
Abstract
Soil drought, which is one of the most threatening abiotic plant stressors, negatively impacts plant growth and yields. Plant phytohormones play a key role in the formation of plant responses to stress. We investigated the effect of moderate soil drought, created in controlled laboratory conditions on plant growth and peculiarities of endogenous abscisic (ABA), indole-3-acetic (IAA), gibberellic (GA3) and salicylic (SA) acids and cytokinins (CKs) accumulation in organs of two closely related wheat species: Triticum aestivum and T. spelta. We demonstrated that following the cessation of watering (within four days), shoots and roots of 18-day-old plants of both species accumulated ABA and SA, while the level of GA3 and IAA decreased. Pool of CKs significantly increased in roots and decreased in shoots. On day 21 following recovery, an increase in ABA content occurred in wheat shoots and roots and spelt shoots. IAA level increased in both species, but not to the levels measured in control plants. The highest level of GA3 was present in wheat roots. While the content of SA in wheat was at control level, spelt SA levels significantly exceeded the controls. There were complex changes in CKs pool, the nature of which depended on the species and plant organ. We propose that specific alterations in the nature of accumulation, localization, and balance between certain classes of phytohormones in wheat and spelt organs under the action of moderate soil drought are one of the main factors in systemic response to stress and the formation of an adaptation strategy.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Through anthropogenic stresses on soil and water supply and due to global climate changes, soil droughts have become a major abiotic threat, which throttles metabolic processes, depresses plant growth and development, and reduces crop yields (Iqbal et al. 2021). Droughts induce closing of stomata, reduce the supply of carbon dioxide to chloroplasts, disrupt photosynthesis (Hu et al. 2019), and retard the growth of shoots (Lind et al. 2015). Concurrently, in search for new sources of water, additional roots are grown, which increases the ratio of root to shoot (Dietrich 2018).
Exogenous stresses precipitate a cascade of significant morphological, physiological, biochemical and molecular changes in affected plants. These changes are regulated by the hormonal system and enable plant survival under unfavorable conditions (Gupta et al. 2020). ABA acts as the main trigger for the signaling cascade that is activated in response to drought stress. Receptors and key transduction factors for ABA-signaling have been identified in major grain crops, including wheat, rice, corn, and barley (Daszkowska-Golec and Szarejkom 2013). Initiation of ABA biosynthesis and hyperexpression of the corresponding hormonal signaling pathway have been shown to increase drought tolerance (Estrada-Melo et al. 2015). The former study identified reductions in stomatal apertures and stress-induced transcriptomic changes, while the latter study reported only transcriptomic changes.
Drought was associated with an increase in ABA content and a reduction of IAA accumulation in winter wheat (Sakhabutdinova et al. 2003). Multiple studies showed that hormones stimulating plant growth (auxins, cytokinins and gibberellins) reduce tolerance to water deficits (Ullah et al. 2018). In drought conditions, researchers observed reduced yields and lower protein and carbohydrate wheat grain content, which occurred in the context of lower levels of endogenous IAA, t-ZR and GA1+3 and accumulation of ABA (Xie et al. 2003). When subjected to drought, wheat roots accumulated auxins, which regulated daytime and nighttime water consumption, modulated hydraulic properties, maintained water supply and increased yields (Sadok and Schoppach 2019). Exogenous SA mitigated the negative impact of drought on wheat plants through a reduction of photosynthetic volume, maintenance of membrane permeability, induction of stress protein biosynthesis, and enhanced activity of antioxidant enzymes (Khalvandi et al. 2021).
Winter wheat Triticum aestivum L. has a leading position among the most important food crops of Ukraine. New genotypes of Ukrainian selection are characterized by ecological plasticity, heat, drought, and frost resistance (Morgun et al. 2016). In modern wheat production, the trend of revival, selection and introduction of so-called antique cereals, among which spelt wheat Triticum spelta L., is clearly remarked. Spelt wheat crosses well with tetraploid wheat, it is used in breeding programs for the improvement of hard and soft wheat, it is resistant to climatic factors and suitable for low-cost organic farming (Lacko-Bartošová et al. 2010; Escarnot et al. 2012; Babenko et al. 2018).
