
Abstract
The JAK2V617F mutation is found in most patients with a myeloproliferative neoplasm (MPN). This gain-of-function mutation dysregulates cytokine signaling and is associated with increased accumulation of DNA damage, a process likely to drive disease evolution. JAK2V617F inhibits NHE-1 upregulation in response to DNA damage and consequently represses Bcl-xL deamidation and apoptosis, thus giving rise to inappropriate cell survival. However, the mechanism whereby NHE-1 expression is inhibited by JAK2V617F is unknown. In this study, we demonstrate that the accumulation of reactive oxygen species (ROS) in cells expressing JAK2V617F compromises the NHE-1/Bcl-xL deamidation pathway by repressing NHE-1 upregulation in response to DNA damage. In JAK2V617F-positive cells, increased ROS levels results from aberrant PI3K signaling, which decreases nuclear localization of FOXO3A and decreases catalase expression. Furthermore, when compared with autologous control erythroblasts, clonally derived JAK2V617F-positive erythroblasts from MPN patients displayed increased ROS levels and reduced nuclear FOXO3A. However, in hematopoietic stem cells (HSCs), FOXO3A is largely localized within the nuclei despite the presence of JAK2V617F mutation, suggesting that JAK2-FOXO signaling has a different effect on progenitors compared with stem cells. Inactivation of FOXO proteins and elevation of intracellular ROS are characteristics common to many cancers, and hence these findings are likely to be of relevance beyond the MPN field.
Similar content being viewed by others

Introduction
Forkhead box O (FOXO) transcription factors are evolutionarily conserved proteins involved in multiple physiological and pathological processes.1 In mammals, three principal members of the FOXO subfamily (FOXO1, FOXO3A, FOXO4) are abundantly expressed in most tissues, where they promote cell cycle arrest, DNA repair, apoptosis or detoxification of reactive oxygen species (ROS). In particular, FOXO3A has been shown to have an important role in hematopoiesis.2, 3, 4, 5 The function of FOXO proteins is largely regulated by posttranslational modifications, the main example of which is serine/threonine phosphorylation downstream of the PI3K/AKT signaling pathway.1 Phosphorylated FOXO factors are exported from the nucleus to the cytoplasm and are targeted for proteasomal degradation. This regulation of FOXO proteins is disrupted in several tumor types including breast cancer, prostate cancer, glioblastoma and chronic myeloid leukemia (CML).6
One of the major functions of FOXO proteins is to modulate ROS homeostasis by transcriptionally activating ROS-detoxifying enzymes such as catalase and SOD.1 Maintenance of appropriate levels of intracellular ROS is vital for signaling networks, which mediate multiple physiological processes.7 In several malignancies, perturbation of physiological ROS levels has been found to induce proliferation,8 amplify survival signaling9, 10 and limit anti-tumor cytotoxicity of various therapeutic agents,11, 12, 13 therefore contributing to maintenance and progression of the cancer phenotype. Increased ROS is also a major cause of DNA damage, which leads to chromosomal abnormalities and genome instability.14 Regulation of ROS homeostasis by FOXO may therefore have a significant role in limiting cancer development and progression.
The myeloproliferative neoplasms (MPNs) are hematological malignancies associated with overproduction of mature, functional blood cells.15 These diseases are characterized by a chronic clinical course together with a risk of transformation to acute myeloid leukemia. In the majority of MPN patients, an acquired gain-of-function mutation (V617F) is found in JAK2, a cytoplasmic tyrosine kinase involved in signaling from multiple cytokine receptors. Cells expressing JAK2V617F harbor more DNA damage,16, 17, 18, 19 and of particular note, the JAK2V617F mutation interferes with the normal apoptotic response to DNA damage,20, 21 thus providing a mechanism for the accumulation of DNA damage. In normal cells, DNA damage upregulates the Na+/H+ antiport NHE-1, thereby increasing intracellular pH, which in turn increases Bcl-xL deamidation and promotes apoptosis. This pathway is inhibited by JAK2V617F in primary cells derived from MPN patients, thus permitting increased cell survival despite the presence of increased DNA damage.21 However, the mechanism by which JAK2V617F inhibits NHE-1 upregulation remains unknown.
Here, we demonstrate that JAK2V617F-mediated PI3K signaling inactivates FOXO3A in both JAK2V617F-expressing cell lines and hematopoietic progenitors from MPN patients and knock-in mice. We show that FOXO3A inactivation associates with increased intracellular ROS levels which in turn blocks the NHE-1/Bcl-xL pathway and therefore contributes to increased survival of JAK2V617F-expressing cells following DNA damage. These findings reveal a mechanism whereby JAK2V617F disrupts ROS homeostasis and reduces the normal apoptotic response to DNA damage. Finally, using hematopoietic stem cells (HSCs) and progenitors isolated from MPN patients and the JAK2V617F knock-in mice,22 we also demonstrate that FOXO3A activity downstream of JAK2V617F is distinct in HSCs compared with progenitors and more differentiated cells.
Results
JAK2V617F elevates intracellular ROS levels and increases survival of JAK2V617F-positive cells following DNA damage
Elevated ROS can contribute to altered signal transduction as well as to transformation by a number of oncogenes. We therefore initially investigated the effect of JAK2V617F on intracellular ROS, using a cell-permeable fluorescent dye in IL3 or Epo-dependent Ba/F3-EpoR cells expressing wild-type JAK2 or the constitutively active JAK2V617F. Cells carrying JAK2V617F had higher levels of ROS compared with those cells expressing wild-type JAK2 (Figure 1a). We also examined the effect of endogenous JAK2V617F on ROS levels by using human erythroleukemia (HEL) cells, which harbors multiple copies of mutant JAK2.23 Following the knockdown of JAK2V617F with short interfering RNA (siRNA) (Supplementary Figure S1A), ROS levels were reduced compared with the cells transfected with a control siRNA (Figure 1b). Furthermore, treatment of HEL cells with the selective JAK2 inhibitor TG101209 also reduced ROS (Supplementary Figure S1B). Together these data indicate that exogenous and endogenous JAK2V617F are both associated with increased ROS levels.

JAK2V617F elevates intracellular ROS levels and increases survival following DNA damage. (a) JAK2V617F expression increases intracellular ROS in Ba/F3-EpoR cells. A representative flow cytometry histogram is shown and the bar graph shows mean fluorescence intensity (MFI) relative to that of the wild type. (b) Knockdown of JAK2V617F reduces ROS in HEL cells. A representative flow cytometry histogram is shown and the bar graph shows DCF-DA MFI relative to that of the wild type. (c) JAK2V617F expression in Ba/F3-EpoR cells increases cell survival following DNA damage. A representative flow cytometry plot is shown and the bar graph shows the percentage of apoptotic cells. (d) Knockdown of JAK2V617F sensitizes cells to DNA damage-induced apoptosis. A representative flow cytometry plot is shown and the bar graph shows the percentage of apoptotic cells. All data are averages of three independent experiments and shown as mean±s.e.m. *P<0.05, **P<0.01 and ***P<0.001.
