
Abstract
The main genetic factor which initiates the flowering is the Vrn gene system, which determines the rate of many of the plant’s growth and development processes. Introgression of new Vrn gene alleles from its relatives into bread wheat makes it possible to increase genetic variables connected to the duration of the growing season parameters and individual developmental phases. Two lines of the winter cultivar of bread wheat Bezostaya 1 (Bez1) with a combination of the dominant alleles Vrn-A1L Vrn-B1a and Vrn-A1L Vrn-B1c were created which included the introgression of the Vrn-A1L (or Vrn-A1c Langdon-type deletion) allele from Triticum petropavlovskyi Udacz. et Migusch. (or T. aestivum ssp. petropavlovskyi (Udacz. et Migusch.) N.P. Gontsch). Homozygous lines were isolated from F3 hybrids by using marker-assisted selection. This lines matured earlier in relation to the original near-isogenic lines which contained the Vrn-A1L, Vrn-B1a, and Vrn-B1c alleles. The Bez1 Vrn-A1L Vrn-B1c line had a shorter germination-first node and germination-heading periods compared to Bez1 Vrn-A1L Vrn-B1a, practically showing no difference, in terms of heading, with the early-maturing line i:Bez1 Vrn-A1a. In the current paper the results of research into the productivity of the lines using different combinations of VRN-1 alleles are presented. Thus, the obtained results indicate the possibility of using the Vrn-A1L allele carrying out modification for earlier maturity arising as a result of combinations with other dominant Vrn-B1 alleles.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The adaptability of wheat to a wide range of environment conditions is mainly controlled by genes, determining the vernalization requirement (Vrn) and photoperiod sensitivity (Ppd). The vernalization pathway is responsible for the prevention of flowering before or during periods of low temperature which are detrimental to the apical meristem of the growth apex. Sensitivity to the photoperiod (day length) in wheat is controlled by the PPD-1 genes: Ppd-D1, Ppd-B1, and Ppd-A1, localized in the 2D, 2B, and 2A chromosomes, respectively, and allelic differences between dominant and recessive alleles are determined by deletions or insertions in the promoter region (Beales et al. 2007; Nishida et al. 2013). PPD-1 is the main activator of TaFT1 (VRN-3).
The response to vernalization is controlled by VRN-1, VRN-2, VRN-3, and VRN-4 genes. VRN-1 encode MADS-box proteins with high similarity to Arabidopsis thaliana meristem identity APETALA1. VRN-1 are three orthologous genes: Vrn-A1, Vrn-B1, and Vrn-D1, located in the 5A, 5B, and 5D chromosomes, respectively (Yan et al. 2003). The presence of at least one VRN-1 dominant allele determines the spring growth habit in wheat. Plants with Vrn-A1 mature earlier than plants with Vrn-B1 or Vrn-D1 (Goncharov 2004), which correlates with the relative level of expression of these genes (Loukoianov et al. 2005). VRN-1 is dominant for spring growth habit, whereas VRN-2 is dominant for winter growth habit (Yan et al. 2004b). Although VRN-3 in wheat was originally identified as a vernalization gene, the VRN-3 (TaFT1) gene is an ortholog of the Arabidopsis FT gene, and an integrator of various pathways, involved in the determination of flowering time (Yan et al. 2006). Vrn-D4 is a VRN-1 paralog on chromosome 5DS (Kippes et al. 2015). In a simplified form, the process of the interaction of VRN genes looks like this: prior to the onset of vernalization, the expression level of VRN-1 and VRN-3 is low, which prevents plants from progressing to the stage of generative development. Due to the effects brought about by the influence of vernalization, VRN-1 expression increases in the cells of the apical meristem, which when reaching a certain threshold value, initiates its transition from the vegetative to the generative stage (Loukoianov et al. 2005). In leaves, the product of the VRN-1 gene suppresses transcription of the flowering repressor gene VRN-2, which in turn suppresses transcription of VRN-3. The product of the VRN-3 gene, whose expression is induced by PPD-1, binds to the promoter region of the VRN-1 gene in apical meristem cells. VRN-1 expression is then activated, which leads to the formation of a positive feedback loop, and thus the initiation of flowering (Chen and Dubcovsky 2012).
A polymorphism in the promoter and the first intron of VRN-1 genes causes the known allelic diversity of wheat in heading time. The most common allele of the Vrn-A1 locus in bread wheat, Vrn-A1a, contain a 222 bp insertion and a duplication in the promoter (Yan et al. 2004a). In addition to Vrn-A1a, several other alleles were described in bread wheat: Vrn-A1b (Vrn-A1b.1) and Vrn-A1c with a 20 bp and 5.5 kb deletion in the promoter (Yan et al. 2004a) and in the first intron (Fu et al. 2005), respectively. Zhang et al. (2022) identified a new dominant Vrn-A1o and resessive vrn-A1n alleles with 54 bp and 11 bp deletions in the promoter region, respectively. In the first intron of bread wheat, large deletions are described, that distinguish the dominant alleles Vrn-B1a (6.8 kb deletion) (Fu et al. 2005), Vrn-B1b (6.8 kb and 36 bp deletions and SNP) (Santra et al. 2009), Vrn-B1c (7.6 kb deletion and 400 bp duplication) (Milec et al. 2012; Shcherban et al. 2012b), and Vrn-B1d (6.8 kb and 187 bp deletions, 4 bp mutation and SNP) (Zhang et al. 2018) from the recessive ones. A 4.2 kb deletion and 0.8 kb insertion in the first intron is characteristic of the Vrn-D1a and Vrn-D1s alleles, respectively (Fu et al. 2005; Muterko et al. 2015). Vrn-D1b and Vrn-D1c are characterized an SNP and a 174 bp insertion in the promoter region, respectively (Zhang et al. 2012, 2015).
