
Abstract
The bacteriocins bactofencin A (class IId) and pediocin PA-1 (class IIa) are encoded by operons with a similarly clustered gene organization including a structural peptide, an immunity protein, an ABC transporter and accessory bacteriocin transporter protein. Cloning of these operons in E. coli TunerTM (DE3) on a pETcoco-2 derived vector resulted in successful secretion of both bacteriocins. A corresponding approach, involving the construction of vectors containing different combinations of these genes, revealed that the structural and the transporter genes alone are sufficient to permit heterologous production and secretion in this host. Even though the accessory protein, usually associated with optimal disulfide bond formation, was not required for bacteriocin synthesis, its presence did result in greater pediocin PA-1 production. The simplicity of the system and the fact that the associated bacteriocins could be recovered from the extracellular medium provides an opportunity to facilitate protein engineering and the overproduction of biologically-active bacteriocins at industrial scale. Additionally, this system could enable the characterization of new bacteriocin operons where genetic tools are not available for the native producers.
Similar content being viewed by others
Introduction
Antimicrobial peptides (AMPs) are produced by numerous organisms including many Gram-positive bacteria as a part of their innate defenses1, 2. AMPs secreted by Lactococcus, Lactobacillus, Pediococcus and other genera of lactic acid bacteria (LAB), have attracted considerable attention due to their activity against spoilage and pathogenic bacteria3. Several of these peptides have proven applications as natural preservatives in the food industry or have potential as alternatives to antibiotics in veterinary and medical applications4,5,6. These bacterial AMPs, also called bacteriocins, comprise a heterogeneous family of gene-encoded, small ribosomally synthesized peptides with a narrow or broad inhibitory spectrum2, 7.
According to Cotter et al.4, bacteriocins are classified into those which are either post-translationally modified (class I) and unmodified or minimally modified peptides (class II)2, 5. Class II bacteriocins are further subdivided into pediocin-like bacteriocins (class IIa), two-peptide bacteriocins (class IIb), circular bacteriocins (class IIc) and those that cannot be assigned to any of the other bacteriocin subgroups (class IId)2, 8. Class IIa bacteriocins are the most extensively studied and are characterized by their strong inhibitory effect on Listeria sp.9, with pediocin PA-1 being the best-known of this class.
Generally, the production of class II bacteriocins requires several genes clustered into one or more operons encoded either on plasmids or on the chromosome. A typical gene cluster consists of a structural gene that encodes the prepeptide (or two structural genes for the two-peptide bacteriocins), and individual genes that encode an immunity protein, an ATP-binding cassette (ABC) transporter, an accessory protein for extracellular translocation of the bacteriocin, and in several cases, two regulatory proteins10. Bacteriocins are usually ribosomally synthesized as inactive prepeptides with an N-terminal leader sequence which is attached to a C-terminal propeptide. The leader peptide is often cleaved at a double-glycine proteolytic processing site during export by an ABC transporter and the mature peptide is released into the external environment. However, a few bacteriocins contain an N-terminal sec-dependent sequence leader and are secreted by general sec-dependent export systems11,12,13.
Characterization of bacteriocins and assessment of their applications often requires that they are obtained in a pure form from the culture medium or by chemical synthetic production. However, bacteriocin yields from the native host are often very low and the methods involved in their purification and chemical synthesis can be laborious and expensive14,15,16. The development of heterologous expression systems for bacteriocin production in different hosts such as E. coli, LAB and yeasts offers several advantages over native systems, including increased production levels, control of bacteriocin gene expression and characterization of bacteriocin systems from genera or species where there are no genetic tools available17,18,19. Compared to other hosts, E. coli is attractive because of its extensive genetic characterization and the availability of versatile cloning tools and expression systems. Indeed, there are many examples of the cloning of genes involved in the biosynthesis of class II bacteriocins in E. coli, such as mesentericin Y10520, gassericin A21, piscicolin 12622, pediocin PA-1 and AcH23,24,25,26,27, divercin V4117, 27, 28, enterocin A29, enterocin P30, enterocin CRL3531 and sakacin P16. However, there can also be some limitations to using E. coli as an expression host for LAB bacteriocins due to its different genetic background and its limited ability to facilitate extensive disulfide bond formation17, 32. To avoid the differences in codon usage that can impede translation due to the demand for tRNAs that are rare or lacking in the expression host, many bacteriocins have been expressed as optimized sequences, replacing the rare codons with synonymous codons, fused to a secretion signal sequence to deliver them to the cytoplasm or expressed in E. coli RosettaTM strains which supply tRNAs for the six codons rarely used in E. coli 31. Bacteriocins have also been expressed as thioredoxin fusion proteins in trxB/gor mutant (a thioredoxin reductase and gluthatione reductase knockout) E. coli to enhance solubility and the establishment of disulfide bonds17, 22. However, using bacteriocin fusion proteins requires an additional cleavage step and further purification to access the mature bacteriocin.
Here, we report the heterologous expression of two disulphide bond-containing class II bacteriocins, the recently described class IId bactofencin A8 and the well-known class IIa pediocin PA- 133, directly into the medium, by cloning their native genes from the natural producers without additional trx gen or signal secretion sequences. This represents the first report of bactofencin A production in a Gram-negative organism. In addition, we identify the minimum gene cluster required for the expression of the respective bacteriocins in E. coli and open up new perspectives for the biotechnological production and modification of these and other bacteriocins.
Results
Cloning of bactofencin A (bfn) and pediocin PA-1 (ped) gene clusters in E. coli
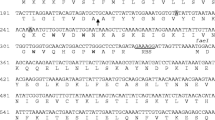
Bactofencin A, produced by Lactobacillus salivarius DPC6502, is a highly basic 22-amino-acid peptide containing an intramolecular disulfide bond between Cys7 and Cys22 (Fig. 1a), with activity against pathogenic species including Staphylococcus aureus and Listeria monocytogenes 8. Pediocin PA-1 is a 44 amino acid peptide with a strong anti-Listeria activity containing four cysteine residues which form two disulfide bonds between Cys9 and Cys14 and between Cys24 and Cys44 (Fig. 1b)33 produced by several strains of Pediococcus and Lactobacillus species15.