As ROS levels modulate drug sensitivity in some cancers, we then determined the effects of elevated ROS on DNA damage-induced apoptosis. Following treatment with the DNA damage-inducing agent etoposide, Ba/F3-EpoR cells carrying JAK2V617F had reduced apoptosis compared with cells expressing wild-type JAK2 (Figure 1c). In addition, knockdown of endogenous JAK2V617F in HEL cells increased the sensitivity of these cells to etoposide treatment compared with the cells transfected with a control siRNA (Figure 1d). These data demonstrate that JAK2V617F increases cell survival following DNA damage.
JAK2V617F-induced increase in ROS mediates cell survival by inhibiting the NHE-1/Bcl-xL deamidation pathway
To determine whether elevated ROS levels in JAK2V617F-positive cells directly contribute to increased survival following DNA damage, we used MitoQ, a potent antioxidant compound targeted specifically to mitochondria, to reduce ROS levels (Figure 2a, Supplementary Figure S2A). Treatment of HEL or Ba/F3-EpoR cells with etoposide in the presence of MitoQ triggered significantly more cell death at all MitoQ concentrations tested compared with treatment with etoposide alone (Figure 2b, Supplementary Figure S2B). To confirm our results, we also expressed catalase in HEL cells to reduce intracellular ROS levels. Compared with HEL cells transfected with a control construct, those transfected with a catalase expression vector showed an increase in catalase levels (Figure 2c), a small but significant reduction in ROS levels (Figure 2d) and increased apoptosis in response to etoposide (Figure 2e). Together these results indicate that elevated ROS levels induced by JAK2V617F contribute to increased survival of mutant cells following DNA damage.

Elevation of ROS levels by JAK2V617F increases resistance to DNA damage-induced apoptosis. (a) MitoQ treatment reduced ROS levels in HEL cells. Fold decrease in ROS levels following MitoQ treatment relative to the untreated culture is shown. (b) Reduced ROS restored sensitivity to DNA damage-induced apoptosis in HEL cells. (c) Western blot shows catalase expression in HEL cells transfected with an empty pZeo-SV2(+) or catalase expression construct pZeo-CAT. (d) Catalase expression in HEL cells (pZeo-CAT) reduced ROS levels. The bar graph shows mean fluorescence intensity relative to that of the wild type. (e) Reduction of ROS levels increased apoptosis in the presence of DNA damage. (f) Reduction in ROS levels allows NHE-1 upregulation in HEL cells in response to DNA damage. (g) ROS reduction allows Bcl-xL deamidation in HEL cells in the presence of DNA damage. The upper bands (deamidated) and lower bands (native) were quantified using the Image J software and the deamidated protein was shown as a percentage of total Bcl-xL. Data are shown as mean±s.e.m.; n=3. *P<0.05, **P<0.01 and ***P<0.001.
Because elevated H2O2 levels have been shown to suppress both NHE-1 promoter activity and protein expression in non-hematopoietic cells,24, 25, 26 we investigated the possibility that the JAK2V617F-mediated increase in ROS levels might modulate the response of the NHE-1/Bcl-xL pathway to DNA damage. Cells with wild-type JAK2 normally respond to DNA damage by increasing NHE-1 expression and Bcl-xL deamidation leading to apoptosis, but these effects are abrogated by the presence of the JAK2V617F mutation.20, 21 Consistent with these data, treatment with etoposide alone did not induce an increase in NHE-1 expression or Bcl-xL deamidation in JAK2V617F-positive HEL cells (Figures 2f and g). However, following catalase expression, the normal response to etoposide treatment was restored with induction of both NHE-1 upregulation and Bcl-xL deamidation (Figures 2f and g). These results demonstrate that reversing the JAK2V617F-mediated increase in ROS levels restores the response of NHE-1/Bcl-xL pathway to DNA damage.
JAK2V617F signaling reduces catalase expression and increases both phosphorylation and cytoplasmic sequestration of FOXO3A
As catalase is one of the main ROS-detoxifying enzymes in vivo,27 we next investigated the effect of JAK2V617F on endogenous catalase levels. Catalase expression was shown to be increased in HEL cells by either of two selective JAK2 inhibitors (Figure 3a) and also by siRNA knockdown of JAK2 (Figure 3b). Moreover, catalase expression was lower in Ba/F3-EpoR cells carrying JAK2V617F compared with the cells with wild-type JAK2 (Figure 3c). Unlike catalase, both glutathione reductase and SOD2 expression was not increased in HEL cells treated with JAK2 inhibitor, but rather a reduction of SOD2 expression was observed. These data suggest that neither glutathione reductase nor SOD2 is responsible for JAK2V617F-induced ROS increase in this context and that the reduction of SOD2 expression may be a consequence of lowered ROS in the presence of JAK2 inhibitor (Supplementary Figure S3).

JAK2V617F reduces nuclear localization of FOXO3A. (a) JAK2 inhibitors increased the expression of catalase in HEL cells. (b) JAK2V617F knockdown in HEL cells increased the expression of catalase. (c) Catalase expression was reduced in Ba/F3-EpoR cells carrying JAK2V617F compared with wild-type JAK2. (d) JAK2V61F knockdown induced nuclear localization of FOXO3A. Representative images are shown. Nuclear FOXO3A is shown as a percentage of total FOXO3A. Numbers in parentheses indicate the number of cells analyzed for each sample. (e) JAK2V617F expression in BaF/3-EpoR cells increased cytoplasmic sequestration of FOXO3A compared with the wild-type JAK2. Data are mean±s.e.m. ***P<0.001.
Because catalase expression is regulated by FOXO transcription factors,1, 5 we next examined the possible role of JAK2V617F in modulating the transcriptional activity of FOXO3A, the abnormal repression or deletion of which has been linked to hematological malignancies.28, 29, 30 In HEL cells, ser253 phosphorylation of FOXO3A was reduced by inhibiting JAK2V617F (Supplementary Figure S4A), indicating that JAK2V617F signaling leads to FOXO3A phosphorylation. As the serine phosphorylation inactivates FOXO3A by excluding the protein from the nucleus and increasing its cytoplasmic sequestration,31 we investigated the sub-cellular localization of FOXO3A by immunofluorescence in HEL cells using an antibody to total FOXO3A. Nuclear localization of FOXO3A was significantly increased by two different JAK2 inhibitors (Supplementary Figure S4B) and also by siRNA-mediated JAK2 knockdown (Figure 3d). Furthermore, in Ba/F3-EpoR cells, nuclear localization of FOXO3A was reduced by the expression of JAK2V617F (Figure 3e) or by a JAK2 exon12 mutation, K539L (Supplementary Figure S5).