In our previous research, we found that short-term heat stress (+ 40 °C, 2 h.) induced rapid specific changes in the accumulation and distribution of endogenous gibberellic and salicylic acids and cytokinins in the winter wheat cv ‘Podolyanka’ and spelt wheat cv ‘Frankenkorn’ organs (Kosakivska et al. 2021). The aim of this work was to analyze the character of accumulation and distribution of endogenous ABA, IAA, GA3, CKs and SA in shoots and roots of these two species, and to identify specific features in phytohormonal balance associated with response to moderate soil drought. The study of endogenous phytohormones involved in the formation of adaptation mechanisms is important for obtaining drought-resistant varieties of cereals, especially under expected adverse climatic changes.
Materials and methods
Plant material
We studied 18- and 21-day-old plants of wheat (Triticum aestivum L., cv ‘Podolyanka’) and spelt wheat (Triticum spelta L., cv ‘Frankenkorn’), the seeds of which were obtained from Institute of Plant Physiology and Genetics of the NAS of Ukraine (Kyiv) and National Centre for Plant Genetic Resources of Ukraine in Kharkiv. Plants of winter wheat (Triticum aestivum L., cultivar ‘Podolyanka’) are highly productive, frost- and drought-resistant. Plants of spelt wheat (Triticum spelta L., cultivar ‘Frankenkorn’) are a medium-sized, resistant to lodging, frost-resistant. Seed calibration and sterilization, as well as plant growing conditions, were described previously (Kosakivska et al. 2021).
Abiotic stress treatments and sample collection
To simulate soil drought, watering of 14-day-old plants was stopped for four days until the leaves withered and the moisture content of the substrate was halved. On the 18th day in the stage of 2–3 leaves watering of plants was resumed. For recovery, plants were grown to 21-day-old (stage of 3–4 leaves) in the above-mentioned conditions. For the experiment, shoots and roots of 18- and 21-day-old plants were selected.
Extraction of IAA, ABA, GA3, SA and CKs
For extraction of endogenous IAA and ABA, we used the method (Kosakivska et al. 2022), and sorption and separation of GA3, SA and CKs were carried out accordingly (Kosakivska et al. 2021).
Determination of IAA, ABA, GA3, SA and CKs
We applied the method of high-performance liquid chromatography on Agilent 1200 LC/MS series instrument (USA) with diode-array detector G1315B and single quadrupole mass detector Agilent G6120A for analytical determination of IAA, ABA, GA3, SA and CKs (Kosakivska et al. 2021; 2022).
Statistical analyses
All measurements were taken with three biological and three analytical replicates. Statistical analysis was carried out in Statistica, version 6.0 (StatSoft Inc.). One-way analysis of variance (ANOVA) was conducted, with P ≤ 0.05 considered statistically significant (Van Emden 2008).
Results
Effect of moderate soil drought on morphophenological characteristics of wheat and spelt plants
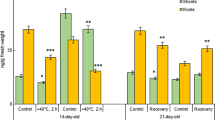
Moderate drought inhibited the growth of wheat and spelt shoots and roots, and the root system of both species was more sensitive (Fig. 1). The height of wheat shoots after drought stress was 6.5%, and after recovery it was 6.6% lower than control. Root length decreased by 28.2% after stress, and after recovery it was 27.1% less than control. The height of spelt shoots decreased by 9.0% after drought stress and by 12.0% after recovery. The length of spelt roots decreased by 28.2% after stopping watering and by 16.2% after recovery. Some changes were observed in fresh weight (FW) and dry weight (DW) of organs. After drought stress, FW of wheat shoots and roots decreased by 25.8% and by 5.6%, respectively. At the same time, there were no significant changes in DW of the organs. On the 21st day (after recovery), compared to control, FW of wheat shoots was 35% lower, and FW of wheat roots was 8.7% lower. Compared to control, DW of shoots was lower by 15.2%, and DW of roots was 10.9% lower. After stopping watering, the FW and DW of 18-day-old spelt shoots decreased by 23.8% and 9.9%, respectively, and in roots by 21.7% and 13.3%, respectively. After recovery on the 21st day, the FW and DW of the shoots were lower than control by 20.4% and 6.2%, respectively, and the roots by 15.0% and 13.0%, respectively.

