
Abstract
Few studies have investigated the relative contribution of specific nutrients to momentary and season-long foraging responses by ants. Using western carpenter ants, Camponotus modoc, and European fire ants, Myrmica rubra, as model species, we: (1) tested preferential consumption of various macro- and micro-nutrients; (2) compared consumption of preferred macro-nutrients; (3) investigated seasonal shifts (late May to mid-September) in nutrient preferences; and (4) tested whether nutrient preferences of C. modoc and M. rubra pertain to black garden ants, Lasius niger, and thatching ants, Formica aserva. In laboratory and field experiments, we measured nutrient consumption by weighing Eppendorf tubes containing aqueous nutrient solutions before and after feeding by ants. Laboratory colonies of C. modoc favored nitrogenous urea and essential amino acids (EAAs), whereas M. rubra colonies favored sucrose. Field colonies of C. modoc and M. rubra preferentially consumed EAAs and sucrose, respectively, with no sustained shift in preferred macro-nutrient over the course of the foraging season. The presence of a less preferred macro-nutrient in a nutrient blend did not diminish the blend’s ‘appeal’ to foraging ants. Sucrose and EAAs singly and in combination were equally consumed by L. niger, whereas F. aserva preferred EAAs. Baits containing both sucrose and EAAs were consistently consumed by the ants studied in this project and should be considered for pest ant control.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Adequate nutrition is vital for colony fitness in ants1,2, affecting colony functioning, brood production and development, and worker survival2. The availability of carbohydrates as an energy source affects both the activity3,4,5,6 and the longevity of worker ants3,4,6,7,8,9,10, which, in turn, modulate foraging activities and aggressiveness of ant colonies3,4,5,6,11,12. Protein sources, in combination with carbohydrates, are essential for egg production by queens and brood development3,4,13,14,15,16, but in high amounts are toxic to workers7,8,9,10.
Ant colonies face challenges to meet their nutritional needs. Foragers must locate and recognize required nutrient resources, and integrate their own nutritional needs with those of their nestmates2. Ants locate food resources by responding to their odor plumes17,18,19,20,21,22,23,24, or by following trail pheromone deposited by forager ants25. The pattern of nutrient collection by ants may shift over time, with more carbohydrate- or protein-rich resources collected at different times of year26,27,28,29. Shifts in nutrient preference may be caused by demographic shifts in ant colonies or shifts in nutrient availability in the ants’ habitat. Experimentally increased amounts of colony brood mobilized foraging in L. niger workers30, prompted more food and more protein collection by Rhytidoponera ants31,32, and more protein consumption by Ochetellus glaber ants33. Ant colonies that have plenty of readily available nutrients preferentially seek foods containing scarcer nutrients34,35.
Ants assess food quality through the presence and concentration of certain macro-nutrients (proteins, carbohydrates, lipids) and micro-nutrients (e.g., salts, vitamins). In aphid honeydew, ants recognize the different types of sugar molecules36,37,38,39,40,41 (e.g., sucrose, melezitose) and generally they prefer resources with high sugar concentration39,41,42. Ants also require proteins, and obtain amino acids—the ‘building blocks’ of proteins—by e.g. (i) ingesting free amino acids from plant nectar43,44,45,46, (ii) digesting proteins from insect prey47, and (iii) by acquiring the amino acids that symbiotic gut microbes produce from nitrogenous waste15,48. Ants prefer essential to non-essential amino acids49, and recognize when specific amino acids are lacking49. Ants also feed on certain oils38, and recognize distinct fatty acids (e.g., oleic acid) and glycerides (e.g., 1,2-diolein) that are present on the surface of insect prey, deceased insects, and seed elaiosomes, and that serve as pick-up cues by ants50,51,52,53. Furthermore, ants recognize and consume micro-nutrients, including salts54,55,56,57 (e.g., NaCl) and some B-vitamins58.
Very few studies have comprehensively examined the ants’ preferences for specific macro- and micro-nutrients or for these nutrients as components of complex nutrient blends37,49,58. Although seasonal shifts in nutrient preference by ants have been demonstrated26,27,28,29, these nutrients were often presented as complex blends (e.g., tuna, fruit conserves) with sometimes different physical properties, making it difficult to attribute the ants’ preferential feeding responses to any one nutrient component.
Previously, we have determined nutrient preferences of two generalist ant species, the western carpenter ant, Camponotus modoc, and the European fire ant, Myrmica rubra41. Camponotus modoc inhabits temperate forests along the western coast of North America59 and excavates nests in the wood of conifer trees60. Myrmica rubra is an aggressive soil-dwelling ant that is native to Eurasia but has invaded the east and west coasts of North America61. While both species prefer sucrose to other saccharides41, and both species prefer essential to non-essential amino acids (AR, unpublished data), only C. modoc also consumes urea (AR, unpublished data). Lasius niger is a widespread62,63, temperate, soil-dwelling ant that regularly tends aphids and prefers aphid-derived sugars such as melezitose to common sugars such as sucrose36,64. Lasius niger also preferentially feeds on diverse amino acid blends64,65 but any potential preference for specific amino acids is not known. Formica aserva is a brood-raiding ant, nesting in woody debris66 such as stumps67. It tends aphids68 and collects insect prey69 but nutrient preferences are not yet documented. We selected these four species to represent ants in diverse taxa with contrasting morphology and body size, life history traits, and habitat preferences.
Here, we investigated momentary (ad hoc), and seasonal, nutrient preferences of C. modoc and M. rubra colonies, and determined whether their observed nutrient preferences apply to other ant taxa. Specifically, we: (1) tested nutrient consumption within groups of either macro-nutrients or micro-nutrients; (2) compared consumption of preferred macro-nutrients; (3) investigated potential seasonal shifts in nutrient preferences; and (4) tested whether nutrient preferences of C. modoc and M. rubra pertain to L. niger and F. aserva.
Materials and methods
Maintenance of laboratory ant colonies
We collected and maintained nine colonies of C. modoc between 2016 and 2020, as reported70. Briefly, we removed infested log sections from coniferous forests near Squamish (British Columbia), and transferred these sections to large plastic bins (64 × 79 × 117 cm) kept in an outdoor, under-cover area of the Science Research Annex at Simon Fraser University. All colonies experienced natural weather and light cycles which can be important for colony survival71. Bins were connected to glass containers (30.5 × 26 × 50.8 cm) which served as the ants’ foraging area. The upper inner bin and container walls were coated with an equal mix of Vaseline (Unilever, London, UK) and paraffin oil (Anachemia, Lachine, QC H8R1A3, CA) to prevent ant escape. Ants were provisioned with apples, deceased cockroaches, and 20% sugar water ad libitum. Containers and bins had mesh covered holes to allow air exchange.
We collected and reared invasive M. rubra similar to previous reports41,72, with some modifications. In the summer of 2021, 10 colonies were dug up with their nesting soil at Inter-river Park (North Vancouver, BC, CA), and temporarily placed in glass jars (1L). Colonies were then transferred to separate glass containers (26 × 21 × 40.6 cm), with mesh-covered holes in container lids, and upper inner container walls coated with Vaseline and paraffin oil. Colonies were maintained indoors at 25–30 °C under a natural daylength cycle, and were provisioned with food as described above. The soil surface of containers served as the ants’ foraging area. Every two weeks, water was added to the soil to ensure adequate moisture content. Colonies were kept indoors, instead of outdoors, to minimize the risk of ant escape on university campus.