Taken together, these results demonstrate that JAK2V617F increases phosphorylation and cytoplasmic sequestration of FOXO3A, and that this is consequently associated with a reduction in catalase expression and an increase in intracellular ROS levels.
Inactivation of FOXO3A by JAK2V617F is mediated by PI3K signaling
To study the mechanism whereby FOXO3A is phosphorylated and inactivated downstream of JAK2V617F, we focused on the PI3K signaling pathway. This pathway is activated in JAK2V617F-positive cells32, 33, 34 and has been reported to phosphorylate FOXO transcription factors in various cell types.1, 28, 31 Treatment of HEL cells with either of two different PI3K inhibitors resulted in a significant increase in nuclear FOXO3A (Figure 4a). Inhibition of PI3K also increased catalase levels (Figure 4b), reduced intracellular ROS levels (Figure 4c) and increased etoposide-induced apoptosis (Figure 4d and e). To investigate whether the increased apoptosis by PI3K inhibition was due to reduced ROS, apoptosis was analyzed on cells treated with sodium azide, an irreversible inhibitor of catalase. The data indicate that an increase in etoposide-induced apoptosis by PI3K inhibition was reversed in the presence of sodium azide (Figure 4d and e), thus demonstrating that sensitization to DNA damage by PI3K inhibition is dependent on reduced ROS. Together, these data indicate that inactivation of FOXO3A by JAK2V617F is mediated by PI3K signaling.

Inactivation of FOXO3A is mediated by the PI3K signaling pathway. (a) Treatment with PI3K inhibitors relocalize FOXO3A to the nucleus in HEL cells. Representative pictures are shown on the left. Percentage of nuclear FOXO3A is shown on the right as an average of minimum 45 cells. (b) Inhibition of PI3K signalling increased catalase expression. (c) Inhibition of PI3K reduced ROS levels. (d and e) Inhibition of PI3K increased the sensitivity of HEL cells to DNA damage-induced apoptosis while the addition of sodium azide, an inhibitor of catalase, reversed this effect. All results are averages of three independent experiments and shown as mean±s.e.m. *P<0.05, **P<0.005 and ***P<0.0005.
Nuclear localization of FOXO3A is reduced in clonally derived JAK2V617F-positive erythroblasts from patients with a MPN
The results described above demonstrate that JAK2V617F reduces nuclear localization of FOXO3A and increases ROS in cell lines. We next sought to determine whether this is also true in primary cells from patients with JAK2V617F-positive MPNs. To this end, we studied 419 peripheral blood BFU-E colonies obtained from three patients with JAK2V617F-positive essential thrombocythemia or polycythemia vera. Individual colonies were picked, genotyped for JAK2 mutation status and analyzed for FOXO3A localization and intracellular ROS levels. As both wild-type and mutant colonies were grown under identical conditions, this experimental strategy allowed a direct comparison of JAK2 wild-type and JAK2V617F-positive erythroblasts from the same patient.
In each patient, cells carrying the JAK2V617F mutation showed a significant reduction in the nuclear localization of FOXO3A compared with those with wild-type JAK2 (Figure 5a and b). From one of the polycythemia vera patients, we obtained both JAK2V617F heterozygous and homozygous BFU-E colonies (Figure 5b, Patient 3) and observed a further reduction of nuclear FOXO3A in the homozygous cells compared with heterozygous cells. It seems that in this patient the strength of mutant JAK2 signaling may correlates with the extent of FOXO3A cytoplasmic sequestration. In addition, ROS levels analyzed in five MPN patients were significantly higher in the erythroblasts from mutant BFU-E colonies compared with those from wild-type colonies (Figure 5c). These results are consistent with our cell line data and demonstrate that, in erythroblasts from MPN patients, the JAK2V617F mutation is also associated with increased cytoplasmic sequestration of FOXO3A and elevated ROS levels.

JAK2V617F induces increased cytoplasmic sequestration of FOXO3A in erythroblasts from MPN patients. (a) JAK2V617F causes increased cytoplasmic sequestration of FOXO3A in BFU-E colonies derived from MPN patients. Representative images are shown. (b) Reduced FOXO3A nuclear localization in cells carrying JAK2V617F compared with the cells with normal JAK2. Graph shows the data obtained from each patient (Patient 1: essential thrombocythemia (ET), heterozygous for JAK2V617F; Patient 2: polycythemia vera (PV), heterozygous for JAK2V617F; Patient 3: PV, both heterozygous and homozygous BFU-E clones for JAK2V617F). Numbers in parentheses indicate the numbers of cells analyzed for each samples. Data are mean±s.e.m. **P<0.005, ***P<0.0005. (c) Mutant BFU-E colonies show increased ROS levels compared with the wild-type colonies. BFU-E colonies from five MPN patients were stained with DCF-DA. The data were normalized (WT=1) and relative fold increase is shown. A representative flow cytometry histogram is shown on the right. **P<0.005.
Nuclear localization of FOXO3A is not reduced by JAK2V617F in hematopoietic stem and early progenitor cells
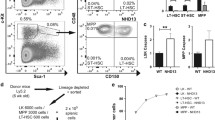
We next studied HSCs to determine whether JAK2V617F also results in cytoplasmic sequestration of FOXO3A in HSCs. To this end, we utilized JAK2V617F knock-in mice that carry human JAK2V617F and have a polycythemia vera-like phenotype.22 In accord with our data from human erythroblasts, levels of nuclear FOXO3A were reduced in JAK2 mutant progenitors (CD150−CD48+CD45+) compared with littermate controls (Figure 6a). By contrast, despite a mild increase in AKT phosphorylation (Supplementary Figure S6), the levels of nuclear FOXO3A were not altered in mutant HSCs from the same mice ((HSCs: CD150+CD48−EPCR+CD45+),35 Figure 6b).

Mutant JAK2-FOXO3A signaling is different in stem cells compared with the progenitors. (a) JAK2V617F induces an increase in the cytoplasmic sequestration of FOXO3A in the hematopoietic progenitors of JAK2V617F knock-in mice. (b) FOXO3A nuclear localization is not altered in the nuclei of primitive HSCs in both wild-type and JAK2V617F-positive mice. Representative images are shown in the left panel and bar graphs show the mean percentages of nuclear FOXO3A in the right panel. Numbers in parentheses indicate the numbers of cells analyzed from each mouse. (c) JAK2V617F induces an increase in the cytoplasmic sequestration of FOXO3A in the hematopoietic progenitors of MPN patients. WT1 and WT2: JAK2V617F-negative patients; VF1, VF2 and VF3: JAK2V617F-positive patients. Numbers in brackets indicate numbers of cells analyzed from each patient. (d) FOXO3A is in the nuclei of primitive HSCs in both JAK2V617F-negative and -positive patients. Data are mean±s.e.m. ***P<0.0005.