Effect of moderate soil drought (four days without watering) on morphophenological characteristics of 18-day-old Triticum aestivum L. cv ‘Podolyanka’ and Triticum spelta L. cv ‘Frankenkorn’ plants and of 21-day-old plants in recovery period. Note: except for height of shoots and length of roots, the figure presents average fresh weight and average dry weight of plant organ. *P < 0.05; **P < 0.01 compared to control at these stages of vegetation
Therefore, moderate soil drought inhibited the growth of 18-day-old wheat and spelt plants and negatively affected the morphophenological characteristics of 21-day-old plants after recovery. Roots of both species were more resistant, and spelt root system recovered better compared to wheat root system.
Changes in endogenous phytohormones accumulation and distribution associated with moderate soil drought
Abscisic acid was stimulated after drought stress in wheat roots and significantly in spelt shoots
Moderate soil drought induced an increase in the content of endogenous ABA in 18-day-old winter shoots and roots by 5% and 29.8%, respectively. In post-stressed plants, the site of hormone accumulation was in the root system and was 1.8-fold higher than in shoots. In control plants, ABA is dominant in roots. On the 21st day following recovery, the content of ABA in the shoots of stressed plants was 27% higher, and in the roots 227% higher than in control. The center of hormone accumulation after recovery shifted to the shoots, where its content was 1.7-fold higher than in the roots. In contrast, the level of endogenous ABA in the shoots of 21-day control plants was 2.7-fold higher than in the roots (Fig. 2).

Dynamics and distribution of endogenous abscisic acid in the organs of 18-day-old plants of Triticum aestivum L., cv ‘Podolyanka’ and spelt wheat Triticum spelta L., cv ‘Frankenkorn’ after moderate soil drought (four days without watering), and on the 21st day after recovery (ng/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation
After a moderate soil drought, the content of ABA in the shoots and roots of spelt plants increased by 334% and 25%, respectively. The ABA level in the shoots was 8.8-fold higher than in the roots. In 21-day-old plants, after recovery, the hormone content decreased in the shoots 3.5-fold, in the roots—1.2-fold, but the recorded value in roots was below control and was 80.1 ± 4.0 ng/g DW, while in the shoots it was 249.3 ± 12.5 ng/g DW—1.1-fold higher than control. In the shoots of regenerative plants, the content of ABA was 3.7-fold higher than in the roots (Fig. 2).
The content of endogenous ABA was higher in 18-day-old control wheat plants than in spelt: 328.5 ± 16.4 and 277.0 ± 13.9 ng/g DW, respectively. After moderate soil drought, at a soil moisture content of 30%, the level of endogenous ABA in both species increased, reaching in wheat 656.4 ± 32.8 ng/g DW, and in spelt 959.4 ± 47.7 ng/g DW. After recovery on the 21st day, ABA dominated in wheat and spelt shoots, but in wheat the level of hormone was higher.
Drought stress caused a decrease in IAA content and stimulated hormone accumulation after recovery
The content of endogenous IAA in 18-day-old control plants of wheat cv ‘Podolyanka’ was 668.3 ± 33.4 (shoots) and 662.5 ± 23.1 (roots) ng/g DW, which exceeded spelt cv ‘Frankenkorn’ by 73%/ and 92%, respectively. After a moderate soil drought, the level of endogenous IAA decreased in the shoots and roots of wheat cv ‘Podolyanka’ 1.8—and twofold, respectively, and in spelt cv ‘Frankenkorn’—1.6- and 1.01-fold. The site of hormone accumulation was located in the shoots of wheat and spelt control and stressed plants (Fig. 3).

Dynamics and distribution of indole-3-acetic acid in the organs of 18-day-old plants of Triticum aestivum L., cv ‘Podolyanka’ and spelt wheat Triticum spelta L., cv ‘Frankenkorn’ after moderate soil drought (four days without watering), and on the 21st day after recovery (ng/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation
After recovery period, the content of IAA in 21-day-old wheat and spelt plants did not reach control levels (Fig. 3), 15.5% and 9.8% lower, respectively. The total content of IAA level in wheat plants after recovery reached 1118.3 ± 55.9 ng/g DW, which was 3.7-fold higher than in spelt, where it was 306.2 ± 15.3 ng/g DW. In general, during the recovery period in wheat roots, the content of IAA was 1.3-fold higher than in the shoots, while in spelt—1.3-fold lower. The site of IAA accumulation in 2-day-old wheat plants was concentrated in the roots, while in spelt—it was concentrated in the shoots.