Preparation of test stimuli
Prior to experiments, test stimuli were prepared by weighing nutrients (TR 204 scale; Denver Instrument Company, CO, USA; see Table 1 for number of test stimuli, and Table S1 for nutrient compositions) and mixing them into water, accounting for ~ 50% of the final volume. Once nutrients were dissolved, distilled water was added until the desired weight by volume solution (w/v) was reached. Aliquots (1 mL) of solutions were pipetted into 1.5-mL Eppendorf tubes (Thermo Fisher Scientific, Waltham, MA 02451, USA) and kept frozen until use in bioassays. For each experiment, we prepared as many tubes, including evaporation control tubes (see below), as required for testing stimuli consumption by all colonies. Sucrose and essential amino acids (EAAs) were selected as nutrients based on previous studies41, and EAAs were assembled drawing on both an article15 and personal communication with its senior author. Fatty acids (oleic, linoleic, linolenic) and glycerides (1,2 diolein, triolein) were tested because they serve as pickup cues for ants50,52,73. Selections of sterols, and their approximate ratio, were based on reported dietary needs of insects47. Salts and vitamins were tested at equal ratio drawing on the composition of a synthetic ant diet74 (see Table S1 for compositions of test stimuli).
General protocol for laboratory bioassays
On any experimental day, test stimuli were removed from the freezer, thawed, and vortexed, thus ensuring that all solutes were dissolved. Eppendorf tubes were then stuffed with a 1-cm-long piece of a cotton dental wick (Richmond Dental & Medical, Charlotte, NC 28205, USA) to allow nutrient consumption by ants without spillage (Fig. 1a). For each test stimulus, two tubes were prepared: one for ant consumption and another for tracking passive water evaporation during bioassays. All tubes were weighed just prior to, and at the end of, bioassays. In each bioassay replicate, we tested the consumption response of a different colony and prepared as many Eppendorf tubes as stimuli were tested (typically 4–5), with each tube containing a specific nutrient solution or a plain water control. Prior to the onset of a bioassay, tubes were weighed and uncapped, and then presented to the ants, allowing them to forage for 4–6 h (see below). Bioassay times for C. modoc and M. rubra colonies were set to 4 h and 6 h, respectively, accounting for differences in worker size60,61 and considering the time that was needed to obtain measurable consumption rates in preliminary experiments. Colonies for laboratory experiments were randomly selected on each experimental day, and were given at least 36 h between experiments. Each colony experienced a particular set of test stimuli only once. All laboratory experiments were conducted between June and early September.

Design of laboratory and field experiments for testing comparative food consumption by ants. (a) Photograph of an Eppendorf tube containing an aqueous nutrient solution retained by a piece of dental cotton wick, enabling ants to consume the liquid bait without spillage. (b) Bioassay container for carpenter ants, Camponotus modoc, fitted with 1.5-mL Eppendorf tubes containing aqueous nutrient solutions. Similar methodology was used for testing food consumption by European fire ants, Myrmica rubra41. Evaporation control tubes were placed on a small platform suspended from the container ceiling. (c) Eppendorf tubes affixed to a tree to test for preferential food consumption by C. modoc and black garden ants, Lasius niger. For field experiments with M. rubra and Formica aserva, Eppendorf tubes were placed on the ground.
Bioassays with C. modoc (and M. rubra below) were run mostly on warm and sunny days when colonies are most active (AR, pers. obs.). Prior to bioassays, colonies were deprived of cockroaches and apples for 24 h, and of sugar water for 4 h (the maximum time elapsed before ants attempted to chew out of their containers). Bioassays were run in plexiglass containers (50.5 × 30.5 × 33 cm) covered by lids with mesh holes to allow ventilation (Fig. 1b). Tubes were taped, with positions randomly assigned and spaced equidistantly in an arc, to the container bottom 22 cm away from the container entrance hole. Corresponding evaporation control tubes were taped to a plexiglass platform suspended from the container lid. Just prior to initiating a bioassay, all tubes were uncapped and each container was connected via Tygon® tubing (diam.: 2.54 cm) and barbed plumbing connectors (diam.: 2.54 cm) to a C. modoc housing bin, allowing ants to freely forage in a container. Bioassay replicates were run for 4 h but were terminated sooner if ants had completely consumed the test solution of any one tube. Bioassay containers were cleaned with hexane and ethanol (70%), and plumbing fixtures and Tygon® tubings were washed with soapy water.
For bioassays with M. rubra, nest containers were co-opted as bioassay containers, and colonies were food-deprived for 24 h to motivate foraging. For each replicate, Eppendorf tubes were taped, randomly assigned, to the edge of a jar lid (diam: 15 cm), and corresponding evaporation control tubes were taped, inaccessible to ants, to the underside of container lids. Bioassay replicates were initiated by uncapping all Eppendorf tubes, and placing jar lids with Eppendorf tubes on the soil surface inside bioassay containers. Bioassays were run for 6 h but were terminated sooner if ants had consumed the entire test solution in an Eppendorf tube. Between replicates, jar lids were washed with soapy water.
Protocol of field experiment
In preparation for field experiments, Eppendorf tubes with nutrient solutions for ant consumption, and evaporation control tubes, were thawed, weighed, and then transported to the field in a cooler. Tubes were spaced around the entrance of ant nests or next to ant foraging trails, with tube positions randomly assigned in each replicate. For C. modoc, Eppendorf tubes were affixed 5 cm apart to trees or logs housing a C. modoc nest (Fig. 1c). For L. niger, we located nests at the base of trees, and affixed tubes next to each other on the trunk of trees alongside the ants’ foraging trails. For M. rubra, tubes were placed 5 cm apart around the entrance of subterranean nests. For F. aserva, tubes were placed on top of tree stumps that contained an ant nest. For all field studies, evaporation control tubes were placed in Tupperware containers (15 × 9 × 10 cm) with a mesh-covered hole in the lid, and containers were set near ant nests. Replicates with colonies of M. rubra (n = 10; repeated on 7 dates), L. niger (n = 10), C. modoc (n = 13; repeated on 6 dates), and F. aserva (n = 10) were run for 4 h, 16 h, 24 h, and 24 h, respectively. Bioassays were run between 10:00–14:00 for M. rubra, 17:00–09:00 for L. niger, and between 11:00–11:00 for both C. modoc and F. aserva. Experimental time periods for each species were set according to time periods needed in preliminary studies to obtain measurable consumption rates. After replicates were terminated, tubes were capped, transported to the laboratory in a cooler, and weighed. All 4- to 24-h field studies were run on warm and sunny days with observable ant activity. Experiments with C. modoc and M. rubra were run from late May to mid-September, and experiments with L. niger and F. aserva were run in June and August, respectively.
Specific experiments
Assessing consumption of various macro- and micro-nutrients (Exps. 1–12; Lab)
In experiments 1–2 (Table 1), we offered colonies of C. modoc and M. rubra a choice between aqueous sucrose solutions at three concentrations (0.625%, 1.25%, and 2.5% w/v), and a water control.