To confirm this finding in MPN patients, we isolated HSC-enriched population (HSCs; Lin−CD34+CD38−CD45RA−CD90+) and more mature progenitors (Lin−CD34+CD38+) from JAK2V617F-negative and JAK2V617F-positive patients with very high mutant allele burden. Nuclear levels of FOXO3A were reduced in more mature progenitors from JAK2 mutant patients compared with those from JAK2 wild-type MPN patients (Figure 6c). However, in HSCs isolated from the same patients, nuclear localization of FOXO3A was not altered by JAK2 genotypes (Figure 6d). Taken together, these results demonstrate that mutant JAK2 regulates nuclear localization of FOXO3A differently in primitive stem and progenitor cells, compared with more mature cells, with the latter showing a marked increase in the cytoplasmic sequestration of FOXO3A.
Discussion
JAK2V617F has previously been shown to increase the rate of DNA damage16, 17, 18, 19 and to inhibit DNA damage-induced apoptosis by repressing NHE-1 upregulation and Bcl-xL deamidation.20, 21 These attributes combine to provide a potent mechanism for the accumulation of DNA damage thought to drive the progression of JAK2-mutated MPNs. However, the mechanism whereby JAK2V617F blocks NHE-1 upregulation has remained unknown. In this paper, we demonstrate that JAK2V617F results in elevated intracellular ROS levels, which in turn inhibit the NHE-1/Bcl-xL pathway in response to DNA damage, thereby promoting cell survival. In addition, we show that the increased ROS levels associated with JAK2V617F reflect aberrant PI3K signaling and cytoplasmic sequestration of FOXO3A with consequent downregulation of catalase expression. However, FOXO3A nuclear localization is differentially modulated in more primitive hematopoietic stem and progenitor population.
Our data provide a mechanistic link between JAK2V617F and the inappropriate survival of cells harboring DNA damage (Figure 7). It is not clear how increased ROS levels repress NHE-1 expression, but one possibility is that a persistent pro-oxidative state, created by high ROS levels, inactivates the AP2 transcription factor, implicated in controlling NHE-1 expression,24, 25, 26 by modifying a thiol moiety in AP2 and thus inhibiting its DNA-binding activity.36 Our data do not exclude the possibility that repression of pro-apoptotic genes by FOXO3A inactivation also contributes to resistance to DNA damage-induced apoptosis. For instance, FOXO3A inactivation was reported to result in a transcriptional repression of pro-apoptotic BIM.29 Nevertheless, our data do demonstrate that increased catalase expression and a reduction in ROS sensitize cells to apoptosis in the presence of DNA damage.

Schematic pathway of JAK2V617F inhibition of the NHE-1/Bcl-xL-regulated apoptosis. Upper panel shows a normal signaling pathway of apoptosis in response to DNA damage. Lower panel shows the dysregulated pathway in JAK2V617F-positive cells. Dashed lines illustrate other unidentified factors or pathways that may potentially regulate ROS and resistance to DNA damage-induced apoptosis in JAK2V617F-positive cells.
The JAK2V617F mutation causes cytokine-independent activation of downstream signaling including the PI3K pathway;32, 33, 34 however, the contribution of activated PI3K signaling to MPN pathogenesis is unclear. Previously, PI3K signaling has been shown to inactivate FOXO3A,28, 31, 37 the main FOXO protein operating in hematopoietic cells.3, 4, 38 We demonstrate that JAK2 mutations compromise FOXO3A function by PI3K-dependent cytoplasmic sequestration in hematopoietic progenitors and clonally derived erythroblasts isolated from MPN patients as well as in cell lines. In addition, we show that JAK2V617F-induced inactivation of FOXO3A leads to reduced expression of catalase and, as a result, accumulation of intracellular ROS. Consistent with our findings, JAK2V617F knock-in mice were shown to have reduced catalase level and elevated ROS.39 However, it remains possible that increased ROS in JAK2V617F-positive cells may also be regulated by other pathways in addition to PI3K/FOXO3A.
ROS function as a 'double-edged sword' and the association of ROS with cancer is both complex and paradoxical. Although high levels of ROS that exceed the antioxidant capacity of the cell could trigger cell death, a moderate increase of ROS may promote cell survival. ROS levels are increased in many types of cancer cells and are associated with increased cell survival.40 Our data demonstrate that reduction of ROS sensitizes cells to apoptosis in response to DNA damage. Consistent with our finding, Lin et al.41 reported that antioxidant dihydromyricetin reduced ROS and promoted HepG2 cell apoptosis in a concentration-dependent manner. In addition, ROS are known to result in intrinsic DNA damage; increased DNA damage has previously been shown in cells from the JAK2V617F knock-in mice and MPN patients and in cell lines expressing JAK2V617F.16, 17, 18, 19, 42 These data indicate that the JAK2V617F-mediated increase in ROS has a dual role in promoting cell survival as well as allowing the accumulation of DNA damage. Furthermore, JAK2V617F has been reported to facilitate mutagenic DNA repair pathways,43 which, together with elevated ROS, would potentiate genome instability and disease progression. Indeed, 5–30% of MPN patients can progress to leukemia that is associated with additional mutations in genes such as TET2, DNMT3A and ASXL1.44
Our findings therefore raise the possibility that JAK2V617F-mediated dysregulation of FOXO3A is critical in the pathogenesis of human MPNs. In accord with these data, FOXO3A−/− mice were reported to exhibit a myeloproliferative phenotype.30 Our results also shed light on whether dysregulation of FOXO3A and increased ROS levels represent early or late events during tumorigenesis. MPNs are detected early, frequently following an incidental blood count, and provide a window on the earliest stages of malignancy that are inaccessible in most cancers. In particular, MPNs have not needed to acquire mutations associated with many aspects of the malignant phenotype (for example, differentiation block, tissue invasion, growth in hypoxic environment). Our data therefore indicate that FOXO3A inactivation occurs early in the evolution of myeloid malignancies, an observation with implications for other cancers in which FOXO proteins are dysregulated.6
Our findings have several therapeutic implications for the treatment of MPNs and other malignancies. The addition of PI3K inhibitors significantly reduces the survival of JAK2V617F-positive cells following DNA damage, and reverses the altered ROS levels and FOXO3A localization. These data indicate that the PI3K pathway is critical for maintaining resistance to apoptosis in malignant cells. Furthermore, we found that counteracting the JAK2V617F-induced increase in ROS sensitizes the JAK2-mutant cells to a DNA damage-inducing agent. This is reminiscent of findings from other cancers where the combination of an antioxidant with chemotherapeutic interventions has been shown to be effective in killing drug-resistant tumors.11, 12, 13 Malignant cells therefore seem to be sensitive to perturbation of the oncogene-mediated altered redox state, suggesting that these cells have adapted to and become dependent on increased ROS. Because increased activation of the PI3K pathway and elevated ROS levels are features common to many cancers, these findings may be relevant well beyond hematological malignancies.