Amino acid sequence of (a) pre-bactofencin A and (b) pre-pediocin PA-1 and the mature sequences formed after processing of the leader peptide and oxidation of the cysteines. The black arrow indicates the leader peptide cleavage site.
Initial attempts to express bactofencin A in E. coli under a constitutive promoter were unsuccessful in that growth of the bacterium was retarded and the plasmid was rapidly lost. To circumvent this problem, the bactofencin A gene cluster (4,110 bp) from Lb. salivarius DPC6502 and the pediocin PA-1 (3,386 bp) operon from Pediococcus acidilactici LMG 2351 and different combinations of the genes within these operons were cloned into pMPB1 (Figs S1 and S2), a modified pETcocoTM-2 (Novagen) vector. The plasmid pETcocoTM-2 has been designed for cloning and expression of toxic proteins through dual control of expression: at transcriptional level by the T7 phage promoter inducible by IPTG, and for amplification at the DNA replication level by L-arabinose34. To generate the bactofencin A- and pediocin PA-1-operon containing plasmids, the genes of interest were amplified by PCR, assembled and joined with restriction enzyme-digested pMPB1 vector by an In-Fusion reaction. Through this process the entire bactofencin A (pMPB1-bfnAbfnIDLSL_0052DLSL_0053) and pediocin PA-1 (pMPB1-pedApedBpedCpedD) gene clusters were successfully cloned under the control of T7lac promoter. In order to evaluate the effect of the expression of one or more genes on heterologous bacteriocin production, 16 chimeric plasmids containing different combinations of genes were constructed (Table 1, Fig. S2).
Induction conditions for bactofencin A and pediocin PA-1 expression in E. coli
To determine the optimum time required for bacteriocin production using the T7lac promoter system, E. coli TunerTM (DE3) cells carrying the control plasmid (pMPB1) and bfn vector (pMPB1-bfnAbfnIDLSL_0052DLSL_0053) were grown in the presence of glucose (low-copy state (LC)) or arabinose (medium-copy state (MC)). Since IPTG concentrations above 100 µM compromised the growth of the producer strains after 24 hours (data not shown), the time for optimum production was determined by collecting samples each hour for 8 hours and at 24 hours in the presence of 25 µM of IPTG. The results showed bactofencin A activity between 2 and 8 hours after induction in LC conditions and between 1 and 4 hours in the MC state (Fig. S3). Therefore, subsequent experiments were performed following 3 hours of induction to ensure bacteriocin expression.
To control both plasmid copy number and expression, E. coli TunerTM (DE3) cells transformed with the control plasmid, bfn and ped (pMPB1-pedApedBpedCpedD) vectors, were grown in LB supplemented with glucose or arabinose for 3 hours in the presence of 0, 25, 50, 75 and 100 µM of IPTG. The results demonstrate that the expressed bactofencin A in the cell-free extracellular fraction is biologically active against the indicator Lactobacillus delbrueckii subsp. bulgaricus LMG6901 in both low copy (LC2) and medium copy (MC2) states under the IPTG assay concentrations (Fig. 2a). The MC2 condition also showed an inhibitory zone when the system was not induced, suggesting a basal bactofencin A expression level. However, the activity of supernatants in the presence of IPTG was lower than those expressed at LC at the same IPTG concentration. As shown in Fig. 2b, the production of active pediocin PA-1 against Listeria innocua DPC3572 was only achieved at low copy level (LC3), increasing from 25 µM to 50 µM IPTG concentration.

Production of bactofencin A and pediocin PA-1 in E. coli. Antibacterial activity of (a) bactofencin A and (b) pediocin PA-1 produced by supernatants of E. coli TunerTM (DE3) carrying the control (LC1, MC1), bfn (LC2, MC2) and ped (LC3, MC3) vectors at different levels of plasmid amplification (low copy (LC) and medium copy (MC)) with 0, 25, 50, 75 and 100 µM of IPTG after 3 h of induction. Lb. bulgaricus LMG6901and L. innocua DPC3572 were used as the indicator strains for bactofencin A and pediocin PA-1 activity, respectively.
Taken together, these results demonstrate the production of active bactofencin A and pediocin PA-1 directly into the extracellular medium by E. coli TunerTM (DE3), with the low copy state being the most suitable condition for the expression of both bacteriocins. In addition, 25 µM IPTG was selected for the induction of bacteriocin production as it was the lowest concentration that showed activity.
Expression of the bacteriocin transporter genes is essential for bfn and ped activity in E. coli
Bfn and ped operons are composed of four genes that encode the structural peptide (bfnA, pedA), the immunity protein (bfnI, pedB), an ABC transporter (DLSL_0052, pedD) and an accessory protein (DLSL_0053, pedC). To determine the genes required for heterologous bacteriocin production, two different groups of plasmids were constructed, vectors from which had each gene individually deleted and vectors designed to express the structural gene in the presence of the immunity, transporter or accessory encoded genes.
The results revealed that the cell-free supernatants contained active antimicrobials exclusively in the presence of the transporters but independent of the presence of the accessory gene in both cases (Fig. 3). Interestingly, small zones of inhibition were also observed in the lysates of bfn constructions containing the structural gene alone or in combination with the DLSL_0053 gene (Fig. 3a). In the case of pediocin PA-1, only the lysate of the strain containing the whole ped operon showed activity (Fig. 3b).

Antimicrobial activity of cell-free supernatants (SN) and lysates (L) of E. coli TunerTM (DE3) after 3 hours of 25 µM IPTG induction. (a) Analysis of bactofencin A activity against Lb. bulgaricus and (b) pediocin PA-1 activity againt L. innocua. The genes that encode the structural peptide of bactofencin and pediocin are indicated in green and purple arrows, respectively. The immunity, the transporter and the accessory protein encoded genes are shown in yellow, blue and red arrows. The black arrow indicates the T7lac promoter. Samples from the natural producers Lb. salivarius DPC6502 and P. acidilactici LMG2351 are indicated as Bfn and Ped.