Experiments 3–8 (Table 1) tested consumption of lipid-related nutrients by C. modoc and M. rubra colonies. Each of three lipid types (glycerides, fatty acids, sterols) consisted of 2–4 constituents (Table S1) which were formulated in an aqueous solution at two concentrations (Exps. 3–4: glycerides: 0.5%, 1.0% w/v; Exps. 5–6: fatty acids: 1.25%, 2.5% w/v; Exps. 7–8: sterols: 0.5%, 1.0% w/v), using Tween 80 as the emulsifier. In each experiment, both Tween 80 in water, and water, served as control stimuli.
Experiment 9–12 (Table 1) tested consumption of micro-nutrient salts or vitamins by C. modoc and M. rubra colonies. Each type of micro-nutrient consisted of 7–11 constituents (Table S1) which were dissolved in water at three concentrations (Exps. 9–10: salts: 0.25%, 0.5%, 1.0% w/v; Exps. 11–12: vitamins: 0.25%, 0.5%, 1.0% w/v), with water serving as the control stimulus in each experiment.
Comparing consumption of preferred macro-nutrients (Exps. 13–20; Lab)
Experiments 13–20 (Table 1) compared consumption of macro-nutrients that C. modoc or M. rubra colonies were previously shown to preferentially consume, including urea (AR, unpublished data), essential amino acids (EAAs; AR, unpublished data), and sucrose41. In experiments 13–16, aqueous solutions of urea, EAAs, and sucrose were tested singly and in ternary combination, with plain water as the control stimulus. Single components were tested at the same ‘unadjusted’ concentration as in the ternary blend (urea 2.5%; EAAs 0.55%; sucrose 2.5% w/v) or at an ‘adjusted’ concentration (5.55% w/v) that equalled the total concentration of the ternary blend (5.55%). Each component in the ternary blend was tested at the lowest concentration found effective in pre-screening experiments (see Result of Exps. 1–12; AR, unpublished data). In experiments 17–20 (Table 1), aqueous solutions of urea, EAAs, and sucrose were tested in all binary and ternary combinations, again with plain water as the control stimulus. Binary combinations were tested at the same ‘unadjusted’ concentration as in the ternary blend (urea [2.5%] + EAA [0.55%]; urea [2.5%] + sucrose [2.5%]; EAA [0.55%] + sucrose [2.5%]) or at an ‘adjusted’ concentration (urea [4.55%] + EAA [1.0%]; urea [2.775%] + sucrose [2.775%]; EAA [1.0%] + sucrose [4.55%]) that equalled the total concentration of the ternary blend (5.55%).
Evaluating potential seasonal shifts in nutrient consumption (Exps. 21–22; Field)
Experiments 21–22 (Table 1) investigated potential seasonal shifts in nutrient preferences exhibited by field colonies of ants. We worked with 13 colonies of C. modoc and 10 colonies of M. rubra located along the Mamquam forest service road (near Squamish, BC, Canada) and at Inter River Park (District of North Vancouver, BC, Canada), respectively. Drawing on results of preceding experiments that both C. modoc and M. rubra had preferentially consumed the ‘adjusted’ binary blend of EAA + sucrose (see “Results”), we offered each ant colony four Eppendorf tubes that contained: (1) EAA (5.55%); (2) sucrose (5.55%), (3) EAA (1.0%) + sucrose (4.55%); and (4) plain water (control). Throughout the summer season, we measured nutrient consumption by colonies in circa 3-week intervals on six dates for C. modoc colonies (18 June 2021 to 07 September 2021), and on seven dates for M. rubra colonies (21 May 2021 to 13 September 2021).
Investigating nutrient consumption of L. niger and F. aserva (Exps. 23–24; Field)
We worked with 10 field colonies each of L. niger and F. aserva that were located on the Burnaby campus of Simon Fraser University and along the Mamquam forest service road (see above), respectively. Each nest was offered four Eppendorf tubes that contained: (1) EAAs (5.55%); (2) sucrose (5.55%), (3) EAAs (1.0%) + sucrose (4.55%); and (4) plain water (control).
Statistical analyses
To calculate the amount of each nutrient solution that was consumed by a colony, we first determined the weight loss of the corresponding evaporation control solution, and then subtracted this value from the weight loss of the test solution. To account for differences in colony size and foraging activity between colonies, we analysed proportions, rather than absolute amounts, of nutrient solutions consumed. To obtain proportional consumption data for a colony in any experimental replicate, we divided the amount (weight) of each nutrient solution consumed by the total amount of all nutrient solutions consumed. When there had been little feeding activity by a colony, some consumption data became less than zero (~ −7 mg) following weight loss subtraction due to water evaporation measured in evaporation controls (see above). As evaporation control tubes were close to the arena vent (Fig. 1b), these small negative values could be due to slightly elevated rates of evaporation. As there could not be ‘negative feeding’ on a nutrient solution by a colony, we considered these less-than-zero values to be zero. We included replicates in data analyses when a colony had positive consumption responses for at least two of all the nutrient solutions that were tested in that replicate. Using a beta distributed generalized linear mixed model (GLMM), we applied a standard transformation to restrict our data between the bounded interval of 0 and 1. In experiments 1–20 and 23–24, we fit proportion consumed as our response variable and treatment as our predictor, with ant colony as a random intercept. For experiments 21–22, we fit proportion consumed as our response variable and treatment, date, and interaction between treatment and date as predictors, with ant colony as a random intercept. We evaluated the significance of predictors, using likelihood ratio tests and made Tukey adjusted pairwise comparisons between mean proportional consumption values between treatments.
Data75 were analysed and graphed using R (v. 4.2.2) and R studio (v. 2022.07.1 + 554)76. Data were processed using functions from the tidyverse77 and the plyr package78. GLMMs were fit using the glmmTMB package79, and model fit, residual normality, variance, and over/under dispersion patterns were inspected, using the DHARMa package80. We obtained estimated marginal means and 95% confidence intervals using the emmeans package81. We produced graphics using the ggplot2 package82 and completed figure assemblies in Inkscape (v. 1.0.2).
Results
Assessing consumption of various macro- and micro-nutrients (Exps. 1–12; Lab)
Concentrations of sucrose in aqueous solutions affected their consumption by C. modoc colonies (χ2 = 8.75, d.f. = 3, p = 0.03; Fig. S1a) and by M. rubra colonies (χ2 = 54.18, d.f. = 3, p < 0.0001; Fig. S1b). Colonies of C. modoc preferentially consumed the 1.5% (w/v) sucrose solution, which was the only solution consumed significantly more than water. Colonies of M. rubra consumed significantly more of the 2.5% (w/v) sucrose solution than of any other solution including the water control (Table S2).
Glycerides in emulsified aqueous solutions did not prompt consumption by C. modoc colonies (χ2 = 2.03, d.f. = 3, p = 0.57; Fig. S2a) and M. rubra colonies (χ2 = 0.72, d.f. = 3, p = 0.87; Fig. S2b, Table S3).
Fatty acids in emulsified aqueous solutions significantly affected feeding responses of C. modoc colonies (χ2 = 8.46, d.f. = 3, p = 0.04; Figure S2c) and M. rubra colonies (χ2 = 8.90, d.f. = 3, p = 0.03; Fig. S2d). Increasing concentrations of fatty acids lowered consumption, with the 2.5% solution being consumed the least (Table S3).