The influence of different mutations of the VRN-1 locus on the timing of the onset of flowering and heading of bread wheat has been established. The most early to mature are the genotypes carrying the Vrn-A1a. Genotypes carrying Vrn-B1c, in turn, are faster to mature compared to carriers of Vrn-B1a (Emtseva et al. 2013). Plants whose development type is controlled by more than one dominant allele (Vrn-A1a, Vrn-B1a, Vrn-B1c or Vrn-D1a) tend to be faster maturing than those containing only one allele (Kiss et al. 2014; Efremova et al. 2016).
The determined distribution of allelic variants of Vrn and Ppd differs by geographical region, and different ecological and geographical zones, with different combinations of alleles possessing an advantage and providing a wide level of ecological plasticity in bread wheat (Shcherban et al. 2012a, 2015; Milec et al. 2013; Zhang et al. 2015, 2022; Efremova et al. 2016; Whittal et al. 2018; Shi et al. 2019). The majority of spring cultivars in Russia (including Western Siberia), as well as a number of temperate regions, is characterized by the presence of the two dominant genes Vrn-A1 (Vrn-A1a) and Vrn-B1 (Vrn-B1a, Vrn-B1c) which are faster in maturing and more productive than cultivars with a single Vrn gene (Potokina et al. 2012; Likhenko et al. 2015; Efremova et al. 2016; Smolenskaya et al. 2022).
To increase the genetic diversity of common wheat, dominant alleles found in the Vrn genes of its wild relatives can be introgressed into bread wheat genome (Stelmakh and Avsenin 1996; Zhang et al. 2008; Ivaničová et al. 2016). We obtained the near-isogenic line (NIL) of the winter cultivar Bezostaya 1 (Bez1) with the dominant Vrn-A1L allele (or Vrn-A1c Langdon-type deletion) introgressed from hexaploid wheat Triticum petropavlovskyi (Udacz. et Migusch.) (or T. aestivum ssp. petropavlovskyi (Udacz. et Migusch.) N.P. Gontsch), the effect of which is connected to a significant increase in heading time when compared to the Vrn-A1a allele (Chumanova et al. 2023). Vrn-A1L occurs in the tetraploid wheat species (Shcherban and Salina 2017; Shi et al. 2019). However, as mentioned above, modern cultivars of spring bread wheat in the world mainly carry alleles of two dominant genes, Vrn-A1 and Vrn-B1. It would therefore be of interest to study the phenotypic effects of the interaction of the Vrn-A1L allele with common alleles of the Vrn-B1 locus (Vrn-B1a и Vrn-B1c) present among modern commercial bread wheat cultivars in Russia (including Western Siberia) and a number of temperate countries. For these reasons this work is devoted to obtaining and studying lines possessing a combination of these alleles in the genetic background of the winter cultivar Bez1 and studying their influence on the duration of individual developmental phases and productivity.
Materials and methods
Plant material
In the current paper, the lines of the winter cultivar Bez1 with a combination of dominant alleles were obtained by crossing a i:Bez1 Vrn-B1a and i:Bez1 Vrn-B1c with the i:Bez1 Vrn-A1L (Table 1). PCR markers were used to identify homozygous plants among F2 and F3 hybrids (Table 2). The lines presented in Table 1 were also used as research material.
DNA extraction and PCR amplification
Genomic DNA was extracted from leaves following Sharp et al. (1988). PCR was performed in a 25 μL volume with 12.5 μl BioMaster HS-Taq PCR (2 ×) (Biolabmix, Russia) (100 mM Tris–HCl (pH 8.5 at 25 °C) 100 mM KCl, 0.4 mM of each dNTP, 4 mM MgCl2, 0.06 U/μl Taq DNA polymerase, 0.2% Tween 20, stabilizers of HS-Taq DNA polymerase), 0.5 μM of each primer, 100 ng of genomic DNA and H2O, up to 25 μL. The structure of the used primers and PCR conditions were consistent with the published protocols (Table 2). The reactions were run on a BIO-RAD T100 Thermal Cycler (Bio-Rad, USA). The amplification products were separated by electrophoresis on a 1.5% agarose gel in 1 × TAE buffer stained with ethidium bromide and visualized using the Doc-Print II gel documentation system (Vilber Lourmat, France).
Growth conditions and data analysis
The study of the duration of the individual developmental phases in the lines presented in Table 1 was carried out during spring sowing in 2022 in the experimental field of the Institute of Cytology and Genetics (IC&G SB RAS) (Novosibirsk) (55°N, 82°E) under natural long day (LD) (day length for the May–August period, 17 h). Seed sowing was on May 17 in 2021 and May 31 in 2022. The experiment was also conducted in a hydroponic greenhouse of the Laboratory of Artificial Plant Growth of IC&G under conditions with controlled temperature (20–25 °C) and illumination. The experiment was conducted under both LD (18 h light) and short day (SD, 14 h light) conditions. Plants were grown in pots with volume of 5 L with ten plants per pot. F1 and F3 hybrids were grown in the greenhouse under SD, while F2 hybrids were grown under LD conditions. Phenological stages were recorded following the Zadoks scale (Tottman et al. 1979). The following developmental phases were noted both in the field and in the greenhouse: Z10 (emergence), Z31 (first node), Z39 (flag leaf) and Z60 (full heading). At least 30 plants from each line were studied in each experiment. To assess the significance of the differences between the mean values, Student’s test was used. The productivity of the plants from the new lines was evaluated during cultivation in the field in 2022. Bez1 Vrn-A1a Vrn-B1a and Bez1 Vrn-A1a Vrn-B1c lines were grown in 2021. The productivity components of the main spike and plant, the weight of 1000 grains, and plant height were examined. 25 plants from each line were studied.