The quantification of the relative extracellular bactofencin A activity produced by bfn constructs was 2-fold higher (640 BU/ml) than that produced by the whole bfn operon under the conditions assayed (Fig. 4a). By contrast, the pediocin PA-1 constructions showed the same production level in the whole operon vector and pedB mutant (160 BU/ml) and a 2-fold reduction in pedC and pedCD mutants (Fig. 4b), indicating that pedC is probably involved in the optimal activation of pediocin PA-1 produced by E. coli. Although the amount of bactofencin A and pediocin PA-1 produced by E. coli was found to be 4 and 8-fold lower than the bacteriocin produced by an overnight culture of the natural producers, it should be noted that this level of bacteriocin production in E. coli is reached in only 3 hours. In addition, these results suggest that the bactofencin A operon can be simplified for heterologous expression, requiring only the transporter gene and the structural gene itself.

Relative quantification of cell-free supernatants (Bacteriocin units (BU)/ml). Antimicrobial activity of (a) bactofencin A and (b) pediocin PA-1 produced by E. coli transformed with the plasmids APC2313 (control), APC2315, APC2318, APC2321 and APC2322, and APC2658, APC2663, APC2664 and APC2666, respectively, in comparison with the samples from the natural producers Lb. salivarius DPC6502 (Bfn) and P. acidilactici LMG2351 (Ped). *Bacteriocin activity (BU/ml) was calculated as the inverse of the highest dilution showing inhibition of indicator strains divided by volume (0.05 ml).
Purification of the active bfn and ped supernatants from E. coli and MALDI TOF mass spectrometry analysis of bacteriocin
The sequence analysis of DLSL_0053 (bactofencin A transport accessory gene) revealed the presence of the cd02947, a thioredoxin family domain, suggesting that this protein is involved in disulfide bond formation. To evaluate the presence of a disulfide bond in the active bfn, the supernatants from the natural producer Lb. salivarius and TunerTM(DE3) E. coli carrying the control plasmid, the entire bfn operon, and the constructs without the accessory gene (pMPB1- bfnAbfnIDLSL_0052), the immunity gene (pMPB1- bfnADLSL_0052DLSL_0053) and without both (pMPB1- bfnADLSL_0052) were purified and analysed for the presence of bactofencin A by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) in positive ion reflectron mode. Interestingly, the results showed that all fractions, with the exception of the negative control, contained the 2,782 Da bactofencin A mass, indicating that the intramolecular disulfide bond formed in all cases, independent of the presence of the accessory gene (Fig. 5a).

MALDI-TOF mass spectrometry analysis. (a) Soluble extracellular fractions from bactofencin A E. coli producers and (b) pediocin PA-1 producers after 3 hours of IPTG induction. Samples from the natural producers Lb. salivarius DPC6502 and P. acidilactici LMG2351 are indicated as Bfn and Ped.
MALDI TOF analysis in reflectron mode was also used to confirm the presence of both disulphide bonds in pediocin PA-1 constructs but the concentration of sample pMPB1-pedApedD was too low to successfully detect the monoistopic mass of pediocin in this sample. Instead pediocin PA-1 samples were analysed in positive ion linear mode where ions were averaged. MALDI TOF MS analysis of pediocin PA-1 fractions from E. coli showed that in addition to the expected mass of 4,625 Da, corresponding to the pediocin PA-1 synthesized by the natural producer P. acidilactici, a 4641 Da mass, consistent with the oxidation of methionine, was also detected (Fig. 5b).
Increasing the heterologous production of bfn in E. coli
The level of production of active bactofencin A using the simplest system, consisting of both structural and transporter genes, was 640 BU/ml under 25 µM of IPTG of induction. This corresponds to approximately 25% of the amount synthesized by the natural producer Lb. salivarius DPC6502 and prompted investigations to determine if the synthesis of active bacteriocin could be improved by increasing the inducer concentration. To do that, E. coli TunerTM (DE3) transformed with pMPB1-bfnADLSL_0052 culture, was induced with 100 µM IPTG and the activity of the supernatant analysed after 3 h of induction. As shown in Fig. 6, the relative quantity of active bactofencin A in 3 hours reached approximately 50% of that produced by the natural producer in an overnight culture.

Quantification of the expression of bactofencin A produced by E. coli Tuner (DE3) transformed with the plasmid containing the structural and the transporter encoded genes after 3 h of 100 µM ITPG induction. The activity of the supernatant from an overnight culture of Lb. salivarius is indicated as Bfn. *Bacteriocin activity (BU/ml) was calculated as the inverse of the highest dilution showing inhibition of Lb. bulgaricus indicator strain divided by volume (0.05 ml).
Discussion
Bacteriocins produced by LAB offer a great variety of different applications for the food, health and pharmaceutical industries5, 35. However, the high cost of bacteriocin purification from natural producers, or chemical synthesis, can limit their large-scale production. This has led to the exploration of alternative heterologous expression hosts such as E. coli. To achieve high-level production of native bacteriocins in this host it is also necessary to express the specialized secretion machinery required for the recognition and processing of their prepeptides. However, the use of systems involving, for example, T7 RNA polymerase can result in cellular toxicity arising from overexpression of secretion machinery and other integral membrane proteins26. For these reasons, in this study pETcocoTM-2 was selected as a backbone vector to overexpress the bactofencin A and pediocin PA-1 operons. The dual replicon nature of this vector facilitates cloning under the T7lac promoter and expression of toxic proteins due to the extremely low basal expression level in the single copy state which promotes the stability of recombinant plasmids, while allowing the amplification of plasmid DNA copy number by arabinose induction and/or the increase of protein expression by adding IPTG. In this work, the PCR-amplified genes of interest from Lb. salivarius DPC6502 and P. acidilactici LMG2351, encoding bactofencin A and pediocin PA-1 operons, respectively, were cloned into the linearized pMPB1 vector, based on the commercial plasmid pETcocoTM-2. This facilitated the successful generation of stable expression vectors with the entire bfn and ped operons as well as the gene-deleted mutants by assembling the remaining genes of interest, avoiding laborious gene inactivation approaches. In addition, the use of pETcocoTM-2 derived vectors has allowed the identification of the optimal conditions (low copy number and <100 μM IPTG concentration) for each bacteriocin expression without compromising bacterial growth. Interestingly, the production of active bacteriocin in the transformed E. coli TunerTM (DE3) cells was not proportional to copy number, implying that the bactofencin A antimicrobial activity decreased at the medium-copy state and it became undetectable in the pediocin PA-1 constructs. Considering that the growth rate of the cultures was similar to the control (data not shown), the induction of plasmid copy number seems to interfere with the ability of E. coli to overproduce these bacteriocins. This can be explained by many factors, including the structural features of the recombinant gene sequences, the stability and translational efficiency of the mRNA, incomplete processing of signal sequences, the metabolic burden on the host strain, the capacity of the transport system and the potential toxicity of the bacteriocin14, 36,37,38. Thus, a low copy vector was the most suitable condition for evaluating the expression of both bacteriocins.