Drought stress resulted in diminution of gibberellic acid level more pronounce in spelt roots and induced the accumulation of hormone in both species after recovery
Moderate soil drought caused a decrease in the content of endogenous GA3 in 18-day-old wheat cv ‘Podolyanka’ organs. In shoots and roots, the hormone level decreased by 48.2% and 46.2%, respectively. GA3 dominated in the roots of stressed and control plants. The hormone content was 1.3-fold higher in the roots of stressed plants than in their shoots. On the 21st day following recovery, the content of GA3 in the shoots and roots increased compared to stressed plants, but the recorded values were lower than the control, 1.4- and 1.3-fold, respectively. Accumulation of GA3 after recovery was more active in the roots (lower than control by 22.5%) than in the shoots (lower than control by 29.2%) (Fig. 4).

Dynamics and distribution of endogenous gibberellic acid in the organs of 18-day-old plants of Triticum aestivum L., cv ‘Podolyanka’ and spelt wheat Triticum spelta L., cv ‘Frankenkorn’ after moderate soil drought (four days without watering), and on the 21st day after recovery (ng/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation
Similar changes in endogenous GA3 content were observed in spelt cv ‘Frankenkorn’ organs after moderate soil drought. Roots were more sensitive to stress: the hormone level decreased by 68% and by 26% in shoots (Fig. 3). In contrast to wheat, in stressed spelt plants, GA3 dominated in the shoots. The content of GA3 in the shoots and roots of stressed spelt plants after recovery was 6.7% and 30.5% lower, respectively, than in the organs of control 21-day-old plants. Gibberellins dominated in the roots of all (except 18-day-old stressed plants) studied variants.
Salicylic acid was highly stimulated in spelt by direct drought stress
After moderate soil drought, the level of endogenous SA increased in shoots and roots of 18-day-old wheat plants by 24% and 1.7%, respectively. In spelt, the content of SA increased by 92% in shoots and by 189% in roots. After recovery on the 21st day, the concentration of SA in wheat plants exceeded the control. Compared with stressed 18-day-old plants, the hormone level increased in shoots by 37.2% and in roots by 51.6% (Fig. 5).

Dynamics and distribution of endogenous salicylic acid in the organs of 18-day-old plants of Triticum aestivum L., cv ‘Podolyanka’ and spelt wheat Triticum spelta L., cv ‘Frankenkorn’ after moderate soil drought (four days without watering), and on the 21st day after recovery (µg/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation
On the 21st day after recovery, the content of endogenous SA in spelt shoots was much lower than control, while in the roots SA levels exceeded control. Compared with 18-day-old stressed plants, SA level decreased 2.7-fold in shoots and 2.5-fold in roots. In general, the character of SA distribution between organs of wheat and spelt was the same, with the dominance of this hormone in shoots. Spelt plants were characterized by significantly lower content of endogenous SA in control condition, after drought, and in the recovery period (Fig. 5).
Significant remodeling of the cytokinins pool took place in wheat and spelt organs after drought stress
We identified in winter wheat organs three forms of CKs: trans-zeatin (t-Z), trans-zeatin-O-glucoside (t-ZOG) and isopentenyladenine (iP). Trans-zeatin riboside (t-ZR) and isopentenyladenosine (iPA) were present in trace amounts. After moderate soil drought, the content of t-Z in roots and shoots increased, and level of t-ZOG and iP decreased significantly. Total CKs content in shoots decreased, and in roots increased, amounting to 2.649 ± 0.13 and 2.660 ± 0.13 ng/g DW, respectively. On the 21st day after recovery, the site of t-ZOG moved to the roots, while t-Z and iP dominated in shoots. The content of t-Z and iP decreased and the level of t-ZOG increased in comparison with 18-day-old stressed plants. In stressed wheat plants after recovery, the content of active forms t-Z and iP in the shoots and t-Z in the roots exceeded control 21-day-old plants, by 15.7%, 1.8% and 15.9%, respectively. At the same time, the levels of t-ZOG in shoots and roots and iP in roots remained below control levels (Fig. 6).