Overall, 2021 was quite favorable in the field of plant growth and development (Fig. 1). May was characterized by high temperatures and a strong lack of precipitation. The average daily temperature was 14.2 °C, which was 3.3 °C higher than annual average. May and July were characterized by below-normal precipitation (70% and 40% of annual average, respectively). Precipitation in June was 130% of the annual average. The meteorological conditions of the 2022 growing season, were unfavorable for the growth of plants. May also was characterized by high temperatures and a strong lack of precipitation. The average daily temperature was 15.4 °C, which was 4.5 °C higher than annual average. The total amount of precipitation for the month (3 mm) was only 8% of the annual average. Before sowing, there was a significant lack of moisture in the soil, which, together with the high temperatures, had a negative impact on the initial periods of plant growth. In June, on the contrary, the first ten days turned out to be cold with an average temperature of about 12 °C. Also, the second ten days of July turned out to be dry, during which only 1 mm of precipitation fell, which could have affected the passage of the phases starting from the appearance of the first node until heading. In August, the first and third ten day periods were characterized by below-normal precipitation (40% of annual average). In general, 2022 was characterized by a significant lack of precipitation during the growing season.

Temperature and precipitation in the 2021 and 2022. Temperature is represented as a line, precipitation as a histogram
Results
Production of Bez1 winter wheat lines with a combination of dominant alleles Vrn-A1L Vrn-B1a and Vrn-A1L Vrn-B1c using molecular markers
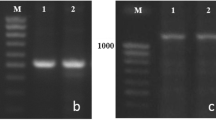
Using PCR markers (Table 1) in plants F1: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a and i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c, amplification fragments possessing the expected size characteristic of heterozygous genotypes were identified (Fig. 2). In the F1 hybrids late maturation prevailed. The duration of the period before the heading of plants in both hybrid combinations was 69 days, which corresponded to the heading time of plants i:Bez1 Vrn-A1L.

Identification of dominant and recessive alleles of Vrn-A1 and Vrn-B1 loci in F1 hybrids: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a (1) and i:Bez1 Vrn-A1L × i:Bez1Vrn-B1c (2) using PCR markers: a Vrn-A1L, b vrn-A1, c Vrn-B1a and Vrn-B1c, d vrn-B1. M – 100 bp Ladder
Analysis was carried out on 38 plants possessing the combination F2 i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a, and 37 of those with the combination F2 i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c. In Fig. 3 and Fig. 4 electrophoregrams of different plant genotypes that were detected in the analysis of F2 hybrids are shown. In the first combination, 27 plants (71.0%) with two dominant alleles were isolated and in the second combination there were 20 plants (54.0%), among which the number of plants with two Vrn genes homozygous for one of the alleles was 32–50%. Homozygous plants with two dominant alleles in the F2 generation2 were not isolated (Tables 3 and 4).

Identification of dominant and recessive alleles of Vrn-A1 and Vrn-B1 loci in F2 hybrids: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a using PCR markers: a Vrn-A1L, b vrn-A1, c Vrn-B1a, d vrn-B1. M – 100 bp Ladder. 1 to 7—different genotypes: 1—Vrn-A1L vrn-A1 vrn-B1 vrn-B1; 2—Vrn-A1L vrn-A1 Vrn-B1a vrn-B1; 3—Vrn-A1L Vrn-A1L Vrn-B1a vrn-B1; 4—Vrn-A1L vrn-A1 Vrn-B1a Vrn-B1a; 5—vrn-A1 vrn-A1 Vrn-B1a Vrn-B1a; 6—Vrn-A1L Vrn-A1L vrn-B1 vrn-B1; 7—vrn-A1 vrn-A1 vrn-B1 vrn-B1

Identification of dominant and recessive alleles of Vrn-A1 and Vrn-B1 loci in F2 hybrids: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c using PCR markers: a Vrn-A1L, b vrn-A1, c Vrn-B1c, d vrn-B1. M—100 bp Ladder. 1 to 6—different genotypes: 1—Vrn-A1L vrn-A1 Vrn-B1c vrn-B1; 2—Vrn-A1L Vrn-A1L Vrn-B1c vrn-B1; 3—Vrn-A1L vrn-A1 vrn-B1 vrn-B1; 4—Vrn-A1L vrn-A1 Vrn-B1c Vrn-B1c; 5—vrn-A1 vrn-A1 Vrn-B1c Vrn-B1c; 6—vrn-A1 vrn-A1 vrn-B1 vrn-B1
The heading time of plants with the combination of F2: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a, carrying the Vrn-B1a allele varied from 48.3 to 53.5 days, and those with the combination F2: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c, carrying the Vrn-B1c allele ranged from 44.8 to 53.9 days. The lowest number of days before heading in both combinations was typical for plants of the Vrn-B1 Vrn-B1 Vrn-A1L vrn-A1 genotype. The highest number of days before heading was observed in plants of the Vrn-A1L vrn-A1 vrn-B1 vrn-B1 genotype (69.0 and 65.4 days, respectively).
Further, during the self-pollination of plants with the Vrn-A1L Vrn-A1L Vrn-B1a vrn-B1 and Vrn-A1L Vrn-A1L Vrn-B1c vrn-B1 genotypes of generation F3, homozygous plants were isolated. In the first combination there were 4 such plants out of the 38 that were analyzed and in the second 13 out of 39 (Table 5 and 6, Fig. 5).