Previous genome sequencing of Lb. salivarius DPC 6502 suggested that bactofencin A biosynthesis involves four genes bfnA, bfnI, DLSL_0052 and DLSL_0053 organized in a single operon8. In the present work, we obtained a full reconstitution of the in vivo synthesis of bactofencin A in a Gram-negative host. TunerTM (DE3) E. coli cells harbouring the complete bfn gene cluster successfully inhibited Lb. bulgaricus. Moreover, the analysis of expression of gene-deleted mutants demonstrated that only the putative ABC transporter (DLSL_0052) is essential for bacteriocin export into the extracellular medium. The ABC transporter contains an N-terminal peptidase C39 domain responsible for the cleavage of double-glycine leader sequences10, 39. In the absence of the transporter, it was not possible to detect antimicrobial activity in supernatants, supporting the theory that this protein plays an essential role in the processing and secretion of the mature active bacteriocin. In addition, some activity was found in the lysate of E. coli carrying plasmids, encoding the single structural peptide together with the putative bactofencin A transport accessory protein (DLSL_0053), suggesting that a small amount of prepeptide could be processed by proteases present in the host. The sequence analysis of DLSL_0053 revealed the presence of cd02947, a thioredoxin family domain involved in the formation of disulfide bonds. However, the DLSL_0053 gene deletion proved that this protein is not required for bactofencin A activity in E. coli. The molecular mass of bactofencin A from active supernatants detected by MALDI-TOF MS analysis was 2,782 Da in all the vectors including those without the transporter accessory gene, indicating the presence of a disulfide bond. Considering this result, it could be hypothesized that the processed peptides could go through the periplasm which contains the enzyme DsbA which catalyzes disulfide bond formation40.
The present study shows that the production of pediocin PA-1 also requires the presence of the structural gene and the ABC transporter. This is in contrast to the results obtained by Venema et al.41 who concluded that PedC was also essential for pediocin PA-1 secretion, and supports the results reported by other authors24, 25. However, it is of interest to note that the amount of active pediocin produced by E. coli increases in the presence of pedC, suggesting that the accessory protein PedC enhances active pediocin PA-1 production in E. coli. These results are in accordance with those described previously by Oppegard et al.42 who indicated that PedC is important to ensure the correct formation of disulfide bonds in pediocin PA-1. In contrast, MALDI-TOF MS analysis of the purified pediocin PA-1 supernatants from E. coli showed an additional peptide fragment of 4,641 Da that is not present in the fractions from the natural producer P. acidilactici (4,625 Da). The 16 Da molecular mass difference is explained by the oxidation of methionine residues to MetSO during the recombinant production as it has been described in other heterologous hosts such as yeasts43, 44 and LABs45,46,47. To protect the peptide from oxidation, Met could be replaced by either Ala, Ile or Leu, with minor effects on the bacteriocin activity48.
In general, overexpressed recombinant proteins in E. coli are accumulated either in the periplasmic space or in the cytoplasm in inclusion bodies so additional purification steps are required to obtain pure active peptide49. In contrast, we show that the use of derived pETcoco-2 vectors together with the TunerTM (DE3) E. coli host allows the expression of active bacteriocins directly into the medium and, therefore, could be used to simplify the purification process. Remarkably, both secreted bacteriocins contained disulfide bonds, overcoming one of the major limitations of disulfide bond production in E. coli.
This is, to our knowledge, the first description of the production of bactofencin A in E. coli and the functional analysis of the genes involved in their heterologous production. The analysis revealed that only the structural gene and the transporter encoded gene are required for expression of fully functional bacteriocins. Induction with 100 µM IPTG led to an increase in the bactofencin A production, reaching in 3 hours 50% of the bactofencin A produced by the natural producer Lb. salivarius in an overnight culture. Even though the levels of heterologous production of bactofencin A and pediocin PA-1 in E. coli were lower than those observed in the natural producer, its recovery from the supernatants may be faster, easier and more cost effective.
The results presented here suggest that this system could be useful for expressing new bacteriocin operons described in silico as well as their characterization at molecular level when there is no access to the original producer strains or the transformation efficiencies of natural producers is a severe impediment for gene analysis, opening the door to the heterologous expression and harnessing of the potential of new bacteriocins in the future.