Sterols in aqueous solutions did not affect consumption by C. modoc colonies (χ2 = 2.47, d.f. = 3, p = 0.48; Fig. S2e) but did affect consumption by M. rubra colonies (χ2 = 9.90, d.f. = 3, p = 0.02; Fig. S2f). Colonies of M. rubra consumed more of the 0.5% sterol solution than of the 1% sterol solution and the water control but not of the tween + water control (Table S3). The 1% sterol solution, Tween + water, and water all prompted comparable consumption (Table S3).
Salts in aqueous solutions did not affect consumption by C. modoc colonies (χ2 = 0.89, d.f. = 3, p = 0.83; Fig. S3a) but was a significant predictor of proportional consumption by M. rubra colonies (χ2 = 32.26, d.f. = 3, p < 0.0001; Fig. S3b). Increasing salt concentrations lowered consumption, with the 1% solution being consumed the least (Table S3).
Vitamins in aqueous solutions did not affect consumption by C. modoc colonies (χ2 = 0.10 d.f. = 3, p = 0.99; Fig. S3c) but affected consumption by M. rubra colonies (χ2 = 20.66, d.f. = 3, p < 0.001; Fig. S3d). Colonies of M. rubra equally consumed the 1% vitamin solution and plain water, both of which more than the 0.25% and 0.5% solution (Table S4).
Comparing consumption of preferred macro-nutrients (Exps. 13–20; Lab)
Colonies of C. modoc and M. rubra differentially consumed 1- and 3-component aqueous solutions of urea, EAAs, and sucrose, and plain water (control stimulus) (Table 1) (C. modoc: unadjusted concentrations of nutrients in aqueous solutions: χ2 = 71.15, d.f. = 4, p < 0.0001, Fig. 2a; C. modoc: adjusted concentrations: χ2 = 57.72, d.f. = 4, p < 0.0001, Fig. 2b; M. rubra: unadjusted concentrations: χ2 = 61.59, d.f. = 4, p < 0.0001, Fig. 2c; M. rubra: adjusted concentrations: χ2 = 91.7, d.f. = 4, p < 0.0001, Fig. 2d). Camponotus modoc colonies preferentially consumed solutions containing urea, EAAs, or both (together with sucrose) (Fig. 2a,b). At adjusted nutrient concentrations, EAAs on their own and in ternary combination with urea and sucrose were most heavily consumed (Table S5). When nutrient concentrations were unadjusted, M. rubra colonies preferentially consumed sucrose, and sucrose in ternary combination with urea and EAAs. At adjusted nutrient concentrations, M. rubra colonies preferentially consumed single-nutrient solutions of EAAs and sucrose, followed by the ternary blend of EAAs, sucrose, and urea (Table S5).

Comparative consumption of liquid food baits by colonies of Camponotus modoc carpenter ants [a (n = 8), b (n = 9)], and Myrmica rubra fire ants [c (n = 9), d (n = 8)]. Colonies were offered a choice of aqueous solutions of macro-nutrients—urea, essential amino acids (EAAs), and sucrose—that were presented singly and in a ternary blend (‘Blend’). The concentration of single macro-nutrients was either not adjusted [a, c; urea (2.5%); EAAs (0.55%); sucrose (2.5%) w/v] or adjusted (b, d; urea, EAAs, and sucrose all 5.55%) to the same total concentration as the ternary blend [urea (2.5%), EAAs (0.55%), and sucrose (2.5%) w/v]. Coloured symbols indicate consumption rates of individual colonies (replicates) and black symbols represent modelled estimated marginal means and 95% confidence intervals. Means with different letters are statistically different (p < 0.05) in pairwise comparisons (see Table S5).
There was also differential consumption of macro-nutrients by C. modoc and M. rubra colonies when sucrose, EAAs, and urea were offered—at unadjusted and adjusted nutrient concentrations—in all possible binary and ternary combinations, along with plain water as the control stimulus (C. modoc: unadjusted concentrations: χ2 = 30.79, d.f. = 4, p < 0.0001, Fig. 3a; C. modoc: adjusted concentrations: χ2 = 30.71, d.f. = 4, p < 0.0001, Fig. 3b; M. rubra: unadjusted concentrations: χ2 = 61.329, d.f. = 4, p < 0.0001, Fig. 3c; M. rubra: adjusted concentrations: χ2 = 115.38, d.f. = 4, p < 0.0001, Fig. 3d). At unadjusted nutrient concentrations, C. modoc colonies equally consumed all binary and ternary nutrient blends, significantly preferring all of them to plain water (Fig. 3a; Table S6). At adjusted nutrient concentrations, C. modoc colonies consumed the blend of urea + EAAs significantly more than the blend of urea + sucrose, and water, but not significantly more than the blend of EAAs + sucrose, and the ternary blend (Fig. 3b; Table S6). At unadjusted nutrient concentrations, M. rubra colonies preferentially consumed the blend of EAAs + sucrose and the ternary blend, followed by blends of urea + sucrose and urea + EAAs, with the latter blend being consumed as little as water (Fig. 3c; Table S6). At adjusted nutrient concentrations, the blend of EAAs + sucrose was most heavily consumed, followed by the ternary blend, and by binary blends of urea + sucrose and urea + EAAs, which had similar levels of consumption, both significantly higher than water (Fig. 3d; Table S6).

Comparative consumption of liquid food baits by colonies of Camponotus modoc carpenter ants [a (n = 9), b (n = 9)] and Myrmica rubra fire ants [c (n = 9), d (n = 9)]. Colonies were offered a choice of aqueous solutions of macro-nutrients—urea, essential amino acids (EAAs), and sucrose—that were tested in binary combinations and in a ternary blend (‘Blend’). The concentration of binary combinations was either not adjusted [a, c; urea (2.5%) and EAAs (0.55%); urea (2.5%) and sucrose (2.5%); EAAs (0.55%) and sucrose (2.5%)] or adjusted [b, d; urea (4.55%) and EAAs (1.0%); urea (2.775%) and sucrose (2.775%); EAAs (1.0%) and sucrose: 4.55%) to the same total concentration as the ternary blend [urea (2.5%), EAAs (0.55%), and sucrose (2.5%) w/v)]. Coloured symbols indicate responses of individual colonies (replicates) and black symbols represent modelled estimated marginal means and 95% confidence intervals. Means with different letters are statistically different (p < 0.05) in pairwise comparisons (see Table S6).
Evaluating potential seasonal shifts in nutrient consumption (Exps. 21–22; Field)
In the field experiment with C. modoc colonies, bait nutrient(s), date, and interaction between bait nutrient(s) and date, were all significant predictors of bait consumption by ants (bait nutrient(s): χ2 = 371.1, d.f. = 18, p < 0.0001; date: χ2 = 117.04, d.f. = 20, p < 0.0001; interaction between bait nutrient(s) and date: χ2 = 109.95, d.f. = 15, p < 0.0001; Fig. 4a). Invariably over time, C. modoc colonies preferentially consumed EAAs, and EAAs + sucrose in a binary blend (Fig. 4a; Table S7). Consumption of water, and of sugar, decreased over time75 (Fig. 4a).