Identification of dominant and recessive alleles of Vrn-A1 and Vrn-B1 loci in F3 hybrids: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a (1–3) and i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c (4–6) using PCR markers: a Vrn-A1L, b Vrn-B1a or Vrn-B1c, c vrn-B1. M—100 bp Ladder
There was a clear trend in heading time differences depending on the allelic state Vrn-B1a and Vrn-B1c locus in F3 hybrids since Vrn-A1L was in the homozygous state in all plants. The effect of Vrn-B1a and Vrn-B1c on heading time was 10–11 days in the presence of the Vrn-A1L allele in homozygous state (Tables 5 and 6). The difference between homozygotes for dominant and recessive alleles was 24 and 38 days in the hybrid combination F3: i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1a and i:Bez1 Vrn-A1L × i:Bez1 Vrn-B1c, respectively. Plants with Vrn-B1c headed earlier than those with Vrn-B1a, which corresponds to the previously obtained data concerning the differences in the timing of heading of isogenic lines i:Bez1 Vrn-B1c and i:Bez1 Vrn-B1a (Emtseva et al. 2013; Chumanova et al. 2020).
Thus, the use of molecular markers has proven to be effective in isolating plants, homozygous for Vrn loci targetting in F3 generation hybrids.
Determination of the influence of combinations of dominant alleles of VRN-1 loci on the heading time and the duration of individual developmental phases
The earliest maturing lines in the field and in the greenhouse were Bez1 Vrn-A1L Vrn-B1a and Bez1 Vrn-A1L Vrn-B1c (Table 7, Figs. 6 and 7) which practically did not differ from each other with regard to the duration up to Z60 period (38.7 and 38.1 days in the field and 40.9 and 42.2 in the greenhouse). In addition, these lines differed significantly in heading time from NIL with the allele Vrn-A1a associated with faster maturation, which possessed a duration of the up to Z60 period which varied from 44.5 to 49.2 days. On the contrary the Vrn-A1L allele is associated with late maturation. NIL was headed in 62.8–69.7 days. The heading time of i:Bez1 Vrn-B1c varied from 47.8 to 59.4 days, and i:Bez1 Vrn-B1a from 48.6 to 59.3 days. Plants of lines Bez1 Vrn-A1L Vrn-B1a and Bez1 Vrn-A1L Vrn-B1c had faster maturation rates than the original NILs with the alleles Vrn-A1L, Vrn-B1a and Vrn-B1c, respectively (P < 0.05), although, initially no differences were assumed with respect to carriers of the Vrn-B1. Thus, the difference in heading time between Bez1 Vrn-A1L Vrn-B1a and NILs with Vrn-A1L and Vrn-B1a alleles was 11.6–25.3 and 3.7–8.7 days depending on the vegetation, respectively (P < 0.05), and the difference between Bez1 Vrn-A1L Vrn-B1c and NILs with alleles Vrn-A1L and Vrn-B1c were 16.2–28.0 and 6.1–8.2 days, respectively (P < 0.05). At the same time, under LD conditions in the greenhouse, there were no differences in heading time between Bez1 Vrn-A1L Vrn-B1a and Bez1 Vrn-A1L Vrn-B1c, in the field or in the greenhouse under SD conditions, the line with the Vrn-B1c allele had faster heading than with the Vrn-B1a allele and the difference reached a period of 6.6 days (P < 0.05). In addition, it turned out that in terms of heading time Bez1 Vrn-A1L Vrn-B1c practically did not differ from the faster maturation line i:Bez1 Vrn-A1a. The observed differences between the genotypes with regard to the heading time, both in the field and in the greenhouse, are determined by differences in the duration of the up to Z31 period, which mainly determines the differences in the duration of the germination-heading period (Emtseva et al. 2013; Chumanova et al. 2020) (Figs. 6 and 7). For the duration of the periods Z31–Z39 and Z39–Z60, there was no unambiguous trend. It should be noted that for all the studied lines, a reaction to the length of the day associated with the Ppd-D1a allele was characteristic and caused moderate photoperiodic sensitivity of the Bez1 cultivar, which was expressed in an increase in the duration of the period before heading (by 4–11 days) under SD conditions mainly due to an increase in the duration of the up to Z31 period (by 4–14 days) while the duration of the periods Z31–Z39 and Z39–Z60 either showed almost no practical change or only decreased slightly.

The duration of developmental phases according to the Zadoks scale (Tottman et al. 1979) of Bez1 lines with dominant alleles of VRN-1 in field 2022. Significant differences: 1—from i:Bez1 Vrn-A1a, 2—from i:Bez1 Vrn-A1L, 3—from i:Bez1 Vrn-B1a, 4—from i:Bez1 Vrn-B1c, 5—from Bez1 Vrn-A1L Vrn-B1a, 6—from Bez1 Vrn-A1a Vrn-B1a, 7—from Bez1 Vrn-A1a Vrn-B1c. *P < 0.05. Error bars are standard deviation of the means

The duration of developmental phases of Bez1 lines with dominant alleles of VRN-1 in greenhouse under SD and LD conditions. Significant differences: 1—from i:Bez1 Vrn-A1a, 2—from i:Bez1 Vrn-A1L, 3—from i:Bez1 Vrn-B1a, 4—from i:Bez1 Vrn-B1c, 5—from Bez1 Vrn-A1L Vrn-B1a, 6—from Bez1 Vrn-A1a Vrn-B1a, 7—from Bez1 Vrn-A1a Vrn-B1c. * P < 0.05. Error bars are SD of the means. SD1 and SD2-experiments under short days in 2022 and in 2023, respectively
Thus, despite the fact that the Vrn-A1L allele causes late maturity in plants, it can be successfully used in breeding for faster maturation as a result of combination with the dominant alleles of the Vrn-B1.