Methods
Bacterial strains, plasmids and culture conditions
Bacterial strains and plasmids used in this study are listed in Table 1. Escherichia coli HST08 strain StellarTM (Takara Bio USA, Inc, Mountain View, CA), was used for standard cloning procedures and Escherichia coli BL21 strain TunerTM (DE3) (also called E. coli TunerTM (DE3)) (Novagen, EMD Millipore, Billerica, MA) was used for gene expression experiments. Escherichia coli strains were grown aerobically at 180 r.p.m. and 37 °C in Luria-Bertani (LB) medium and supplemented with ampicillin (50 μg/ml) for the selection and maintenance of plasmids. Lactobacillus salivarius DPC6502, used as a bactofencin A producer, and the bactofencin A-sensitive strain Lactobacillus delbrueckii subsp. bulgaricus LMG6901 (Lb. bulgaricus LMG 6901) were grown under anaerobic conditions at 37 °C in MRS medium (Difco Laboratories, Detroit, MI). Anaerobic conditions were maintained with the use of anaerobic jars and Anaerocult A gas packs (Merck, Darmstadt, Germany). Pediococcus acidilactici LMG2351, used as a pediocin PA-1 producer, was cultured aerobically in MRS broth at 30 °C without shaking. The pediocin PA-1-sensitive indicator strain Listeria innocua DPC 3572 was grown aerobically at 37 °C in brain heart infusion (BHI) medium (Merck) without shaking. Gene expression was induced by IPTG (isopropyl- β-D-thiogalactopyranoside). IPTG was supplied by Fisher Scientific (Dublin, Ireland) dissolved in water, filter sterilized, and added to the media. (D) + -glucose, L- + -Arabinose and other chemical reagents were obtained from Sigma Aldrich (Arklow, Ireland). Pediococcus acidilactici LMG2351 was a gift from Dzung Bao Diep, Laboratory of Microbial Gene Technology, Agricultural University of Norway, Ås, Norway.
Molecular biology general procedures
All DNA manipulations were performed following standard protocols50. All oligonucleotides used in this study are described in Supplementary Tables S1 and S2. The plasmid pETcocoTM-2 (Novagen) was used for gene construction and expression. Firstly, the multiple cloning site (MCS) was replaced by the product of annealing of MCS F SphI and MCS R AvrII primers, digested with SphI-AvrII and cloning into pETcocoTM-2 SphI-AvrII, generating plasmid pMPB1 (APC2313). A detailed description of the cloning of the bactofencin A and pediocin PA-1 gene fragments into the linearized pMPB1 is provided in the Supplementary information. The primers were designed according to the instructions of In-Fusion HD cloning and they were obtained from Sigma Aldrich (Arklow, Ireland). Briefly, the bactofencin A and pediocin PA-1 sequences were amplified by polymerase chain reaction (PCR) using CloneAmp HiFi PCR premix (Takara Bio USA, Inc, Mountain View, CA) and genomic DNA from L. salivarius DPC 6502 and plasmid DNA from P. acidilactici LMG2351 were used as DNA templates, respectively. The PCR products were eluted with illustra GFX PCR DNA and Gel Band Purification kit (GE Healthcare, Buckinghamshire, UK) and inserted into the linearized pMPB1 vector using In-Fusion HD cloning Plus (638910, Takara Bio USA, Inc, Mountain View, CA) as described previously51. The constructions were transformed into StellarTM competent cells and colonies were selected on plates containing ampicillin 50 μg/ml) and D (+)-glucose (0.2%). The transformants were confirmed by colony PCR reactions. Then, the number of copies of the recombinant plasmids was increased by L-arabinose induction before plasmid DNA isolation was analysed by double digestion and sequencing. All DNA sequencing were performed by GATC Biotech (Koln, Germany). The resulting vectors are summarised in the Table 1 and Fig. S2. Restriction enzymes and T4 DNA ligase were obtained from New England Biolabs (Ipswich, M). Genomic and plasmid DNA were isolated using GenEluteTM Bacterial Genomic DNA kit (Sigma Aldrich, St.Louis, MO) and NucleoSpin® plasmid kit (Macherey-Nagel, Duren, Germany), respectively.
Induction of bacteriocin expression
Bactofencin A and pediocin PA-1 production were tested under different expression conditions (plasmid copy number and IPTG concentration) in order to find the best condition to induce bacteriocin production. For this purpose, TunerTM (DE3) E. coli cells were transformed with the control vector (pMPB1) and those carrying bfn (pMPB1- bfnAbfnIDLSL_0052 DLSL_0053) and ped operons (pMPB1-pedApedBpedCpedD), and isolated colonies were grown in LB broth containing ampicillin (50 μg/ml) and 0.2% D(+)-glucose (LBAp50Glc0.2), which maintains the plasmid in a single-copy state, for 16 hours at 37 °C with shaking. The cultures were diluted 1:50 into fresh LBAp50Glc0.2 medium to keep the low copy state (LC) and fresh LB containing ampicillin (50 μg/ml) and L-+-Arabinose (0.01%) to increase the number of copy of plasmids (Medium Copy state (MC)). When cells reached OD600 0.5–0.6, IPTG was added at concentrations of 0, 25, 50, 75 and 100 µM and the cultures were incubated for 3 h at 37 °C before determining the antimicrobial effect of the supernatants against the indicator strains as described in bacteriocin activity assay.
Bacteriocin activity assay
Bactofencin A and pediocin PA-1 antimicrobial activities were determined by the well-diffusion method against the indicator strain L. delbrueckii subsp. bulgaricus LMG6901 and L. innocua DPC3572, respectively. Briefly, 5 ml of transformed E. coli cells were centrifuged at 4,000 × g at 4 °C for 15 minutes. The bacterial pellets were washed and resuspended in 5 ml of PBS solution and further lysed via sonication (5 pulses at 16 W, 15 s/pulse and intervals of 45 s on ice) in a MSE Soniprep 150 (MSE, London, UK) and centrifuged at 4,000 × g at 4 °C for 15 min. The resulting cell-free intracellular fraction from the lysate and the cell-free supernatant of the cultures were filtered with 0.20 μm membrane filters (Sartorius, Germany). 50 μl of cell-free supernatants (SN) and lysates (L) of each culture were applied to 5 mm diameter wells on MRS 1% agar plates containing L. delbrueckii subsp. bulgaricus LMG6901 and BHI 0.8% agar plates containing L. innocua DPC3572. MRS and BHI agar plates were refrigerated for two hours prior to incubation at 37 °C under anaerobic and aerobic conditions, respectively, for 24 hours.
As positive controls of bactofencin A and pediocin PA-1 production, the natural producers L. salivarius DPC6502 and P. acidilactici LMG2351 were grown in MRS broth at 37 °C (under anaerobic conditions) and 30 °C (aerobic conditions) for 16 h, respectively. Cell-free culture supernatants were obtained by centrifugation of cultures at 4,000 × g at 4 °C for 15 min. The pH was adjusted to pH 7 using 1 M NaOH to avoid any antimicrobial activity caused by the acidity of the cell free supernatants and filtered through 0.20 μm pore-size filters. The pellets were sonicated as described previously.