FOXO3A inactivation has been previously reported in other hematological malignancies such as BCR-ABL-positive CML. Similar to JAK2V617F, BCR-ABL activates PI3K pathway which in turn inactivates FOXO3A.28, 45 BCR-ABL-specific tyrosine kinase inhibitor treatment led to rapid cytogenetic response in the majority of CML patients in the chronic phase,46 and this is associated with FOXO3A reactivation.47 However, numerous studies have demonstrated that tyrosine kinase inhibitors cannot fully eliminate BCR-ABL transcripts and eradicate disease.48, 49, 50 The mechanisms for tyrosine kinase inhibitor resistance of CML stem and progenitor cells are not entirely clear, and it has been previously reported that primitive CML cells do not depend on BCR-ABL kinase activity for survival.51, 52 One line of evidence suggests that the anti-proliferative effects of tyrosine kinase inhibitors lead to an increase in quiescent CML stem and progenitor cells, making them difficult to target. Importantly, this effect was shown to be mediated by FOXO transcription factors.47 Moreover, Naka et al.53 demonstrated that FOXO3A is predominantly localized in the nuclei of CML leukemia-initiating cells and that deletion of FOXO3a compromised the ability of leukemia-initiating cells to initiate the disease in transplant recipients. Together, these findings suggest that the function of FOXO transcription factors seem to depend largely on the context in which they are expressed. Consistent with this notion, JAK2 inhibitors also fail to eradicate primitive HSCs that carry the JAK2V617F mutation54 and previous evidence has shown that JAK2V617F has distinct effects on the maintenance and proliferation of HSCs compared with progenitors.42 As demonstrated in this study, FOXO3A is mainly localized in the nuclei of JAK2V617F-positive primitive HSCs. Nuclear FOXO3A in mutant HSCs from MPN patients could potentially be responsible for drug resistance,54 maintenance of quiescence5 and/or alternatively, induction of protective autophagy under conditions of stress,55 or efficient DNA repair possibly by interacting with ATM.56 Thus, our data suggest that MPN HSCs may be resistant to cellular stress in a FOXO-dependent manner. Patients with chronic phase MPNs lead normal lives, whereas accelerated phase disease is associated with a marked increase in morbidity and mortality. Prevention of disease evolution is therefore important and this study suggests that combinational therapies specific to HSCs together with JAK2 inhibitors are required to effectively eradicate the disease.
Materials and methods
Reagents
AT9283 and TG101209 were synthesized at Astex Therapeutics (Cambridge, UK). LY294002 was purchased from Cell Signaling (Hitchin, UK) and PI103 from Symansis (Buckingham, UK). MitoQ was kindly provided by Michael Murphy (MRC Mitochondrial Biology Unit, Cambridge, UK). All other reagents were purchased from Sigma (Poole, UK) unless otherwise stated.
Cell lines
HEL cells were cultured in RPMI supplemented with 10% fetal calf serum. HEL cells were treated with DMSO, 1 μM AT9283, or 3 μM TG101209 for 4 h for JAK2V617F inhibition. Ba/F3-EpoR cells expressing wild-type or mutant JAK2 were maintained in RPMI with 10% fetal calf serum, 1 U/ml erythropoietin (R&D Systems, Abingdon, UK) and 0.5 mg/ml G418.
Mice
JAK2V617F knock-in mice were previously generated and reported.22 All mice were kept in specific pathogen-free conditions and all procedures were performed according to rules of the Animal Welfare and Ethical Review Body (AWERB) and the UK Home Office regulations.
MPN patients
MPN patients diagnosed with either essential thrombocythemia or polycythemia vera according to the World Health Organisation (WHO) criteria57 were recruited from the MPN clinic at the Addenbrooke's Hospital in Cambridge, UK. Informed consent was given from all patient subjects. The work involving patient samples was approved by the Cambridge and Eastern Region Ethics Committee and was performed in accordance with the principles of the Declaration of Helsinki.
Analysis of BFU-E colonies
Venous blood samples (40 ml) were collected from each patient and peripheral blood mononuclear cells were isolated using Lymphoprep (Axis Shield, Kimbolton, UK) according to the manufacturer's instructions. Peripheral blood mononuclear cells (3 × 105 cells/ml) were then plated in methocult (H4531; Stem Cell Technologies, Grenoble, France) supplemented with 0.01 U/ml erythropoietin and cultured for 14 days at 37 °C. Individual BFU-E colonies were then plucked and resuspended in 30 μl phosphate-buffered saline (PBS), of which 2 μl were used for genotyping by real-time qPCR to determine the JAK2 mutational status.58 Colonies with JAK2V617F percentages of less than 10% were designated 'wild-type', those between 40–60% 'heterozygous' and those greater than 90% 'homozygous'.
Stem and progenitor cell isolation from MPN patients
Peripheral blood mononuclear cells from MPN patients were washed with PBS supplemented with 2% fetal bovine serum and 5 mM EDTA and stored overnight at 4 °C in PBS supplemented with 10% fetal bovine serum and 5 mM EDTA. Cells were then washed with PBS with 2% fetal bovine serum and 5 mM EDTA and spun at 300 g for 7 min, and the resulting pellets were treated with 50 ul of 1 mg/ml DNase I solution (STEMCELL Technologies, Cambridge, UK) to avoid clumping. Cells were then depleted of differentiated hematopoietic cells using the EasySep Human Hematopoietic Progenitor enrichment kit (STEMCELL Technologies) with the following modifications: All samples were processed in 500 μl of recommended medium for the first step, the progenitor enrichment cocktail was used at 5 μl/ml cells, and the sample was only taken through one round of separation. Cells were then stained with antibodies to isolate the various progenitor compartments: anti-CD34 Per-CpCy5.5 (clone 581 BioLegend, London, UK), anti-CD38 FITC (clone HIT2, BD Biosciences), anti-CD90 APC (clone 5E10, BD Biosciences, Oxford, UK), anti-FLK2 PE (clone BV10A4H2, BioLegend), anti-CD10 APC-Cy7 (clone HI10a, BioLegend) and anti-CD45RA Horizon V450 (HI100, BD Biosciences). HSCs were isolated as Lin−CD34+CD38−CD45RA−CD90+, progenitors as Lin−CD34+CD38+. Cells were sorted using a BD Influx sorter into tubes containing PBS supplemented with 10% fetal bovine serum and EDTA.
JAK2 knockdown
HEL cells were transfected with either human JAK2 ON-TARGETplus SMARTpool or ON-TARGETplus Non-targeting Pool (control) (Dharmacon RNAi Technologies, Little Chalfont, UK). In detail, 5 ul 20 uM RNAi (either JAK2 or control) was mixed in 500 ul opti-mem (Invitrogen, Paisley, UK) in tube A, and 5 ul Lipofectamine was mixed in 500 ul opti-mem (Invitrogen) in tube B. Both tubes were incubated at room temperature for 10 min. Then the mixture of tube A and B were left at room temperature for 30 min and added to 2 ml HEL cells (1 × 106 cells in six-well plate). Cells were collected 48 h later for analysis.