Study of the productivity of lines in the Bez1 cultivar
Determining the influence of the various dominant alleles of Vrn loci and their combinations on productivity indicators is an important task that allows us to select valuable genotypes for breeding purposes. A comparative study of the productivity of new lines revealed that the main spike and plant productivity of the line with a combination of alleles Vrn-A1L and Vrn-B1a, in all of the studied characters, exceeded those with a line possessing a combination of alleles Vrn-A1L and Vrn-B1c (Table 8). The greatest differences were observed in terms of the spikelet number, grain weight per spike and plant productivity (exceeding by 13–25%) (P < 0.05). Comparison of lines with a combination of Vrn-A1a Vrn-B1a and Vrn-A1a Vrn-B1c alleles showed that the first line was more productive than the second line, which is especially evident with respect to grain weight per spike and per plant and weight of 1000 grains (exceeding by 33–49%) (P < 0.05) (Table 8).
Discussion
DNA markers (marker-assisted selection) are widely used in genetic research and plant breeding to solve theoretical problems and accelerate and simplify the breeding process. Markers are also used for the pyramiding of genes controlling the same trait. The use of markers allows for the identification of genotypes containing the targeted combinations of genes at the earliest stages of the creation of breeding material (Song et al. 2023). In this work, homozygous Bez1 cultivar lines were obtained among F3 hybrids. The use of PCR markers developed for the regions of the promoter and the first intron of VRN-1 makes it possible to increase the screening efficiency of alleles of Vrn genes and reduce the selection time of target gene combinations.
The use of different alleles or different combinations of allelic variants of VRN-1 with different phenotypic manifestations at the heading time provides an opportunity for the manipulation of the length of the growing season inherent in bread wheat (Kamran et al. 2014; Shi et al. 2019). The genetic models we used include combinations of Vrn-A1a/Vrn-A1L alleles with Vrn-B1aVrn-B1c. The presence of different sets of genotypes makes it possible to study the genetic effects of different combinations of alleles within a specific targeted genetic background. The lines with the Vrn-A1 locus: Vrn-A1a and Vrn-A1L differ from each other in heading time (Chumanova et al. 2023), and the differences between NILs with Vrn-B1c and Vrn-B1a are were about 4 days (Emtseva et al. 2013). Based on this, the dominant alleles of VRN-1 in order to decrease the duration of the period before heading can be arranged as follows: Vrn-A1a ˃ Vrn-B1c ˃ Vrn-B1a ˃ Vrn-A1L. The data presented and obtained earlier (Chumanova et al. 2020) show that the lines with theVrn-A1a Vrn-B1a and Vrn-A1a Vrn-B1c alleles do not differ in the timing of heading or the duration of the developmental phases, and they are also faster in maturation compared to i:Bez1 Vrn-A1a. It is known that Vrn-A1 has the strongest effect on the acceleration of heading among the three VRN-1 genes (Goncharov et al. 2004) and that the Vrn-A1a allele is epistatic with respect to the alleles of the Vrn-B1 (Li et al. 2017). However, plants bearing combinations of two or three dominant alleles tend to possess faster maturation than those with a single allele, including Vrn-A1a, which is associated with the additive effect of genes (Potokina et al. 2012; Kiss et al. 2014; Zhang et al. 2014; Shcherban et al. 2015; Efremova et al. 2016). Interesting results were obtained when studying the second group of lines possessing a combination of alleles. The introduction of the dominant Vrn-A1L allele into the genotype of the winter cultivar Bez1, which determines the late maturity of the plants, in combination with alleles of the Vrn-B1 led to the production of faster maturing lines relative to the original NILs, carriers of these alleles. It is probable that there is also an additive effect of homeologous Vrn genes, that is that in the presence of Vrn-B1, there is an increase in the expression of Vrn-A1L, which manifests itself in a decrease in the duration of the period from germination to heading, due to a decrease in the duration of the period before the appearance of the first node. We hope that experiments into the study of expression, which we plan to carry out in the future, will help to answer this question. In addition, the Bez1 Vrn-A1L Vrn-B1c line differed slightly from the NIL with the Vrn-A1a. Similar results were obtained in the work (Dowla et al. 2020). A synthetic cultivar of wheat with Vrn-A1L from Langdon durum wheat and Vrn-B1a, had a pre-heading period duration only 2 days longer than lines with Vrn-A1a Vrn-B1a alleles. In our work, the combination of Vrn-A1L and Vrn-B1c alleles leads to slightly earlier heading, as well as lessening the duration of the period before the appearance of the first node, than that of the combination of Vrn-A1L and Vrn-B1a, which may be explained by differences in the heading times of monogenic carriers with Vrn-B1c and Vrn-B1a as described Emtseva et al. (2013) as well as the genetic background of the lines.