To quantify the relative active bacteriocin production, 50 µl of two-fold serial dilutions of each sample were placed in wells. The bacteriocin activity was expressed in bacteriocin units per ml (BU ml−1) and was defined as the reciprocal of the highest dilution showing inhibition of the indicator strain.
Purification and MALDI TOF mass spectrometry analysis of bfn and ped
Twenty five ml of culture of L. salivarius DPC 6502 and E. coli Tuner DE3 containing the bacteriocin expression plasmids were harvested by centrifugation at 4,000 × g at 4 °C for 20 min, the bactofencin A supernatants were applied to a column containing 2 ml SP Sepharose fast-flow cation-exchange resin (GE Healthcare, United Kingdom), equilibrated with buffer A [20 mM potassium phosphate buffer and 25% acetonitrile, pH 2.5]. The column was then washed with buffer A and with 10 ml buffer B (buffer A containing 500 mM KCl) and peptides eluted with 10 ml of Buffer C (buffer A containing 1000 mM KCl). The 1 M KCl eluents were desalted using C18 SPE zips (Millipore, Cork, Ireland). Zip tips were primed by washing with 3 × 10 µl aliquots of 100% acetonitrile followed by 5 × 10 µl aliquots of 0.1% TFA. The sample was bound to the zip tip by repeated pipetting of a 50 µl aliquot of sample, the tip was washed with 3 × 10 µl aliquots of 0.1% TFA and bactofencin A eluted by repeated pipetting in 3 µl CHCA matrix. Samples were analysed for the presence of bactofencin A by MALDI TOF mass spectrometry (Axima- TOF2; Shimadzu Biotech, Manchester, United Kingdom) in positive ion reflectron mode.
One hundred ml of pediocin PA-1 samples were applied to 500 mg 6 ml Strata C18-E SPE columns pre-equilbrated with methanol and water. The columns were washed with 6 ml 30% ethanol and the samples eluted with 6 ml of IPA (70% propan-2-ol, 0.1% TFA). The IPA eluents were analysed by MALDI TOF mass spectrometry in positive ion reflectron and linear mode.
References
Hiemstra, P. S. & Zaat, S. A. J. Antimicrobial Peptides and Innate Immunity. Progress in Inflammation Research. Springer Basel (ed. Parnham, M. J. and Faist, E.), doi:10.1007/978-3-0348-0541-4 (2013).
Rea, M. C., Ross, R. P., Cotter, P. D. & Hill, C. In Prokaryotic Antimicrobial Peptides: From genes to applications. Springer (ed. Drider, D. and Rebuffat, S.) 29–53, doi:10.1007/978-1-4419-7692-5 (2011).
Vaughan, A., Eijsink, V. G. & Van Sinderen, D. Functional characterization of a composite bacteriocin locus from malt isolate Lactobacillus sakei 5. Applied and environmental microbiology 69, 7194–720, doi:10.1128/AEM.69.12.7194-7203.2003 (2003).
Cotter, P. D., Hill, C. & Ross, R. P. Bacteriocins: developing innate immunity for food. Nature reviews. Microbiology 3, 777–788, doi:10.1038/nrmicro1273 (2005).
Cotter, P. D., Ross, R. P. & Hill, C. Bacteriocins - a viable alternative to antibiotics? Nature reviews. Microbiology 11, 95–105, doi:10.1038/nrmicro2937 (2013).
Hassan, M., Kjos, M., Nes, I. F., Diep, D. B. & Lotfipour, F. Natural antimicrobial peptides from bacteria: characteristics and potential applications to fight against antibiotic resistance. Journal of applied microbiology 113, 723–736, doi:10.1111/j.1365-2672.2012.05338.x (2012).
Perez, R. H., Zendo, T. & Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): various structures and applications. Microbial cell factories 13(Suppl 1), S3, doi:10.1186/1475-2859-13-S1-S3 (2014).
O’Shea, E. F. et al. Bactofencin A, a new type of cationic bacteriocin with unusual immunity. mBio 4, e00498–00413, doi:10.1128/mBio.00498-13 (2013).
Cui, Y. et al. Class IIa bacteriocins: diversity and new developments. International journal of molecular sciences 13, 16668–16707, doi:10.3390/ijms131216668 (2012).
Nes, I. F. et al. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie van Leeuwenhoek 70, 113–128, doi:10.1007/BF00395929 (1996).
Sanchez, J. et al. Amino acid and nucleotide sequence, adjacent genes, and heterologous expression of hiracin JM79, a sec-dependent bacteriocin produced by Enterococcus hirae DCH5, isolated from Mallard ducks (Anas platyrhynchos). FEMS Microbiology Letters 270, 227–236, doi:10.1111/j.1574-6968.2007.00673.x (2007).
Cintas, L. M., Casaus, P., Havarstein, L. S., Hernandez, P. E. & Nes, I. F. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Applied and environmental microbiology 63, 4321–4330 (1997).
De Kwaadsteniet, M., Fraser, T., Van Reenen, C. A. & Dicks, L. M. Bacteriocin T8, a novel class IIa sec-dependent bacteriocin produced by Enterococcus faecium T8, isolated from vaginal secretions of children infected with human immunodeficiency virus. Applied and environmental microbiology 72, 4761–4766, doi:10.1128/AEM.00436-06 (2006).
Rodriguez, J. M., Martinez, M. I., Horn, N. & Dodd, H. M. Heterologous production of bacteriocins by lactic acid bacteria. International journal of food microbiology 80, 101–116, doi:10.1016/S0168-1605(02)00153-8 (2003).
Rodriguez, J. M., Martinez, M. I. & Kok, J. Pediocin PA-1, a wide-spectrum bacteriocin from lactic acid bacteria. Critical reviews in food science and nutrition 42, 91–121, doi:10.1080/10408690290825475 (2002).
Chen, H. et al. Cloning and heterologous expression of a bacteriocin sakacin P from Lactobacillus sakei. Escherichia coli. Applied microbiology and biotechnology 94, 1061–1068, doi:10.1007/s00253-012-3872-z (2012).