Transfection and retroviral transduction
MSCV-neo-based retroviral vectors MSCV-JAK2(WT)-neo, MSCV-JAK2V617F-neo and MSCV-JAK2K539L-neo were transfected into 293T cells using FuGENE 6 Transfection Reagent (Roche Applied Science, Burgess Hill, UK) and culture supernatants were harvested 48 h later. Viral infection of Ba/F3-EpoR cells was done by spinoculation (2000 r.p.m. for 2 h at 30 °C) and the cells were cultured in RPMI with 10% fetal calf serum and 1 U/ml erythropoietin. G418 was added 2 days after infection to select for the transduced cells.
Immunofluorescence
Cytospin slides were fixed and permeabilized in 1:1 mixture of methanol and acetone. The slides were blocked for 1 h at room temperature and then stained overnight at 4 °C in 0.2% gelatine in PBS using primary antibody against FOXO3A (Cell Signaling, 2497S). For immunostaining of pAKT (Cell Signaling, 4060S), the slides were blocked for 1 h at room temperature in PBS/5% normal serum/0.3% Triton X-100) and then stained overnight at 4 °C in PBS/1% bovine serum albumin/0.3% Triton X-100. The secondary Alexa Fluor 555-conjugated antibody (Life Technologies, Paisley, UK; A-21428) was applied at room temperature for 1 h in the respective primary antibody staining buffer. Nuclei were stained with 4', 6-diamidino-2-phenylindole (Vectashield, Burlingame, CA, USA) and images were captured and scored blindly using ZEISS Axioskop2 mot plus fluorescence microscope equipped with a × 40 Plan NEOFLUAR lens and an AxioCam MRm camera (ZEISS, Cambridge, UK). Images were analyzed using Isis software (Oxford Isis, Oxford, UK).
Real-time quantitative PCR
RNA was extracted using Tri reagent (Sigma) and real-time quantitative PCR was performed for human NHE1 expression using qPCR SYBR Green (Agilent Technologies UK Ltd., Workingham, UK) and Mx3000P Real-Time PCR system (Stratagene, La Jolla, CA, USA). The forward and reverse primers were 5′-TTGTTTTTGGGGAGTCCTTG-3′ and 5′-ATGCCCACGTGTTCGTAGTT-3′. Results were calculated using the delta-delta-Ct method with ABL as a control. The forward and reverse primers for ABL are 5′-GCGTGAGAGTGAGAGCAG-3′ and 5′-TCTCGGAGGAGACGTAG-3′. NHE1 expression in each sample was normalized to ABL expression and the relative NHE1 expression in bar graphs represent the relative levels to control which was normalized to 1.
Analyses of ROS and apoptosis
For ROS measurement, cells were stained with 10 μM 2',7'-dichlorodihydrofluorescein diacetate (H2DCF-DA, Invitrogen). For Ba/F3-EpoR cells, cells were stained with 5 μM dihydrorhodamine 6G (Invitrogen) for 30–45 min at 37 °C. Cells were then washed in PBS and the fluorescent signals were analyzed by fluorescence-activated cell sorting. To analyze apoptosis, cells were stained with Annexin V antibody (BioLegend) and 7-aminoactinomycin D (Invitrogen) in Annexin V binding buffer (BD Biosciences).
Western blot analyses
Western blots on total cell lysates were performed using the following antibodies: anti-FOXO3A (Cell Signaling, 2497 S), phospho-FOXO3A (Ser319) (Cell Signaling, 9466 S), STAT5 (Santa Cruz, Dallas, TX, USA; SC-835), phospho-STAT5 (Tyr694) (Cell Signaling, 9351 S), Catalase (Abcam, ab16731), Bcl-xL (BD Biosciences, 610212), anti-SOD2 (Life Technologies, MA1-106), anti-Glutathione reductase (Abcam, Cambridge, UK; ab128933), β-actin (Sigma, A2228) and α-Tubulin (Sigma, T9026).
Fluorescence-activated cell sorting of mouse HSCs and progenitor populations
Mouse HSCs and progenitors were obtained from JAK2V617F knock-in mice22 and their littermate controls (8–10-weeks old, age- and sex-matched mice in C57BL/6 background) by fluorescence-activated cell sorting of bone marrow MNCs. E-SLAM HSCs were isolated as previously described42 with HSCs as CD150+CD48-EPCR+CD45+ and mature progenitors as CD150−CD48+CD45+.
Statistics
The statistical significance was determined by a two-tailed, unpaired Student t-test.
References
Calnan DR, Brunet A . The FoxO code. Oncogene 2008; 27: 2276–2288.
Bakker WJ, van Dijk TB, Parren-van Amelsvoort M, Kolbus A, Yamamoto K, Steinlein P et al. Differential regulation of Foxo3a target genes in erythropoiesis. Mol Cell Biol 2007; 27: 3839–3854.
Miyamoto K, Araki KY, Naka K, Takubo K, Yamazaki S, Matsuoka S et al. Foxo3a is essential for maintenance of the hematopoietic stem cell pool. Cell Stem Cell 2007; 1: 101–112.
Miyamoto K, Miyamoto T, Kato R, Yoshimura A, Motoyama N, Suda T . FoxO3a regulates hematopoietic homeostasis through a negative feedback pathway in conditions of stress or aging. Blood 2008; 112: 4485–4493.
Tothova Z, Gilliland DG . FoxO transcription factors and stem cell homeostasis: insights from the hematopoietic system. Cell Stem Cell 2007; 1: 140–152.
Fu Z, Tindall DJ . FOXOs cancer and regulation of apoptosis. Oncogene 2008; 27: 2312–2319.
Pourova J, Kottova M, Voprsalova M, Pour M . Reactive oxygen and nitrogen species in normal physiological processes. Acta Physiol (Oxf) 198: 15–35.
Weinberg F, Hamanaka R, Wheaton WW, Weinberg S, Joseph J, Lopez M et al. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc Natl Acad Sci USA 107: 8788–8793.
Naughton R, Quiney C, Turner SD, Cotter TG . Bcr-Abl-mediated redox regulation of the PI3K/AKT pathway. Leukemia 2009; 23: 1432–1440.
Chan DW, Liu VW, Tsao GS, Yao KM, Furukawa T, Chan KK et al. Loss of MKP3 mediated by oxidative stress enhances tumorigenicity and chemoresistance of ovarian cancer cells. Carcinogenesis 2008; 29: 1742–1750.
Rakshit S, Bagchi J, Mandal L, Paul K, Ganguly D, Bhattacharjee S et al. N-acetyl cysteine enhances imatinib-induced apoptosis of Bcr-Abl+ cells by endothelial nitric oxide synthase-mediated production of nitric oxide. Apoptosis 2009; 14: 298–308.