Comparative consumption of liquid food baits by field colonies of Camponotus modoc carpenter ants (n = 12–13) (a), and Myrmica rubra fire ants (n = 10) (b) during 21 May to 13 September 2021. Colonies were offered a choice between aqueous solutions of essential amino acids [EAAs (5.55%), w/v), sucrose (5.55%, w/v), and both [EAAs (1%), sucrose (4.55%), w/v]. Coloured symbols indicate the responses of individual colonies, and black symbols represent modelled estimated marginal means and 95% confidence intervals. Nutrient solution, date, and interaction between nutrient solution and date, were all significant predictors of bait consumption (see “Results”). Statistical results of pairwise comparisons within date are reported in Table S7.
Similarly, in the field experiment with M. rubra colonies, bait nutrient(s), date, and interaction between bait nutrient(s) and date, were all significant predictors of bait consumption by ants [bait nutrient(s): χ2 = 403.59, d.f. = 21, p < 0.0001; date: χ2 = 267.51, d.f. = 24, p < 0.0001; interaction between bait nutrient(s) and date: χ2 = 245.17, d.f. = 18, p < 0.0001; Fig. 4b]. Across sampling dates, M. rubra colonies generally consumed more sucrose, and more sucrose + EAAs in a binary blend, than EAAs and water (Fig. 4b; Table S7). Sucrose consumption declined over time, whereas the consumption of sucrose in a binary blend with EAAs increased during the last three sampling dates75 (Fig. 4b).
Investigating nutrient consumption of L. niger and F. aserva (Exps. 23–24; Field)
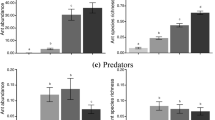
Bait nutrients affected bait consumption by L. niger colonies (χ2 = 48.33, d.f. = 3, p < 0.0001; Fig. 5a) and F. aserva colonies (χ2 = 12.29, d.f. = 3, p = 0.006; Fig. 5b). Colonies of L. niger equally consumed baits containing EAAs, sucrose, and EAAs + sucrose, all of which being preferred to plain water (control stimulus) (Table S7). Colonies of F. aserva preferentially consumed baits containing EAAs, which they consumed more than sucrose baits but (statistically) not more than EAA + sucrose baits (Table S7).

Comparative consumption of liquid food baits by colonies of Lasius niger black garden ants (n = 10) and Formica aserva thatching ants (n = 10). Colonies were offered a choice between aqueous solutions of essential amino acids [EAAs (5.55%) w/v], sucrose (5.55%, w/v), and both [EAAs (1%); sucrose (4.55%) w/v). Coloured symbols represent responses of individual colonies and black symbols are estimated marginal means and 95% confidence intervals. Treatment (bait composition) was a significant predictor of bait consumption (see results). Means with different letters are statistically different (p < 0.05) in pairwise comparisons (see Table S8).
Discussion
Macronutrient preferences differed among the ant taxa we tested in our study which varied in size, life history, and preferred habitat. Camponotus modoc and M. rubra fed on macro-nutrients but not on micro-nutrients, and preferentially consumed specific macro-nutrients such as essential amino acids (EAAs) and sucrose. Each species, however, preferred a different macro-nutrient. Camponotus modoc favored nitrogenous urea and EAAs, whereas M. rubra favored sucrose. In contrast, neither species consumed micro-nutrients such as salts and vitamins, and lipid-related compounds such as glycerides and fatty acids. There was no shift in preferred macro-nutrient(s) over the course of the foraging season. Colonies of C. modoc preferentially and consistently consumed EAAs, and EEAs blended with sucrose, whereas M. rubra colonies generally consumed sucrose, and sucrose blended with EAAs. Macro-nutrients preferentially consumed by C. modoc and M. rubra were also readily consumed by L. niger and F. aserva. We did not observe other ant species at baits during field trials, indicating that all data were generated exclusively by our study species. Although macro-nutrient preference and consumption differed among species, the underlying drivers for these differences, such as contrasting life-history and spatiotemporal food availability, are yet to be studied.
Various species of ants distinguish between different saccharides37,38,39,40,41, and between different amino acids49. To date, only Camponotus ants have been demonstrated to consume urea (this study;83,84,85). Lipid-related compounds such as glycerides and sterols were as unappealing as water controls, and fatty acids were ingested even less than water. These results were not expected considering that ants use glycerides and fatty acids on the surface of food items as pickup cues50,51,52,53. Additionally, ants ingest oils38, although it is not known how they distinguish between oil types. The propensity of ants to salt-feed increases with distance from the ocean54, and is generally common in arboreal and herbivorous species54,57 but see56. However, C. modoc did not consume salt solutions and M. rubra was deterred by them. This may have been due, in part, to the salt composition we tested for consumption by ants. We offered a blend of salts drawing on the composition of a synthetic ant diet74, whereas other studies offered sodium chloride (NaCl) as a single salt. Vitamins did not elicit foraging responses in our study but B vitamin added to water improved its acceptance by the imported fire ant Solenopsis richerti, albeit to a lesser extent than sugars or amino acids58. The effects of certain vitamins on ant colony health remain inconsistent14,86.
Foraging preferences by ants in our study were largely driven by the specific macro-nutrient that each ant favored. Colonies of C. modoc and F. aserva preferentially foraged on the nitrogenous macro-nutrients, urea and/or EAAs, whereas M. rubra colonies preferentially consumed sucrose. Based on mean feeding responses, field colonies of L. niger consumed sucrose and EEAs equally. However, feeding preferences of individual colonies observably differed, with some colonies favoring sucrose and others EAAs75. These data indicate that the colonies’ preferred macro-nutrient may shift over time in accordance with the colonies’ demographics and/or resource availability or competition in their habitat. Similarly, bait selectivity by tropical ants was affected by prior feeding experience and by competition with ant community members at bait stations37. In contrast, field colonies of C. modoc and M. rubra consistently favored EAAs and sucrose, respectively, demonstrating persistent selection of a specific macro-nutrient. Similarly, tropical ants species preferred specific, and contrasting, blends of sucrose and particular amino acids37. Geometric framework studies (investigating the effects of nutrient mixtures on ant health) concluded that ants prioritise sustained carbohydrate supplies2 which are deemed essential for colony health3,4,6,7,8,9,10, whereas field studies revealed that many ant species prefer blends of sucrose and amino acids to sucrose alone37. In our season-long (21 May to 13 September) field study with C. modoc and M. rubra (Fig. 4), the blend of sucrose + EAAs was consistently consumed at a level comparable to consumption levels of EAAs or sucrose alone. Also, there was no sustained temporal shift in preferred macro-nutrient or blend of macro-nutrients, contrasting with previous reports that ants selectively seek carbohydrates or proteins at certain times during the foraging season26,27,28,29.