Dynamics and distribution of endogenous cytokinins in the organs of 18-day-old plants of Triticum aestivum L. cv ‘Podolyanka’ after moderate soil drought (four days without watering) and on the 21st day after recovery (ng/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation. Notes: t-Z—trans-zeatin, iP—isopentenyladenine, ZOG—zeatin-O-glucoside; traces—less than 0,5 ng/g DW
In spelt, wheat organs were found five forms of CKs. After moderate soil drought, the content of t-Z and t-ZR decreased, at the same time the level of t-ZOG, and especially iPA in shoots decreased. The content of all CKs forms, except t-Z, in the roots increased. Total CKs content in shoots of stressed spelt plants reached 327.1 ± 16.35 ng/g DW, compared to 392.7 ± 19.64 ng/g DW in control plants. In the roots of stressed plants, the CKs pool was measured at 272.7 ± 13.64 ng/g DW, and in control plants: at 179.0 ± 8.95 ng/g DW. After recovery in 21-day-old plants CKs forms, except t-ZR, dominated in roots. The content of t-Z, t-ZR, t-ZOG and iP in the shoots and roots increased, while the level of iPA in the shoots decreased, and in the roots increased 5.8- and 3.1-fold, respectively. However, the levels of all CK forms in the organs of stress plants remained below controls (Fig. 7).

Dynamics and distribution of endogenous cytokinins in the organs of 18-day-old plants of Triticum spelta L. cv. ‘Frankenkorn’ after moderate soil drought (four days without watering) and on the 21st day after recovery (ng/g DW). Differences between the mean values were evaluated using Bonferroni-corrected ANOVA, considered to be significant at P < 0.05; *P < 0.05; **P < 0.01; ***P < 0.001 compared to control at this stage of vegetation. Notes: t-Z—trans-zeatin, ZR—zeatin riboside; iPA—isopentenyladenosine, iP—isopentenyladenine, ZOG—zeatin-O-glucoside; traces—less than 0,5 ng/g DW
Discussion
Phytohormones play an important role in regulating physiological and metabolic processes, which are involved in plant adaptation to abiotic stresses. Adaptation to soil drought in wheat plants occurs through the maintenance of water balance, retardation of shoot growth, and enhancement of root system development (Kosakivska et al. 2018). We showed that morphometrically, plant organs of 18-day wheat and spelt plants after stress and 21-day-old plants following recovery were inferior to control plants. Roots were more sensitive to drought; however, root recovery in spelt was superior to that in wheat. ABA is recognized as the main stress signal, conducted from root to shoot, which suppresses leaf growth, induces changes in leaf hydrations and in nutrient content (Schachtman and Goodger 2008). Accumulation of endogenous ABA under drought conditions plays a significant role in supporting root growth (Giuliani et al. 2005), hair formation, and elongation (Chen et al. 2006; Xu et al. 2013). Following moderate soil drought, we noted an increase in endogenous ABA content in wheat and spelt organs. The hormone was accumulated in wheat roots and spelt shoots. Following recovery, it dominated hormone in shoots of both plants, with wheat cv ‘Podolyanka’ displaying significantly higher ABA content compared to spelt cv ‘Frankenkorn.
Zhang and colleagues (2020) reported that in rice, ABA and IAA were involved in the formation of root and root hair response to water deficits. It is also known that the accumulation of auxins in the root system of wheat plants contributed to a reduction in water consumption, both during the day and at night, and strengthened hydraulic properties of plants, enabling them to maintain water supplies and increase yield in drought conditions (Sadok and Schoppach 2019). Auxins contribute to root branching and the development of drought resistance in tobacco (Verma et al. 2016; Wang et al. 2018). We demonstrated that endogenous IAA accumulation was occurring more actively in wheat plants under all conditions. Subject to moderate soil drought, endogenous IAA content in shoots and roots of 18-day-old wheat and spelt plants decreased. Following recovery, IAA content in 21-day-old plants failed remained below control levels. It is important that the content of the hormone was significantly higher in wheat cv ‘Podolyanka’ shoots, as other researchers have shown young leaves, characterized by the highest IAA content and its active de novo synthesis (Ljung et al. 2001), generally exhibit an increased stress tolerance (Mühlenbock et al. 2008).