Sikora MJ, Bauer JA, Verhaegen M, Belbin TJ, Prystowsky MB, Taylor JC et al. Anti-oxidant treatment enhances anti-tumor cytotoxicity of (-)-gossypol. Cancer Biol Ther 2008; 7: 767–776.
Tsai SY, Sun NK, Lu HP, Cheng ML, Chao CC . Involvement of reactive oxygen species in multidrug resistance of a vincristine-selected lymphoblastoma. Cancer Sci 2007; 98: 1206–1214.
Sedelnikova OA, Redon CE, Dickey JS, Nakamura AJ, Georgakilas AG, Bonner WM . Role of oxidatively induced DNA lesions in human pathogenesis. Mutat Res 2010; 704: 152–159.
Campbell PJ, Green AR . The myeloproliferative disorders. N Engl J Med 2006; 355: 2452–2466.
Plo I, Nakatake M, Malivert L, de Villartay JP, Giraudier S, Villeval JL et al. JAK2 stimulates homologous recombination and genetic instability: potential implication in the heterogeneity of myeloproliferative disorders. Blood 2008; 112: 1402–1412.
Li J, Spensberger D, Ahn JS, Anand S, Beer PA, Ghevaert C et al. JAK2 V617F impairs hematopoietic stem cell function in a conditional knock-in mouse model of JAK2 V617F-positive essential thrombocythemia. Blood 2010; 116: 1528–1538.
Chen E, Ahn JS, Massie CE, Clynes D, Godfrey AL, Li J et al. JAK2V617F promotes replication fork stalling with disease-restricted impairment of the intra-S checkpoint response. Proc Natl Acad Sci USA 2014; 111: 15190–15195.
Kagoya Y, Yoshimi A, Tsuruta-Kishino T, Arai S, Satoh T, Akira S et al. JAK2V617F+ myeloproliferative neoplasm clones evoke paracrine DNA damage to adjacent normal cells through secretion of lipocalin-2. Blood 2014; 124: 2996–3006.
Zhao R, Oxley D, Smith TS, Follows GA, Green AR, Alexander DR . DNA damage-induced Bcl-xL deamidation is mediated by NHE-1 antiport regulated intracellular pH. PLoS Biol 2007; 5: e1.
Zhao R, Follows GA, Beer PA, Scott LM, Huntly BJ, Green AR et al. Inhibition of the Bcl-xL deamidation pathway in myeloproliferative disorders. N Engl J Med 2008; 359: 2778–2789.
Li J, Kent DG, Godfrey AL, Manning H, Nangalia J, Aziz A et al. JAK2V617F homozygosity drives a phenotypic switch in myeloproliferative neoplasms, but is insufficient to sustain disease. Blood 2014; 123: 3139–3151.
Quentmeier H, MacLeod RA, Zaborski M, Drexler HG . JAK2 V617F tyrosine kinase mutation in cell lines derived from myeloproliferative disorders. Leukemia 2006; 20: 471–476.
Kumar AP, Chang MK, Fliegel L, Pervaiz S, Clement MV . Oxidative repression of NHE1 gene expression involves iron-mediated caspase activity. Cell Death Differ 2007; 14: 1733–1746.
Akram S, Teong HF, Fliegel L, Pervaiz S, Clement MV . Reactive oxygen species-mediated regulation of the Na+-H+ exchanger 1 gene expression connects intracellular redox status with cells' sensitivity to death triggers. Cell Death Differ 2006; 13: 628–641.
Jhumka Z, Pervaiz S, Clement MV . Resveratrol regulates the expression of NHE-1 by repressing its promoter activity: critical involvement of intracellular H2O2 and caspases 3 and 6 in the absence of cell death. Int J Biochem Cell Biol 2009; 41: 945–956.
Gaetani GF, Ferraris AM, Rolfo M, Mangerini R, Arena S, Kirkman HN . Predominant role of catalase in the disposal of hydrogen peroxide within human erythrocytes. Blood 1996; 87: 1595–1599.
Komatsu N, Watanabe T, Uchida M, Mori M, Kirito K, Kikuchi S et al. A member of Forkhead transcription factor FKHRL1 is a downstream effector of STI571-induced cell cycle arrest in BCR-ABL-expressing cells. J Biol Chem 2003; 278: 6411–6419.
Essafi A, Fernandez de Mattos S, Hassen YA, Soeiro I, Mufti GJ, Thomas NS et al. Direct transcriptional regulation of Bim by FoxO3a mediates STI571-induced apoptosis in Bcr-Abl-expressing cells. Oncogene 2005; 24: 2317–2329.
Yalcin S, Marinkovic D, Mungamuri SK, Zhang X, Tong W, Sellers R et al. ROS-mediated amplification of AKT/mTOR signalling pathway leads to myeloproliferative syndrome in Foxo3(-/-) mice. Embo J 2010; 29: 4118–4131.
Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS et al. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999; 96: 857–868.
James C, Ugo V, Le Couedic JP, Staerk J, Delhommeau F, Lacout C et al. A unique clonal JAK2 mutation leading to constitutive signalling causes polycythaemia vera. Nature 2005; 434: 1144–1148.
Levine RL, Wadleigh M, Cools J, Ebert BL, Wernig G, Huntly BJ et al. Activating mutation in the tyrosine kinase JAK2 in polycythemia vera, essential thrombocythemia, and myeloid metaplasia with myelofibrosis. Cancer Cell 2005; 7: 387–397.
Kralovics R, Passamonti F, Teo SS, Brutsche M, Tiedt R, Tichelli A et al. A gain of function mutation in Jak2 is frequently found in patients with myeloproliferative disorders. N Engl J Med 2005; 352: 1779–1790.
Kent DG, Copley MR, Benz C, Wöhrer S, Dykstra BJ, Ma E et al. Prospective isolation and molecular characterization of hematopoietic stem cells with durable self-renewal potential. Blood 2009; 113: 6342–6350.
Huang Y, Domann FE . Redox modulation of AP-2 DNA binding activity in vitro. Biochem Biophys Res Commun 1998; 249: 307–312.
You H, Pellegrini M, Tsuchihara K, Yamamoto K, Hacker G, Erlacher M et al. FOXO3a-dependent regulation of Puma in response to cytokine/growth factor withdrawal. J Exp Med 2006; 203: 1657–1663.
Yalcin S, Zhang X, Luciano JP, Mungamuri SK, Marinkovic D, Vercherat C et al. Foxo3 is essential for the regulation of ataxia telangiectasia mutated and oxidative stress-mediated homeostasis of hematopoietic stem cells. J Biol Chem 2008; 283: 25692–25705.
Marty C, Lacout C, Droin N, Le Couédic JP, Ribrag V, Solary E et al. A role for reactive oxygen species in JAK2 V617F myeloproliferative neoplasm progression. Leukemia 2013; 27: 2187–2195.