Both intrinsic and extrinsic factors affect foraging preferences in ants. As an intrinsic factor, the presence of brood motivates food and protein collections31,32,33. However, this intrinsic factor did not seem to have affected seasonal foraging patterns of C. modoc and M. rubra colonies (Fig. 4). Colonies of both species have seasonal egg production peaks, but larvae are consistently present in nests60,87, suggesting that fluctuations in the number of larvae may be too subtle to affect seasonal foraging activities. As an extrinsic factor, resource availability in time and space affects foraging patterns of ants, with scarce nutrients most intensely sought34,35. Seasonally, yellow crazy ants, Anoplolepis gracilipes, preferentially seek sugar-rich food in the wet season and protein-rich food in the dry season, despite brood being present in nests year round27. These preferences align with the shortages of protein-rich invertebrates during the dry season and sugar-rich honeydew from scale insects during the wet season27. Nutrient preferences may also differ according to the strata occupied by ants in an ecosystem28,88,89,90. For example, across six tropical biomes in South America, arboreal ants foraged most intensely on carbohydrates, whereas ground-nesting ants preferentially foraged on lipids88. Consistent preference by C. modoc for EAAs, and by M. rubra for sucrose, could imply that these resources are consistently limited and therefore are preferentially sought. That M. rubra equally consumed EAAs and sucrose in a laboratory experiment (Fig. 2d) but favoured sucrose, and sucrose blended with EEAs, in the season-long field experiment (Fig. 4b) could have been due to contrasting nutrients obtainable by laboratory and field colonies. In ants, both intrinsic and extrinsic factors are likely at play, simultaneously, although these interactions have not yet been explored.
Lastly, findings in our study have significant implications for control of (invasive) pest ants. Presently, leading commercial baits appeal to ‘sweet-loving ants’ but not to species that preferentially seek protein-rich food, and thus would require baits containing essential amino acids. In our study, colonies of C. modoc that preferentially ingested aqueous solutions of EAAs, and colonies of M. rubra that preferentially ingested aqueous solutions of sucrose, all consumed baits containing both EAAs and sucrose to the same extent as they consumed baits containing only their preferred macro-nutrient, indicating that the presence of a less preferred macro-nutrient as a bait constituent did not diminish the bait’s ‘appeal’. The same conclusion applies to the other ant taxa tested in this study, L. niger and F. aserva. It follows that both sucrose and EAAs could be constituents in the same bait, and thus would be appealing to both ‘sugar- and protein-loving ants’. Moreover, even if there were pest ants that shift their macro-nutrient preference over the foraging season from carbohydrates to proteins, or vice versa, both macro-nutrients would be present in the bait, thus retaining its season-long appeal to foraging ants. We favor boric acid as the lethal constituent in such a bait because—like sucrose and EAAs—it is water-soluble and once dissolved in water expresses antimicrobial activity91,92, thus preventing spoilage of the bait’s macro-nutrients.
Data availability
Data are available from Mendeley Data and can be accessed at: https://doi.org/10.17632/zbtvkyzwcj.175.
References
Feldhaar, H. Ant nutritional ecology: Linking the nutritional niche plasticity on individual and colony-level to community ecology. Curr. Opin. Insect Sci. 5, 25–30 (2014).
Csata, E. & Dussutour, A. Nutrient regulation in ants (Hymenoptera: Formicidae): A review. Myrmecol. News 29, 111–124 (2019).
Grover, C. D., Kay, A. D., Monson, J. A., Marsh, T. C. & Holway, D. A. Linking nutrition and behavioural dominance: Carbohydrate scarcity limits aggression and activity in Argentine ants. Proc. R. Soc. B Biol. Sci. 274, 2951–2957 (2007).
Shik, J. Z. & Silverman, J. Towards a nutritional ecology of invasive establishment: Aphid mutualists provide better fuel for incipient Argentine ant colonies than insect prey. Biol. Invasions 15, 829–836 (2013).
Kaspari, M., Donoso, D., Lucas, J. A., Zumbusch, T. & Kay, A. D. Using nutritional ecology to predict community structure: A field test in Neotropical ants. Ecosphere 3, 93 (2012).
Wittman, S. E., O’Dowd, D. J. & Green, P. T. Carbohydrate supply drives colony size, aggression, and impacts of an invasive ant. Ecosphere 9, e02403 (2018).
Cook, S. C., Eubanks, M. D., Gold, R. E. & Behmer, S. T. Colony-level macronutrient regulation in ants: Mechanisms, hoarding and associated costs. Anim. Behav. 79, 429–437 (2010).
Bazazi, S., Arganda, S., Moreau, M., Jeanson, R. & Dussutour, A. Responses to nutritional challenges in ant colonies. Anim. Behav. 111, 235–249 (2016).
Arganda, S. et al. Parsing the life-shortening effects of dietary protein: Effects of individual amino acids. Proc. R. Soc. B Biol. Sci. 284, 16–19 (2017).
Dussutour, A. & Simpson, S. Ant workers die young and colonies collapse when fed a high-protein diet. Proc. R. Soc. B Biol. Sci. 279, 2402–2408 (2012).
Kay, A., Taylor, Z., Heinen, J. L., Marsh, T. G. & Holway, D. A. Nutrition and interference competition have interactive effects on the behavior and performance of Argentine ants. Ecology 91, 57–64 (2010).
Kay, A., Shik, J. Z., Van Alst, A., Miller, K. A. & Kaspari, M. Diet composition does not affect ant colony tempo. Funct. Ecol. 26, 317–323 (2012).
Porter, S. D. Effects of diet on the growth of laboratory fire ant colonies (Hymenoptera: Formicidae). J. Kansas Entomol. Soc. 62, 288–291 (1989).
Mankowski, M. E. & Morrell, J. J. Effects of B vitamin deletion in chemically defined diets on brood development in Camponotus vicinus (Hymenoptera: Formicidae). J. Econ. Entomol. 107, 1299–1306 (2014).
Feldhaar, H. et al. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 5, 1–11 (2007).
Evans, J. D. & Pierce, N. E. Effects of diet quality and queen number on growth in leptothoracine ant colonies (Hymenoptera: Formicidae). J. New York Entomol. Soc. 103, 91–99 (1995).
Buehlmann, C., Graham, P., Hansson, B. S. & Knaden, M. Desert ants locate food by combining high sensitivity to food odors with extensive crosswind runs. Curr. Biol. 24, 960–964 (2014).
Schatz, B., Anstett, M. C., Out, W. & Hossaert-McKey, M. Olfactive detection of fig wasps as prey by the ant Crematogaster scutellaris (Formicidae; Myrmicinae). Naturwissenschaften 90, 456–459 (2003).
Yusuf, A. A., Crewe, R. M. & Pirk, C. W. W. Olfactory detection of prey by the termite-raiding ant Pachycondyla analis. J. Insect Sci. 14, 1–10 (2014).
Renyard, A. et al. Floral and bird excreta semiochemicals attract western carpenter ants. Front. Ecol. Evol. 10, 923871 (2022).
Wanjiku, C., Khamis, F. M., Teal, P. E. A. & Torto, B. Plant volatiles influence the African weaver ant-cashew tree mutualism. J. Chem. Ecol. 40, 1167–1175 (2014).
De Vega, C., Herrera, C. M. & Dötterl, S. Floral volatiles play a key role in specialized ant pollination. Perspect. Plant Ecol. Evol. Syst. 16, 32–42 (2014).
Schiestl, F. P. & Glaser, F. Specific ant-pollination in an alpine orchid and the role of floral scent in attracting pollinating ants. Alp. Bot. 122, 1–9 (2012).
Fischer, C. Y. et al. Bacteria may enhance species association in an ant–aphid mutualistic relationship. Chemoecology 25, 1–10 (2015).
Czaczkes, T. J., Grüter, C. & Ratnieks, F. L. W. Trail pheromones: An integrative view of their role in social insect colony organization. Annu. Rev. Entomol. 60, 581–599 (2015).