Richard, C., Drider, D., Elmorjani, K., Marion, D. & Prevost, H. Heterologous expression and purification of active divercin V41, a class IIa bacteriocin encoded by a synthetic gene in Escherichia coli. Journal of bacteriology 186, 4276–4284, doi:10.1128/JB.186.13.4276-4284.2004 (2004).
Axelsson, L., Katla, T., Bjornslett, M., Eijsink, V. G. & Holck, A. A system for heterologous expression of bacteriocins in Lactobacillus sake. FEMS Microbiology Letters 168, 137–143, doi:10.1111/j.1574-6968.1998.tb13266.x (1998).
Caetano, T., Krawczyk, J. M., Mosker, E., Sussmuth, R. D. & Mendo, S. Heterologous expression, biosynthesis, and mutagenesis of type II lantibiotics from Bacillus licheniformis. Escherichia coli. Chemistry & biology 18, 90–100, doi:10.1016/j.chembiol.2010.11.010 (2011).
Biet, F., Berjeaud, J. M., Worobo, R. W., Cenatiempo, Y. & Fremaux, C. Heterologous expression of the bacteriocin mesentericin Y105 using the dedicated transport system and the general secretion pathway. Microbiology 144(Pt 10), 2845–2854, doi:10.1099/00221287-144-10-2845 (1998).
Kawai, Y. et al. Heterologous expression of gassericin A, a bacteriocin produced by Lactobacillus gasseri LA39. Animal Science Journal 74, 45–51, doi:10.1046/j.1344-3941.2003.00085.x (2003).
Gibbs, G. M., Davidson, B. E. & Hillier, A. J. Novel expression system for large-scale production and purification of recombinant class IIa bacteriocins and its application to piscicolin 126. Applied and environmental microbiology 70, 3292–3297, doi:10.1128/AEM.70.6.3292-3297.2004 (2004).
Moon, G. S., Pyun, Y. R. & Kim, W. J. Expression and purification of a fusion-typed pediocin PA-1 in Escherichia coli and recovery of biologically active pediocin PA-1. International journal of food microbiology 108, 136–140, doi:10.1016/j.ijfoodmicro.2005.10.019 (2006).
Marugg, J. D. et al. Cloning, expression, and nucleotide sequence of genes involved in production of pediocin PA-1, and bacteriocin from Pediococcus acidilactici PAC1.0. Applied and environmental microbiology 58, 2360–2367 (1992).
Bukhtiyarova, M., Yang, R. & Ray, B. Analysis of the pediocin AcH gene cluster from plasmid pSMB74 and its expression in a pediocin-negative Pediococcus acidilactici strain. Applied and environmental microbiology 60, 3405–3408 (1994).
Miller, K. W., Schamber, R., Chen, Y. & Ray, B. Production of active chimeric pediocin AcH in Escherichia coli in the absence of processing and secretion genes from the Pediococcus pap operon. Applied and environmental microbiology 64, 14–20 (1998).
Ingham, A. B., Sproat, K. W., Tizard, M. L. & Moore, R. J. A versatile system for the expression of nonmodified bacteriocins in Escherichia coli. Journal of applied microbiology 98, 676–683, doi:10.1111/j.1365-2672.2004.02502.x (2005).
Yildirim, S. et al. Production of recombinant bacteriocin divercin V41 by high cell density Escherichia coli batch and fed-batch cultures. Applied microbiology and biotechnology 77, 525–531, doi:10.1007/s00253-007-1188-1 (2007).
Klocke, M., Mundt, K., Idler, F., Jung, S. & Backhausen, J. E. Heterologous expression of enterocin A, a bacteriocin from Enterococcus faecium, fused to a cellulose-binding domain in Escherichia coli results in a functional protein with inhibitory activity against Listeria. Applied microbiology and biotechnology 67, 532–538, doi:10.1007/s00253-004-1838-5 (2005).
Gutierrez, J. et al. Cloning, production and functional expression of enterocin P, a sec-dependent bacteriocin produced by Enterococcus faecium P13, in Escherichia coli. International journal of food microbiology 103, 239–250, doi:10.1016/j.ijfoodmicro.2004.11.035 (2005).
Masías, E. et al. Co-expression and characterization of enterocin CRL35 and its mutant. In Escherichia coli Rosetta. Peptidomics 1, 30-42, doi:10.2478/ped-2014-0003 (2014).
Makrides, S. C. Strategies for achieving high-level expression of genes in Escherichia coli. Microbiological reviews 60, 512–538 (1996).
Henderson, J. T., Chopko, A. L. & van Wassenaar, P. D. Purification and primary structure of pediocin PA-1 produced by Pediococcus acidilactici PAC-1.0. Archives of biochemistry and biophysics 295, 5–12, doi:10.1016/0003-9861(92)90480-K (1992).
Sektas, M. A. S. W. Novel single-copy pETcocoTM vector with dual controls for amplification and expression. inNovations 14 (2002).
Lohans, C. T. & Vederas, J. C. Development of Class IIa Bacteriocins as Therapeutic Agents. International journal of microbiology 2012, 386410–13, doi:10.1155/2012/386410 (2012).
Carrier, T., Jones, K. L. & Keasling, J. D. mRNA stability and plasmid copy number effects on gene expression from an inducible promoter system. Biotechnology and bioengineering 59, 666–672, doi:10.1002/(ISSN)1097-0290 (1998).
Choi, J. H. & Lee, S. Y. Secretory and extracellular production of recombinant proteins using Escherichia coli. Applied microbiology and biotechnology 64, 625–635, doi:10.1007/s00253-004-1559-9 (2004).
Mergulhao, F. J. et al. Medium and copy number effects on the secretion of human proinsulin in Escherichia coli using the universal stress promoters uspA and uspB. Applied microbiology and biotechnology 61, 495–501, doi:10.1007/s00253-003-1232-8 (2003).