Several studies reported that the content of endogenous gibberellins, which influence plant growth and enable adaptations, is altered through stress. Drought represses processes that are regulated by GA, including seed germination, growth of shoots (Munns and Tester 2008). Osmotic stress retards GA accumulation in maize leaves (Nelissen et al. 2018) and tomato plants (Shohat et al. 2021). Mutant lines with reduced GA content exhibit salt and drought resistance due to slower biosynthesis or faster degradation of the hormone (Achard et al. 2006; Llanes et al. 2016). Our results show that in wheat cv ‘Podolyanka’ and spelt cv ‘Frankenkorn’ moderate soil drought was associated with reductions in GA3 content. We obtained similar results when studying the effect of heat stress on GA3 accumulation and distribution in wheat and spelt plants. The level of GA3 decreased in shoots and roots of both plants (Kosakivska et al. 2021). In control conditions, spelt had a significantly higher level of gibberellins compared to wheat. Subject to stress conditions, the hormone dominated in wheat roots and spelt shoots. At the same time, root and shoot growth was throttled, however, in spelt roots were more dramatically impacted, while in wheat—shoots were. In both plants, gibberellin accumulated primarily in roots following recovery. GA3 was most abundant in wheat roots. After heat stress, an increase in GA3 content occurred in both species, but not to the levels measured in control plants. More active accumulation of GA3 was observed in the roots (Kosakivska et al. 2021).
Salicylic acid acts as a signaling molecule which activated the plant defense under stress (Wani et al. 2017). SA induced an increase in hydrogen peroxide content, which stimulated the biosynthesis of defensive antioxidant compounds and improved stress tolerance (Hara et al. 2012). Notably, changes in endogenous SA and ABA content in Brassica napus depended on the duration and intensity of drought stress. In the initial stress phase, SA and ABA levels rose. Later stages were characterized by activation of indirect ABA reaction, downregulation of SA synthesis genes, and a reduction in hormone level (Park et al. 2021). Water deficit was associated with a twofold increase of endogenous SA in barley roots (Bandurska and Stroiński 2005). We determined that during moderate soil droughts endogenous SA content in organs of 18-day-old wheat and spelt plants increased, more prominently in roots compared to shoots. At the same time, we previously observed a reduction in SA content in wheat, while in spelt SA content increased after heat stress (Kosakivska et al. 2021). In 21-day-old wheat plants following recovery from stress, SA content was similar to control conditions, while in spelt SA content was markedly higher than in control conditions, similar to what was observed after heat stress. These results provide indirect evidence that endogenous SA participates in the formation of a defensive reaction to soil drought, primarily in shoots of wheat and spelt.
When drought conditions are present, cytokinins regulate plant growth and stabilize photosynthetic processes (Rivero et al. 2009; Prerostova et al. 2018). Of note, development of drought tolerance was observed when both during increase and decrease of endogenous CKs content (Rivero et al. 2007; Werner et al. 2010; Nishiyama et al. 2011).
Prerostova et al (2018) reported that a reduction in endogenous CKs during droughts was accompanied by lower water loss rates in transgenic Arabidopsis plant leaves and improved drought tolerance. However, these plants are slower to recover after being watered. Elevated CKs content during drought in other Arabidopsis mutants resulted in significant loss of moisture. However, these plants were able to recover faster and more effectively.
Here, we show that significant remodeling of the CK pool takes place in wheat and spelt plants during the formation of a response reaction to moderate soil drought. Total CKs content in wheat cv ‘Podolyanka’ was threefold higher compared spelt cv ‘Frankenkorn’ under varied conditions. During growth, total content of CKs in control wheat plants diminished, while in spelt plants it increased. Moderate soil drought resulted in elevated total CKs content in both plants that we studied. In the recovery period, CKs pool in wheat dropped by 25,6% in comparison with 18-day-old stressed plants and was greater by 13.3% compared to control 21-day-old plants. On the other hand, total content of CKs in spelt during the recovery period rose and exceeded the content in 18-day-old stressed plants by 19%, while remaining 12,6% lower than in 21-day-old control plants (Table 1).