Cook, S. C., Eubanks, M. D., Gold, R. E. & Behmer, S. T. Seasonality directs contrasting food collection behavior and nutrient regulation strategies in ants. PLoS ONE 6, 25407 (2011).
Abbott, K. L., Green, P. T. & O’Dowd, D. J. Seasonal shifts in macronutrient preferences in supercolonies of the invasive Yellow Crazy Ant Anoplolepis gracilipes (Smith, 1857) (Hymenoptera: Formicidae) on Christmas Island, Indian Ocean. Austral Entomol. 53, 337–346 (2014).
Hahn, D. A. & Wheeler, D. E. Seasonal foraging activity and bait preferences of ants on Barro Colorado Island, Panama. Biotropica 34, 348–356 (2002).
Rust, M. K., Reierson, D. A., Paine, E. & Blum, L. J. Seasonal activity and bait preferences of the Argentine ant (Hymenoptera: Formicidae). J. Agric. Urban Entomol. 17, 201–212 (2000).
Portha, S., Deneubourg, J. L. & Detrain, C. Self-organized asymmetries in ant foraging: A functional response to food type and colony needs. Behav. Ecol. 13, 776–781 (2002).
Dussutour, A. & Simpson, S. J. Carbohydrate regulation in relation to colony growth in ants. J. Exp. Biol. 211, 2224–2232 (2008).
Dussutour, A. & Simpson, S. J. Communal nutrition in ants. Curr. Biol. 19, 740–744 (2009).
Cornelius, M. L. & Grace, J. K. Influence of brood on the nutritional preferences of the tropical ant species, Pheidole megacephala (F.) and Ochetellus glaber (Mayr). J. Entomol. Sci. 32, 421–429 (1997).
Kay, A. The relative availabilities of complementary resources affect the feeding preferences of ant colonies. Behav. Ecol. 15, 63–70 (2004).
Kay, A. Applying optimal foraging theory to assess nutrient availability ratios for ants. Ecology 83, 1935–1944 (2002).
Völkl, W., Woodring, J., Fischer, M., Lorenz, M. W. & Hoffmann, K. H. Ant-aphid mutualisms: The impact of honeydew production and honeydew sugar composition on ant preferences. Oecologia 118, 483–491 (1999).
Blüthgen, N. & Fiedler, K. Preferences for sugars and amino acids and their conditionality in a diverse nectar-feeding ant community. J. Anim. Ecol. 73, 155–166 (2004).
Cornelius, M. L., Grace, J. K. & Yates, J. R. Acceptability of different sugars and oils to three tropical ant species (Hymen., Formicidae). Anz. Schädlingskde. Pflanzenschutz Umweltschutz 69, 41–43 (1996).
Zhou, A. M., Kuang, B. Q., Gao, Y. R. & Liang, G. W. Sucrose triggers honeydew preference in the ghost ant, Tapinoma melanocephalum (Hymenoptera: Formicidae). Florida Entomol. 98, 1217–1222 (2015).
Madsen, N. E. L. & Offenberg, J. Seasonal changes in sugar and amino acid preference in red wood ants of the Formica rufa group. Sociobiology 67, 144–152 (2020).
Renyard, A. et al. All sugars ain’t sweet: Selection of particular mono-, di- and trisaccharides by western carpenter ants and European fire ants. R. Soc. Open Sci. 8, 210804 (2021).
Sola, F. J. & Josens, R. Feeding behavior and social interactions of the Argentine ant Linepithema humile change with sucrose concentration. Bull. Entomol. Res. 106, 522–529 (2016).
Lanza, J., Vargo, E. L., Pulim, S. & Chang, Y. Z. Preferences of the fire ants Solenopsis invicta and S. geminata (Hymenoptera, Formicidae) for amino acid and sugar components of extrafloral nectars. Environ. Entomol. 22, 411–417 (1993).
Lanza, J. Response of fire ants (Formicidae: Solenopsis invicta and S. geminata) to artificial nectars with amino acids. Ecol. Entomol. 16, 203–210 (1991).
Blüthgen, N. & Fiedler, K. Competition for composition: Lessons from nectar-feeding ant communities. Ecology 85, 1479–1485 (2004).
González-Teuber, M. & Heil, M. The role of extrafloral nectar amino acids for the preferences of facultative and obligate ant mutualists. J. Chem. Ecol. 35, 459–468 (2009).
Chapman, R. F. The Insects: Structure and Function (Cambridge University Press, 2013).
Hu, Y. et al. Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome. Nat. Commun. 9, 964 (2018).
Csata, E. et al. Ant foragers compensate for the nutritional deficiencies in the colony. Curr. Biol. 30, 135–142 (2020).
Brew, C. R., O’Dowd, D. J. & Rae, I. D. Seed dispersal by ants: Behaviour-releasing compounds in elaiosomes. Oecologia 80, 490–497 (1989).
Marshall, D. L., Beattie, A. J. & Bollenbacher, W. E. Evidence for diglycerides as attractants in an ant-seed interaction. J. Chem. Ecol. 5, 335–344 (1979).
Hughes, L., Westoby, M. & Jurado, E. Convergence of elaiosomes and insect prey: Evidence from ant foraging behaviour and fatty acid composition. Funct. Ecol. 8, 358–365 (1994).
Pfeiffer, M., Huttenlocher, H. & Ayasse, M. Myrmecochorous plants use chemical mimicry to cheat seed-dispersing ants. Funct. Ecol. 24, 545–555 (2010).
Kaspari, M., Yanoviak, S. P. & Dudley, R. On the biogeography of salt limitation: A study of ant communities. Proc. Natl. Acad. Sci. USA. 105, 17848–17851 (2008).
Chavarria Pizarro, L., McCreery, H. F., Lawson, S. P., Winston, M. E. & O’Donnell, S. Sodium-specific foraging by leafcutter ant workers (Atta cephalotes, Hymenoptera: Formicidae). Ecol. Entomol. 37, 435–438 (2012).
Hernández, L. M. A., Todd, E. V., Miller, G. A. & Frederickson, M. E. Salt intake in amazonian ants: Too much of a good thing?. Insect. Soc. 59, 425–432 (2012).
Vieira, J. & Vasconcelos, H. L. Inter-generic and inter-habitat variation in the demand for sodium by Neotropical ants. Insect. Soc. 62, 133–140 (2015).
Ricks, B. L. & Vinson, B. S. Feeding acceptability of certain insects and various water-soluble compounds to two varieties of the imported fire ant. J. Econ. Entomol. 63, 145–148 (1970).
Hansen, L. D. & Klotz, J. H. Carpenter Ants of the United States and Canada (Cornell University Press, 2005).
Hansen, L. D. & Akre, R. D. Biology of carpenter ants in Washington state. Melanderia 43, 1–65 (1985).
Wetterer, J. K. & Radchenko, A. G. Worldwide spread of the ruby ant, Myrmica rubra (Hymenoptera: Formicidae). Myrmecol. News 14, 87–96 (2010).
Guénard, B., Weiser, M. D., Gómez, K., Narula, N. & Economo, E. P. The Global Ant Biodiversity Informatics (GABI) database: Synthesizing data on the geographic distribution of ant species (Hymenoptera: Formicidae). Myrmecol. News 24, 83–89 (2017).