It is well known that the heading time of bread wheat and the duration of the individual developmental phases are extremely important features that determine the ability of bread wheat cultivars to adapt to varied climatic conditions, thereby contributing to a consistently high productivity (Kamran et al. 2014). When choosing cultivars for cultivation that are most suitable for the climatic conditions of a particular region, it is necessary to take into account the maturation speed of different cultivars, so that the most sensitive phases of development, take place under the most optimal environmental conditions (Santra et al. 2009; Gomez et al. 2014; Kamran et al. 2014; Grogan et al. 2016; Amo et al. 2022). In Siberia the selection of early ripening cultivars is one of the important aspects in the breeding of spring bread wheat. Among modern widespread cultivars in Western Siberia, genotypes with a combination of Vrn-A1a and Vrn-B1a or Vrn-B1c alleles predominate, since it is this combination of alleles that ensures optimal heading times and yield potential when taking into account the specific features of the climatic conditions of the region (Likhenko et al. 2015; Efremova et al. 2016; Smolenskaya et al. 2022). The lines with a combination of Vrn-A1L and Vrn-B1a or Vrn-B1c alleles can be classified as medium-ripe in terms of heading time, which demonstrates the importance of the results obtained for the conditions of Western Siberia. Therefore, the Vrn-A1L allele, which was previously introgressed by us from T. petropavlovskyi, which had previously not yet been involved in the selection of bread wheat, can be used in breeding for faster maturation as a result of the combination with alleles of the Vrn-B1. Introgression of dominant alleles of Vrn genes from related cereal species, or the use of rare alleles already available in the gene pool but not yet seeing wide distribution, is a popular direction for of fundamental and breeding research, since it allows for the increasing of the adaptive potential of bread wheat by increasing the existing allelic diversity in genes that determine the timing of key developmental phases, including flowering.
A comparative study of the productivity of new lines showed that a line combining Vrn-A1L and Vrn-B1a turned out to result in higher productivity than that containing Vrn-A1L and Vrn-B1c. However, a comparison of combinations of Vrn-A1a Vrn-B1a and Vrn-A1a Vrn-B1c alleles with each other, in fact showed the opposite to be true. It is known that a longer vegetative period allows the wheat plant to produce a greater number of productive shoots and consequently a high number of grains under favorable conditions (Royo et al. 2018). Probably, the observed differences in productivity between Bez1 Vrn-A1L Vrn-B1a and Bez1 Vrn-A1L Vrn-B1c lines are due to the longer period before first node formation of the first line (3 days difference), due to which we observe the formation of a greater number of productive shoots and, consequently, a higher grain number and grain weight per plant. However, the observed differences between Bez1 Vrn-A1L Vrn-B1a and Bez1 Vrn-A1L Vrn-B1c lines cannot be explained by differences in the duration of developmental phases. It is known that in Western Siberia cultivars with a combination of alleles Vrn-A1a Vrn-B1c are more common than with a combination of Vrn-A1a Vrn-B1a (Efremova et al. 2016; Smolenskaya et al. 2022), which is associated not only with a shorter period to earing, but also, probably, with greater adaptation to climatic conditions and productivity of such genotypes. Unfortunately, experiments conducted over the course of different years do not allow a comparison to be made between these four combinations, since the realization of productive potential occurs in close interaction with the genotype and the external conditions. Therefore, we plan to conduct repeated experiments in order to obtain unambiguous conclusions regarding the effect of different combinations of VRN-1 on actual yield, which would allow us to identify genotypes capable of maximizing their yield potential in Western Siberia.
Thus, the possibility has been demonstrated for the manipulation of the length of the growing season of bread wheat by combining the Vrn-A1L allele, which determines late maturity, with alleles of the Vrn-B1 locus, has been demonstrated to facilitate obtaining earlier ripening genotypes that can be used for breeding of bread wheat in Western Siberia.
Conclusion
In this work, lines with two dominant alleles: Vrn-A1L Vrn-B1a and Vrn-A1L Vrn-B1c were obtained within the genetic background of the winter cultivar of wheat Bez1. Isolation of homozygous plants was performed using marker-assisted selection. The obtained lines produced a shorter duration of germination-heading and germination-first node periods relative to the original NILs with alleles Vrn-A1L, Vrn-B1a and Vrn-B1c. At the same time, the Bez1 Vrn-A1L Vrn-B1c line possessed a shorter duration of the period before heading and the period of germination of the first node compared to Bez1 Vrn-A1L Vrn-B1a practically did not differ from the precocious NIL with the Vrn-A1a allele. In summary, Vrn-A1L allele can be successfully used in breeding for faster maturation in Western Siberia in combination with Vrn-B1a and Vrn-B1c alleles.
References
Amo A, Serikbay D, Song L et al (2022) Vernalization and photoperiod alleles greatly affected phenological and agronomic traits in bread wheat under autumn and spring sowing conditions. Crop Environ 1:241–250. https://doi.org/10.1016/J.CROPE.2022.11.002