Previously, we demonstrated that following heat stress the pool of CKs significantly increased in wheat, while in spelt—decreased more than twofold. After recovery, in shoots of both plants the pool of CKs decreased, while in wheat roots it did not change, and in spelt it decreased. The total CKs content in stressed wheat plants was twice as high as in spelt (Kosakivska et al. 2021). Therefore, comparing the results obtained after heat stress and soil drought, we can state that changes in the accumulation and distribution of GA3, SA and CKs were specific to the studied species, and also depended on the type of stress.
In summary, our study showed that moderate soil drought induced rapid nonspecific and specific changes in hormone accumulation and distribution in wheat and spelt plants (Fig. 8). In wheat cv ‘Podolyanka’, an accumulation of endogenous ABA, SA and zeatin took place, which dominated in roots, whereas the content of IAA, GA3, t-ZOG тa iP decreased.

Changes in endogenous phytohormones balance recorded after moderate soil drought in 18-day-old plants of Triticum aestivum L., cv ‘Podolyanka’ and Triticum spelta L., cv ‘Frankenkorn’ and on 21st day (in comparison with moderate soil drought) after recovery
Following recovery, we observed increased levels of ABA, IAA, GA3, SA, t-ZOG and decreased levels of t-Z and iP. In stressed spelt plants, an accumulation of ABA and SA took place in shoots, while an increase in ABA, CK and SA content and a decrease in IAA, GA3 levels took place in roots. Following recovery, we observed increased levels of IAA, GA3 and CK in shoots. Wheat plants retained high levels of ABA and SA following recovery, but spelt only ABA. These fluctuations in the patterns of accumulation, localization, and ratios of specific classes of phytohormones suggest a direct involvement of endogenous abscisic acid, indole-3-acetic acid, cytokinins, gibberellic and salicylic acids in the formation of wheat and spelt responses to soil drought and may be useful in screening of stress-resistant genotypes of cereals, taking future climatic changes into account.
Abbreviations
- ABA:
-
Abscisic acid
- IAA:
-
Indole-3-acetic acid
- GA3 :
-
Gibberellic acid
- SA:
-
Salicylic acid
- CKs:
-
Cytokinins
- iP:
-
Isopentenyladenine
- iPA:
-
Isopentenyladenosine
- t-Z:
-
trans-Zeatin
- t-ZR:
-
trans-Zeatin riboside
- t-ZOG:
-
trans-Zeatin-O-glucoside
- FW:
-
Fresh weight
- DW:
-
Dry weight
References
Achard P, Cheng H, De Grauwe L, Decat J, Schoutteten H, Moritz T, Van Der Straeten D, Peng J, Harberd NP (2006) Integration of plant responses to environmentally activated phytohormonal signals. Science 311(5757):91–94. https://doi.org/10.1126/science.1118642
Babenko LM, Hospodarenko HM, Rozhkov RV, Pariy YF, Babenko AV, Kosakivska IV (2018) Triticum spelta: origin, biological characteristics and perspectives for use in breeding and agriculture. Regul Mech Biosyst 9(2):250–257. https://doi.org/10.15421/021837 (in Ukrainian)
Bandurska H, Stroiński A (2005) The effect of salicylic acid on barley response to water deficit. Acta Physiol Plant 27:379–386. https://doi.org/10.1007/s11738-005-0015-5
Chen C-W, Yang Y-W, Lur HS, Tsai Y-G, Chang M-C (2006) A novel function of abscisic acid in the regulation of rice (Oryza sativa L.) root growth and development. Plant Cell Physiol 47(1):1–13. https://doi.org/10.1093/pcp/pci216
Daszkowska-Golec A, Szarejko I (2013) The molecular basis of ABA-mediated plant response to drought. In: Vahdati K, Leslie C (eds) Abiotic stress—plant responses and applications in agriculture. pp 103–133. https://doi.org/10.5772/53128