Trachootham D, Alexandre J, Huang P . Targeting cancer cells by ROS-mediated mechanisms: a radical therapeutic approach? Nat Rev Drug Discov 2009; 8: 579–591.
Lin B, Tan X, Liang J, Wu S, Liu J, Zhang Q et al. A reduction in reactive oxygen species contributes to dihydromyricetin-induced apoptosis in human hepatocellular carcinoma cells. Sci Rep 2014; 4: 7041.
Kent DG, Li J, Tanna H, Fink J, Kirschner K, Pask DC et al. Self-renewal of single mouse hematopoietic stem cells is reduced by JAK2V617F without compromising progenitor cell expansion. PLos Biol 2013; 11: e1001576.
Fernandes MS, Reddy MM, Gonneville JR, DeRoo SC, Podar K, Griffin JD et al. BCR-ABL promotes the frequency of mutagenic single-strand annealing DNA repair. Blood 2009; 114: 1813–1819.
Vainchenker W, Delhommeau F, Constantinescu SN, Bernard OA . New mutations and pathogenesis of myeloproliferative neoplasms. Blood 2011; 118: 1723–1735.
Ghaffari S, Jagani Z, Kitidis C, Lodish HF, Khosravi-Far R . Cytokines and BCR-ABL mediate suppression of TRAIL-induced apoptosis through inhibition of forkhead FOXO3a transcription factor. Proc Natl Acad Sci USA 2003; 100: 6523–6528.
Druker BJ, Guilhot F, O'Brien SG, Gathmann I, Kantarjian H, Gattermann N et al. Five-year follow-up of patients receiving imatinib for chronic myeloid leukemia. N Engl J Med 2006; 355: 2408–2417.
Pellicano F, Scott MT, Helgason GV, Hopcroft LE, Allan EK, Aspinall-O'Dea M et al. The antiproliferative activity of kinase inhibitors in chronic myeloid leukemia cells is mediated by FOXO transcription factors. Stem Cells 2014; 32: 2324–2337.
Hughes TP, Kaeda J, Branford S, Rudzki Z, Hochhaus A, Hensley ML et al. Frequency of major molecular responses to imatinib or interferon alfa plus cytarabine in newly diagnosed chronic myeloid leukemia. N Engl J Med 2003; 349: 1423–1432.
Chu S, McDonald T, Lin A, Chakraborty S, Huang Q, Snyder DS et al. Persistence of leukemia stem cells in chronic myelogenous leukemia patients in prolonged remission with imatinib treatment. Blood 2011; 118: 5565–5572.
Chomel JC, Bonnet ML, Sorel N, Bertrand A, Meunier MC, Fichelson S et al. Leukemic stem cell persistence in chronic myeloid leukemia patients with sustained undetectable molecular residual disease. Blood 2011; 118: 3657–3660.
Corbin AS, Agarwal A, Loriaux M, Cortes J, Deininger MW, Druker BJ . Human chronic myeloid leukemia stem cells are insensitive to imatinib despite inhibition of BCR-ABL activity. J Clin Invest 2011; 121: 396–409.
Hamilton A, Helgason GV, Schemionek M, Zhang B, Myssina S, Allan EK et al. Chronic myeloid leukemia stem cells are not dependent on Bcr-Abl kinase activity for their survival. Blood 2012; 119: 1501–1510.
Naka K, Hoshii T, Muraguchi T, Tadokoro Y, Ooshio T, Kondo Y et al. TGF-beta-FOXO signalling maintains leukaemia-initiating cells in chronic myeloid leukaemia. Nature 2010; 463: 676–680.
Mullally A, Lane SW, Ball B, Megerdichian C, Okabe R, Al-Shahrour F et al. Physiological Jak2V617F expression causes a lethal myeloproliferative neoplasm with differential effects on hematopoietic stem and progenitor cells. Cancer Cell 2010; 17: 584–596.
Imai Y, Takahashi A, Hanyu A, Hori S, Sato S, Naka K et al. Crosstalk between the Rb pathway and AKT signaling forms a quiescence-senescence switch. Cell Rep 2014; 7: 194–207.
Tsai WB, Chung YM, Takahashi Y, Xu Z, Hu MC . Functional interaction between FOXO3a and ATM regulates DNA damage response. Nat Cell Biol 2008; 10: 460–467.
Swerdlow SH, Campo E, Harris NL, Jaffe ES, Pileri SA, Stein H . WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues. IARC Press: Lyon, 2008.
Levine RL, Belisle C, Wadleigh M, Zahrieh D, Lee S, Chagnon P et al. X-inactivation-based clonality analysis and quantitative JAK2V617F assessment reveal a strong association between clonality and JAK2V617F in PV but not ET/MMM, and identifies a subset of JAK2V617F-negative ET and MMM patients with clonal hematopoiesis. Blood 2006; 107: 4139–4141.
Acknowledgements
We thank patients and staff of the Addenbrookes MPN clinic who have contributed samples to this study; Yvonne Silber for technical assistance; Philip Beer, Anna Godfrey, Christina Ortmann and Joanna Baxter (Translational Research Laboratory, University of Cambridge) for help with acquiring clinical samples; Peter Campbell (Sanger Institute) for helpful discussions; and Michael Murphy (MRC MBU) for supplying MitoQ. Work in the Green lab is supported by Leukemia and Lymphoma Research, Cancer Research UK, the Kay Kendall Leukaemia Fund, the NIHR Cambridge Biomedical Research Centre, the Cambridge Experimental Cancer Medicine Centre and the Leukemia & Lymphoma Society of America. DGK was supported by a postdoctoral fellowship from the Canadian Institutes of Health Research (Ottawa, ON), and a Lady Tata Memorial Trust International Award for Research in Leukaemia (London, UK). HJP was supported by a postdoctoral fellowship from the Human Frontier Science Program.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Supplementary information
Rights and permissions
About this article

Cite this article
Ahn, J., Li, J., Chen, E. et al. JAK2V617F mediates resistance to DNA damage-induced apoptosis by modulating FOXO3A localization and Bcl-xL deamidation. Oncogene 35, 2235–2246 (2016). https://doi.org/10.1038/onc.2015.285
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2015.285
- Springer Nature Limited
This article is cited by
-
Hypoxia-inducible factor 1 (HIF-1) is a new therapeutic target in JAK2V617F-positive myeloproliferative neoplasms
Leukemia (2020)
-
The ruxolitinib effect: understanding how molecular pathogenesis and epigenetic dysregulation impact therapeutic efficacy in myeloproliferative neoplasms
Journal of Translational Medicine (2018)
-
Amiodarone-Induced Retinal Neuronal Cell Apoptosis Attenuated by IGF-1 via Counter Regulation of the PI3k/Akt/FoxO3a Pathway
Molecular Neurobiology (2017)