Beales J, Turner A, Griffiths S et al (2007) A pseudo-response regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor Appl Genet 115:721–733. https://doi.org/10.1007/s00122-007-0603-4
Chen A, Dubcovsky J (2012) Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLOS Genet 8:e1003134. https://doi.org/10.1371/JOURNAL.PGEN.1003134
Chumanova EV, Efremova TT, Kruchinina YV, Pershina LA (2018) Development and investigation of common wheat lines of winter cultivar Bezostaya 1 with combinations of dominant alleles of VRN-1 loci. Vavilovskii Zhurnal Genet Selektsii 22:951–956. https://doi.org/10.18699/VJ18.437
Chumanova EV, Efremova TT, Kruchinina YV (2020) The effect of different dominant VRN alleles and their combinations on the duration of developmental phases and productivity in common wheat lines. Russ J Genet 56:822–834. https://doi.org/10.1134/S1022795420070029
Chumanova E, Efremova T, Vavilova V (2023) Characterization of the VRN-A1 allele introgressed from T. aestivum ssp petropavlovskyi that influences the heading time in bread wheat. Euphytica 219:53. https://doi.org/10.1007/s10681-023-03178-1
Dowla MANNU, Islam S, Stefanova K et al (2020) Effects on the adaptation of selected wheat (Triticum aestivum L.) advanced lines across diverse water-limited environments of Western Australia. Agric 10:1–18. https://doi.org/10.3390/agriculture10100470
Efremova T, Arbuzova V, Leonova I, Makhmudova K (2011) Multiple allelism in the Vrn-B1 locus of common wheat. Cereal Res Commun 39:12–21. https://doi.org/10.1556/CRC.39.2011.1.2
Efremova TT, Chumanova EV, Trubacheeva NV et al (2016) Prevalence of VRN1 locus alleles among spring common wheat cultivars cultivated in Western Siberia. Russ J Genet 52:146–153. https://doi.org/10.1134/S102279541601004X
Emtseva MV, Efremova TT, Arbuzova VS (2013) The influence of Vrn-B1a and Vrn-B1c alleles on the length of developmental phases of substitution and near-isogenic lines of common wheat. Russ J Genet 49:545–552. https://doi.org/10.1134/S1022795413050050
Fu D, Szűcs P, Yan L et al (2005) Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol Genet Genom 273:54–65. https://doi.org/10.1007/s00438-004-1095-4
Gomez D, Vanzetti L, Helguera M et al (2014) Effect of Vrn-1, Ppd-1 genes and earliness per se on heading time in Argentinean bread wheat cultivars. F Crop Res 158:73–81. https://doi.org/10.1016/J.FCR.2013.12.023
Goncharov NP (2004) Response to vernalization in wheat: its quantitative or qualitative nature. Cereal Res Commun 32:323–330. https://doi.org/10.1007/BF03543317
Grogan SM, Anderson J, Stephen Baenziger P et al (2016) Phenotypic plasticity of winter wheat heading date and grain yield across the US great plains. Crop Sci 56:2223–2236. https://doi.org/10.2135/CROPSCI2015.06.0357
Ivaničová Z, Jakobson I, Reis D et al (2016) Characterization of new allele influencing flowering time in bread wheat introgressed from Triticum militinae. N Biotechnol 33:718–727. https://doi.org/10.1016/j.nbt.2016.01.008
Kamran A, Iqbal M, Spaner D (2014) Flowering time in wheat (Triticum aestivum L.): a key factor for global adaptability. Euphytica 197:1–26. https://doi.org/10.1007/s10681-014-1075-7
Kippes N, Debernardi JM, Vasquez-Gross HA et al (2015) Identification of the VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proc Natl Acad Sci 112:E5401–E5410. https://doi.org/10.1073/pnas.1514883112
Kiss T, Balla K, Veisz O et al (2014) Allele frequencies in the VRN-A1, VRN-B1 and VRN-D1 vernalization response and PPD-B1 and PPD-D1 photoperiod sensitivity genes, and their effects on heading in a diverse set of wheat cultivars (Triticum aestivum L.). Mol Breed 34:297–310. https://doi.org/10.1007/S11032-014-0034-2
Li G, Boontung R, Powers C et al (2017) Genetic basis of the very short life cycle of “Apogee” wheat. BMC Genomics 18:838. https://doi.org/10.1186/s12864-017-4239-8
Likhenko IE, Stasyuk AI, Shcherban AB et al (2015) Study of allelic composition of Vrn-1 and Ppd-1 genes in early–ripening and middle–early varieties of spring soft wheat in Siberia. Russ J Genet Appl Res 5:198–207. https://doi.org/10.1134/S2079059715030107
Loukoianov A, Yan L, Blechl A et al (2005) Regulation of VRN-1 vernalization genes in normal and transgenic polyploid wheat. Plant Physiol 138:2364–2373. https://doi.org/10.1104/pp.105.064287.2364
Milec Z, Tomková L, Sumíková T, Pánková K (2012) A new multiplex PCR test for the determination of Vrn-B1 alleles in bread wheat (Triticum aestivum L.). Mol Breed 30:317–323. https://doi.org/10.1007/s11032-011-9621-7
Milec Z, Sumíková T, Tomková L, Pánková K (2013) Distribution of different Vrn-B1 alleles in hexaploid spring wheat germplasm. Euphytica 192:371–378. https://doi.org/10.1007/s10681-013-0863-9
Muterko A, Balashova I, Cockram J et al (2015) The new wheat vernalization response allele Vrn-D1s is caused by dna transposon insertion in the first intron. Plant Mol Biol Report 33:294–303. https://doi.org/10.1007/s11105-014-0750-0
Nishida H, Yoshida T, Kawakami K et al (2013) Structural variation in the 5′ upstream region of photoperiod-insensitive alleles Ppd-A1a and Ppd-B1a identified in hexaploid wheat (Triticum aestivum L.), and their effect on heading time. Mol Breed 31:27–37. https://doi.org/10.1007/s11032-012-9765-0
Potokina EK, Koshkin VA, Alekseeva EA et al (2012) The combination of the Ppd and Vrn gene alleles determines the heading date in common wheat varieties. Russ J Genet Appl Res 2:311–318. https://doi.org/10.1134/S2079059712040089
Royo C, Ammar K, Alfaro C, Dreisigacker S, García del Moral LF, Villegas D (2018) Effect of Ppd-1 photoperiod sensitivity genes on dry matter production and allocation in durum wheat. Field Crops Res 221:358–367. https://doi.org/10.1016/j.fcr.2017.06.005