Janicki, J., Narula, N., Ziegler, M., Guénard, B. & Economo, E. P. Visualizing and interacting with large-volume biodiversity data using client-server web-mapping applications: The design and implementation of antmaps.org. Ecol. Inform. 32, 185–193 (2016).
Woodring, J., Wiedemann, R., Fischer, M. K., Hoffmann, K. H. & Völkl, W. Honeydew amino acids in relation to sugars and their role in the establishment of ant-attendance hierarchy in eight species of aphids feeding on tansy (Tanacetum vulgare). Physiol. Entomol. 29, 311–319 (2004).
Madsen, N. E. L., Sørensen, P. B. & Offenberg, J. Sugar and amino acid preference in the black garden ant Lasius niger (L.). J. Insect Physiol. 100, 140–145 (2017).
Higgins, R. J., Gillingham, M. G. & Lindgren, B. S. Critical habitat elements, with an emphasis on coarse woody debris, associated with ant presence or absence in the moist cold sub-boreal forests of the interior of British Columbia. Forests 8, 129 (2017).
Wu, J. & Wong, H. R. Colonization of lodgepole pine stumps by ants (Hymenoptera: Formicidae). Can. Entomol. 119, 397–398 (1987).
Phillips, I. D. & Willis, C. K. R. Defensive behavior of ants in a mutualistic relationship with aphids. Behav. Ecol. Sociobiol. 59, 321–325 (2005).
Savolainen, R. & Deslippe, R. Facultative and obligate slave making in Formica ants. Naturwissenschaften 88, 347–350 (2001).
Renyard, A., Alamsetti, S. K., Gries, R., Munoz, A. & Gries, G. Identification of the trail pheromone of the carpenter ant Camponotus modoc. J. Chem. Ecol. 45, 901–913 (2019).
Sorvari, J., Haatanen, M. K. & Vesterlund, S. R. Combined effects of overwintering temperature and habitat degradation on the survival of boreal wood ant. J. Insect Conserv. 15, 727–731 (2011).
Hoefele, D., Chalissery, J. M., Renyard, A. & Gries, G. Experimentally guided development of a food bait for European fire ants. Entomol. Exp. Appl. 169, 780–791 (2021).
Bigley, W. S. & Vinson, S. B. Characterization of a brood pheromone isolated from the sexual brood of the imported fire ant, Solenopsis invicta. Ann. Entomol. Soc. Am. 68, 301–304 (1975).
Straka, J. & Feldhaar, H. Development of a chemically defined diet for ants. Insect. Soc. 54, 100–104 (2007).
Renyard, A., Gooding, C., Chalissery, J., Petrov, J. & Gries, G. R-code and data from: Effects of macro- and micro-nutrients on momentary and season-long feeding responses by select species of ants V1. Mendeley Data https://doi.org/10.17632/zbtvkyzwcj.1 (2024).
R Core Team. R: A Language and Environment for Statistical Computing. (2022).
Wickham, H. et al. Welcome to the tidyverse. J. Open Source Softw. 4, 1686 (2019).
Wickham, H. The split-apply-combine strategy for data analysis. J. Stat. Softw. 40, 1–29 (2011).
Brooks, M. E. et al. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400 (2019).
Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. (2022).
Lenth, R. Estimated Marginal Means, Aka Least-Squares Means. (2023).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2016).
Menzel, F., Staab, M., Chung, A. Y. C., Gebauer, G. & Blüthgen, N. Trophic ecology of parabiotic ants: Do the partners have similar food niches?. Austral Ecol. 37, 537–546 (2012).
Petit, S., Stonor, M. B., Weyland, J. J., Gibbs, J. & Amato, B. Camponotus ants mine sand for vertebrate urine to extract nitrogen. Austral Ecol. 45, 168–176 (2020).
Shetty, P. S. Gustatory preferences of ants (Camponotus compressus) for urea and sugars. Experientia 38, 259–260 (1982).
Poissonnier, L. A., Simpson, S. J. & Dussutour, A. Observations of the ‘“Egg White Injury”’ in ants. PLoS ONE 9, 1–10 (2014).
Kipyatkov, V. E. & Lopatina, E. B. Experimental study of seasonal cycle of rapid brood production in the ants Myrmica rubra L. and M. ruginodis Nyl. from two different latitudes. in Proceedings of the International Colloquia on Social Insects (ed. V. E. Kipyatkov) vols 3–4 195–206 (1997).
Lasmar, C. J. et al. Testing the context dependence of ant nutrient preference across habitat strata and trophic levels in Neotropical biomes. Ecology 104, 1–12 (2023).
Fowler, D., Lessard, J. P. & Sanders, N. J. Niche filtering rather than partitioning shapes the structure of temperate forest ant communities. J. Anim. Ecol. 83, 943–952 (2014).
Yanoviak, S. P. & Kaspari, M. Community structure and the habitat templet: Ants in the tropical forest canopy and litter. Oikos 89, 259–266 (2000).
Hernandez-Patlan, D. et al. Evaluation of the antimicrobial and intestinal integrity properties of boric acid in broiler chickens infected with Salmonella enteritidis: Proof of concept. Res. Vet. Sci. 123, 7–13 (2019).
Güzel, Y. et al. The efficacy of boric acid used to treat experimental osteomyelitis caused by methicillin-resistant Staphylococcus aureus: An in vivo study. Biol. Trace Elem. Res. 173, 384–389 (2016).
Acknowledgements
This research was supported by a Graduate Fellowship from Simon Fraser University and an Alexander Graham Bell CGSM scholarship from the Natural Sciences and Engineering Research Council of Canada (NSERC) to AR, an NSERC–Undergraduate Student Research Award (USRA) each to CG and JP. This research was further supported by an NSERC–Industrial Research Chair to GG, with BASF Canada Inc. and Scotts Canada Ltd. as the industrial sponsors. We thank two anonymous reviewers for meticulous and insightful reviews of the manuscripts, Michael Gudmundson for field assistance in locating and collecting carpenter ant nests, Grady Ott for generously donating plastic bins for housing carpenter ants, Erika Nassichuk for permission to work in Inter River Park, Regine Gries for technical support, Adam Blake for statistics and graphics advice, colleagues at the StatsBeerz group, particularly Hannah Watkins, for advice on data analyses, Robert Higgins for assistance in ant identification, Jenelle Breen, Kennedy Hoven, Kenza Zobaidi, Ranjot Bhandal, April Lin, and Jasper Li for help with ant care, and Sharon Oliver for comments.
Author information
Authors and Affiliations
Contributions
A.R., C.G., and G.G.: conceptualization. A.R., C.G., and G.G.: experimental design. A.R., C.G., J.C., and J.P.: data collection. AR: statistical analysis and graphics. A.R. and G.G.: writing—original draft preparation. A.R., G.G., C.G., J.C., and J.P.: writing—review and editing. All authors contributed to the article and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Renyard, A., Gooding, C., Chalissery, J.M. et al. Effects of macro- and micro-nutrients on momentary and season-long feeding responses by select species of ants. Sci Rep 14, 5727 (2024). https://doi.org/10.1038/s41598-024-56133-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-56133-y
- Springer Nature Limited
Keywords
This article is cited by
-
New lethal liquid bait for control of pest ants
Journal of Pest Science (2024)