. MrpacC regulates sporulation, insect cuticle penetration and immune evasion in Metarhizium robertsii. Environ. Microbiol., 17, 994–1008 (2015).
Wang, C., Duan, Z. & St Leger, R. J. MOS1 osmosensor of Metarhizium anisopliae is required for adaptation to insect host hemolymph. Eukaryot Cell 7, 302–309 (2008).
Larkin, M. A. et al. Clustal W and Clustal X version 2.0. Bioinformatics 23, 2947–2948, (2007).
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30, 2725–2729, (2013).
Wang, C. & St Leger, R. J. The Metarhizium anisopliae Perilipin Homolog MPL1 Regulates Lipid Metabolism, Appressorial Turgor Pressure and Virulence. J Biol. Chem. 282, 21110–21115 (2007).
Hockenbery, D. M., Oltvai, Z. N., Yin, X. M., Milliman, C. L. & Korsmeyer, S. J. Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell 75, 241–251 (1993).
Gao, Q., Shang, Y., Huang, W. & Wang, C. Glycerol-3-phosphate Acyltransferase contributes to triacylglycerol biosynthesis, lipid droplet formation and host invasion in Metarhizium robertsii. Appl. Environ. Microbiol. 79, 7646–7653 (2013).
Wang, C. & St Leger, R. J. A collagenous protective coat enables Metarhizium anisopliae to evade insect immune responses. Proc. Natl. Acad. Sci. USA 103, 6647–6652 (2006).
Duan, Z., Shang, Y., Gao, Q., Zheng, P. & Wang, C. A phosphoketolase Mpk1 of bacterial origin is adaptively required for full virulence in the insect-pathogenic fungus Metarhizium anisopliae. Environ. Microbiol. 11, 2351–2360 (2009).
Acknowledgements
This work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences (XDB11030100) and the National Nature Science Foundation of China (31225023).
Author information
Authors and Affiliations
Contributions
C.X., Z.D., P.C. and Y.S. performed the experiments. C.X. and Z.D. wrote the main manuscript text. C.W. designed the experiments and revised the manuscript. All authors reviewed the manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chen, Y., Duan, Z., Chen, P. et al. The Bax inhibitor MrBI-1 regulates heat tolerance, apoptotic-like cell death and virulence in Metarhizium robertsii. Sci Rep 5, 10625 (2015). https://doi.org/10.1038/srep10625
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep10625
- Springer Nature Limited
This article is cited by
-
The Bax inhibitor UvBI-1, a negative regulator of mycelial growth and conidiation, mediates stress response and is critical for pathogenicity of the rice false smut fungus Ustilaginoidea virens
Current Genetics (2019)
-
Wheat Bax Inhibitor-1 interacts with TaFKBP62 and mediates response to heat stress
BMC Plant Biology (2018)
-
The Ste12-like transcription factor MaSte12 is involved in pathogenicity by regulating the appressorium formation in the entomopathogenic fungus, Metarhizium acridum
Applied Microbiology and Biotechnology (2017)
-
The transcription factor SKN7 regulates conidiation, thermotolerance, apoptotic-like cell death and parasitism in the nematode endoparasitic fungus Hirsutella minnesotensis
Scientific Reports (2016)