Santra DK, Santra M, Allan RE et al (2009) Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1, and Vrn-D1 in spring wheat germplasm from the Pacific Northwest Region of the U.S.A. Plant Breed 128:576–584. https://doi.org/10.1111/j.1439-0523.2009.01681.x
Sharp PJ, Kreis M, Shewry PR, Gale MD (1988) Location of β-amylase sequences in wheat and its relatives. Theor Appl Genet 75:286–290. https://doi.org/10.1007/BF00303966
Shcherban AB, Salina EA (2017) Evolution of VRN-1 homoeologous loci in allopolyploids of Triticum and their diploid precursors. BMC Plant Biol 17:188. https://doi.org/10.1186/s12870-017-1129-9
Shcherban A, Emtseva M, Efremova T (2012a) Molecular genetical characterization of vernalization genes Vrn-A1, Vrn-B1 and Vrn-D1 in spring wheat germplasm from Russia and adjacent regions. Cereal Res Commun 40:351–361. https://doi.org/10.1556/CRC.40.2012.3.4
Shcherban AB, Efremova TT, Salina EA (2012b) Identification of a new Vrn-B1 allele using two near-isogenic wheat lines with difference in heading time. Mol Breed 29:675–685. https://doi.org/10.1007/s11032-011-9581-y
Shcherban AB, Börner A, Salina EA (2015) Effect of VRN-1 and PPD-D1 genes on heading time in European bread wheat cultivars. Plant Breed 134:49–55. https://doi.org/10.1111/pbr.12223
Shi C, Zhao L, Zhang X et al (2019) Gene regulatory network and abundant genetic variation play critical roles in heading stage of polyploidy wheat. BMC Plant Biol 19:6. https://doi.org/10.1186/s12870-018-1591-z
Smolenskaya S, Efimov V, Kruchinina Y et al (2022) Earliness and morphotypes of common wheat cultivars of Western and Eastern Siberia. Vavilovskii Zhurnal Genet i Sel = Vavilov J Genet Breed 26:662–674. https://doi.org/10.18699/VJGB-22-81
Song L, Wang R, Yang X et al (2023) Molecular markers and their applications in marker-assisted selection (MAS) in bread wheat (Triticum aestivum L.). Agric 13:642. https://doi.org/10.3390/AGRICULTURE13030642
Stelmakh AF, Avsenin VI (1996) Alien introgression of spring habit dominant genes into bread wheat. Euphytica 89:65–68. https://doi.org/10.1007/BF00015720
Tottman DR, Makepeace RJ, Broad H (1979) An explanation of the decimal code for the growth stages of cereals, with illustrations. Ann Appl Biol 93:221–234. https://doi.org/10.1111/J.1744-7348.1979.TB06534.X
Whittal A, Kaviani M, Graf R et al (2018) Allelic variation of vernalization and photoperiod response genes in a diverse set of North American high latitude winter wheat genotypes. PLoS ONE 13:e0203068. https://doi.org/10.1371/JOURNAL.PONE.0203068
Yan L, Loukoianov A, Tranquilli G et al (2003) Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci U S A 100:6263–6268. https://doi.org/10.1073/pnas.0937399100
Yan L, Helguera M, Kato K et al (2004a) Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor Appl Genet 109:1677–1686. https://doi.org/10.1007/s00122-004-1796-4
Yan L, Loukoianov A, Blechl A et al (2004b) The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science 303:1640–1644. https://doi.org/10.1126/science.1094305
Yan L, Fu D, Li C et al (2006) The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc Natl Acad Sci USA 103:19581–19586. https://doi.org/10.1073/pnas.0607142103
Zhang XK, Xiao YG, Zhang Y et al (2008) Allelic variation at the vernalization genes Vrn-A1, Vrn-B1, Vrn-D1, and Vrn-B3 in Chinese wheat cultivars and their association with growth habit. Crop Sci 48:458–470. https://doi.org/10.2135/CROPSCI2007.06.0355
Zhang J, Wang Y, Wu S et al (2012) A single nucleotide polymorphism at the Vrn-D1 promoter region in common wheat is associated with vernalization response. Theor Appl Genet 125:1697–1704. https://doi.org/10.1007/s00122-012-1946-z
Zhang J, Dell B, Biddulph B et al (2014) Vernalization gene combination to maximize grain yield in bread wheat (Triticum aestivum L.) in diverse environments. Euphytica 198:439–454. https://doi.org/10.1007/S10681-014-1120-6
Zhang X, Gao M, Wang S et al (2015) Allelic variation at the vernalization and photoperiod sensitivity loci in Chinese winter wheat cultivars (Triticum aestivum L.). Front Plant Sci 6:470. https://doi.org/10.3389/fpls.2015.00470
Zhang B, Wang X, Wang X et al (2018) Molecular characterization of a novel vernalization allele Vrn-B1d and its effect on heading time in Chinese wheat (Triticum aestivum L.) landrace Hongchunmai. Mol Breed 38:137. https://doi.org/10.1007/s11032-018-0870-6
Zhang B, Guo Y, Fan Q et al (2022) Characterization and distribution of novel alleles of the vernalization gene Vrn-A1 in Chinese wheat (Triticum aestivum L.) cultivars. Crop J 11:852–962. https://doi.org/10.1016/j.cj.2022.10.002
Acknowledgements
Reproduction of the plant material was carried out on the basis of the Laboratory of Artificial Plant Growth of ICG SB RAS within the budgetary project FWNR-2022-0017.
Funding
This study was supported by the Russian Science Foundation (Grant No. 22–26-00085).
Author information
Authors and Affiliations
Contributions
EC wrote and edited the article, performed PCR, field and greenhouse experiments, and data analysis. TE conceived and designed research, edited the article, performed field and greenhouse experiments.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Ankica Kondic-Spika.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chumanova, E., Efremova, T. Marker-assisted development of wheat lines of the winter cultivar Bezostaya 1 and the effects of interaction between alleles of Vrn-A1L and Vrn-B1 loci on heading time. CEREAL RESEARCH COMMUNICATIONS (2023). https://doi.org/10.1007/s42976-023-00478-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42976-023-00478-z