Michiels, J., Dirix, G., Vanderleyden, J. & Xi, C. Processing and export of peptide pheromones and bacteriocins in Gram-negative bacteria. Trends in microbiology 9, 164–168, doi:10.1016/S0966-842X(01)01979-5 (2001).
de Marco, A. Strategies for successful recombinant expression of disulfide bond-dependent proteins in Escherichia coli. Microbial cell factories 8, 26, doi:10.1186/1475-2859-8-26 (2009).
Venema, K. et al. Functional analysis of the pediocin operon of Pediococcus acidilactici PAC1.0: PedB is the immunity protein and PedD is the precursor processing enzyme. Molecular microbiology 17, 515–522, doi:10.1111/mmi.1995.17.issue-3 (1995).
Oppegard, C., Fimland, G., Anonsen, J. H. & Nissen-Meyer, J. The Pediocin PA-1 Accessory Protein Ensures Correct Disulfide Bond Formation in the Antimicrobial Peptide Pediocin PA-1. Biochemistry 54, 2967–2974, doi:10.1021/acs.biochem.5b00164 (2015).
Basanta, A. et al. Development of bacteriocinogenic strains of Saccharomyces cerevisiae heterologously expressing and secreting the leaderless enterocin L50 peptides L50A and L50B from Enterococcus faecium L50. Applied and environmental microbiology 75, 2382–2392, doi:10.1128/AEM.01476-08 (2009).
Borrero, J. et al. Cloning, production, and functional expression of the bacteriocin enterocin A, produced by Enterococcus faecium T136, by the yeasts Pichia pastoris, Kluyveromyces lactis, Hansenula polymorpha, and Arxula adeninivorans. Applied and environmental microbiology 78, 5956–5961, doi:10.1128/AEM.00530-12 (2012).
Jimenez, J. J. et al. Cloning, production, and functional expression of the bacteriocin sakacin A (SakA) and two SakA-derived chimeras in lactic acid bacteria (LAB) and the yeasts Pichia pastoris and Kluyveromyces lactis. Journal of industrial microbiology & biotechnology 40, 977–993, doi:10.1007/s10295-013-1302-6 (2013).
Martin, M. et al. Cloning, production and expression of the bacteriocin enterocin A produced by Enterococcus faecium PLBC21 in Lactococcus lactis. Applied microbiology and biotechnology 76, 667–675, doi:10.1007/s00253-007-1044-3 (2007).
Borrero, J. et al. Protein expression vector and secretion signal peptide optimization to drive the production, secretion, and functional expression of the bacteriocin enterocin A in lactic acid bacteria. Journal of biotechnology 156, 76–86, doi:10.1016/j.jbiotec.2011.07.038 (2011).
Johnsen, L., Fimland, G., Eijsink, V. & Nissen-Meyer, J. Engineering increased stability in the antimicrobial peptide pediocin PA-1. Applied and environmental microbiology 66, 4798–4802, doi:10.1128/AEM.66.11.4798-4802.2000 (2000).
Terpe, K. Overview of bacterial expression systems for heterologous protein production: from molecular and biochemical fundamentals to commercial systems. Applied microbiology and biotechnology 72, 211–222, doi:10.1007/s00253-006-0465-8 (2006).
Sambrook, J., Fritsch, E. F. & Maniatis, T. Molecular cloning:a laboratory manual 2nd edn Cold Spring Harbor (1989).
Zhu, B., Cai, G., Hall, E. O. & Freeman, G. J. In-fusion assembly: seamless engineering of multidomain fusion proteins, modular vectors, and mutations. BioTechniques 43, 354–359, doi:10.2144/000112536 (2007).
O’Shea, E. F. et al. Characterization of enterocin- and salivaricin-producing lactic acid bacteria from the mammalian gastrointestinal tract. FEMS Microbiology Letters 291, 24–34, doi:10.1111/j.1574-6968.2008.01427.x (2009).
O’Keeffe, T., Hill, C. & Ross, R. P. Characterization and heterologous expression of the genes encoding enterocin a production, immunity, and regulation in Enterococcus faecium DPC1146. Applied and environmental microbiology 65, 1506–1515 (1999).
Acknowledgements
We are grateful to all members of the laboratory for their insights and helpful suggestions. We also thank Dzung Bao Diep for the gift of Pediococcus acidilactici LMG2351. This work was supported by Science Foundation Ireland (SFI) under grant number SFI/12/RC/2273.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: B.M.-P., P.M.O., M.C.R., P.D.C., C.H. and R.P.R. Performed the experiments: B.M.-P. and P.M.O. Analysed the data: B.M.-P., P.M.O., M.C.R., P.D.C., C.H. and R.P.R. Contributed reagents/materials/analysis tools: B.M.-P., P.M.O., M.C.R., P.D.C., C.H. and R.P.R. Wrote the manuscript: B.M.-P., P.M.O., M.C.R., P.D.C., C.H. and R.P.R.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mesa-Pereira, B., O’Connor, P.M., Rea, M.C. et al. Controlled functional expression of the bacteriocins pediocin PA-1 and bactofencin A in Escherichia coli . Sci Rep 7, 3069 (2017). https://doi.org/10.1038/s41598-017-02868-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-02868-w
- Springer Nature Limited
This article is cited by
-
Bacteriocin diversity, function, discovery and application as antimicrobials
Nature Reviews Microbiology (2024)
-
Two Recombinant Bacteriocins, Rhamnosin and Lysostaphin, Show Synergistic Anticancer Activity Against Gemcitabine-Resistant Cholangiocarcinoma Cell Lines
Probiotics and Antimicrobial Proteins (2024)
-
Heterologous expression of pediocin PA-1 in Pichia pastoris: cloning, expression, characterization, and application in pork bologna preservation
Brazilian Journal of Microbiology (2024)
-
Genomic Analysis of Bacteriocin-Producing Staphylococci: High Prevalence of Lanthipeptides and the Micrococcin P1 Biosynthetic Gene Clusters
Probiotics and Antimicrobial Proteins (2023)
-
Antimicrobial Peptides (Bacteriocins) Produced by Lactococcus lactis and Pediococcus pentosaceus Strains with Activity Against Clinical and Food-Borne Pathogens
Probiotics and Antimicrobial Proteins (2023)