Dietrich D (2018) Hydrotropism: how roots search for water. J Exp Bot 69(11):2759–2771. https://doi.org/10.1093/jxb/ery034
Escarnot E, Jacquemin J-M, Agneessens R, Paquot M (2012) Comparative study of the content and profiles of macronutrients in spelt and wheat, a review. Biotechnol Agron Soc Environ 16(2):243–256
Estrada-Melo AC, Ma C, Reid MS, Jiang CZ (2015) Overexpression of an ABA biosynthesis gene using a stress-inducible promoter enhances drought resistance in petunia. Hortic Res 2:15013. https://doi.org/10.1038/hortres.2015.13
Giuliani S, Sanguineti MC, Tuberosa R, Bellotti M, Salvi S, Landi P (2005) Root-ABA1, a major constitutive QTL, affects maize root architecture and leaf ABA concentration at different water regimes. J Exp Bot 56(422):3061–3070. https://doi.org/10.1093/jxb/eri303
Gupta A, Rico-Medina A, Cano-Delgado AI (2020) The physiology of plant responses to drought. Science 386(6488):266–269. https://doi.org/10.1126/science.aaz7614
Hara M, Furukawa J, Sato A, Mizoguchi T, Miura K (2012) Abiotic stress and role of salicylic acid in plants. In: Ahmad P, Prasad M (eds) Abiotic stress responses in plants. Springer, New York, pp 235–252. https://doi.org/10.1007/978-1-4614-0634-1_13
Hu W, Ren T, Meng F, Cong R, Li X, White PJ, Lu J (2019) Leaf photosynthetic capacity is regulated by the interaction of nitrogen and potassium through coordination of CO2 diffusion and carboxylation. Physiol Plant 167(3):418–432. https://doi.org/10.1111/ppl.12919
Iqbal S, Wang X, Mubeen I, Kamran M, Kanwal I, Díaz GA, Abbas A, Parveen A, Atiq MN, Alshaya H, El-Abedin TKZ, Fahad S (2021) Phytohormones trigger drought tolerance in crop plants: outlook and future perspectives. Front Plant Sci 12:799318. https://doi.org/10.3389/fpls.2021.799318
Khalvandi M, Siosemardeh A, Roohi E, Keramati S (2021) Salicylic acid alleviated the effect of drought stress on photosynthetic characteristics and leaf protein pattern in winter wheat. Heliyon 7(1):e05908. https://doi.org/10.1016/j.heliyon.2021.e05908
Kosakivska IV, Vasyuk VA, Voytenko LV (2018) Drought stress effects on growth characteristics of two relative wheats Triticum aestivum L. and Triticum spelta L. Fisiol Rast Genet 50(3):241–252. https://doi.org/10.15407/frg2018.03.241 (in Ukrainian)
Kosakivska IV, Vasyuk VA, Voytenko LV, Shcherbatiuk MM (2021) Changes in hormonal status of winter wheat (Triticum aestivum L.) and spelt wheat (Triticum spelta L.) after heat stress and in recovery period. Cereal Res Commun (IF1.240) 50:821. https://doi.org/10.1007/s42976-021-00206-5
Kosakivska IV, Voytenko LV, Vasyuk VA, Shcherbatiuk MM (2022) Effect of pre-sowing priming of seeds with exogenous abscisic acid on endogenous hormonal balance of spelt wheat under heat stress. Zemdirb Agric 109(1):21–26. https://doi.org/10.13080/z-a.2022.109.003
Lacko-Bartosova N, Korczyk-Szabo J, Razny R (2010) Triticum spelta—a specialty grain for ecological farming systems. Res J Agricult Sci 42(1):143–147
Lind C, Dreyer I, López-Sanjurjo EJ, von Meyer K, Ishizaki K, Kohchi T, Lang D, Zhao Y, Kreuzer I, Al-Rasheid KAS, Ronne H, Reski R, Zhu J-K, Geiger D, Hedrich R (2015) Stomatal guard cells co-opted an ancient ABA-dependent desiccation survival system to regulate stomatal closure. Curr Biol 25(7):928–935. https://doi.org/10.1016/j.cub.2015.01.067
Ljung K, Bhalerao RP, Sandberg G (2001) Sites and homeostatic control of auxin biosynthesis in Arabidopsis during vegetative growth. Plant J 28(4):465–474. https://doi.org/10.1046/j.1365-313x.2001.01173.x
Llanes A, Andrade A, Alemano S, Luna V (2016) Alterations of endogenous hormonal levels in plants under drought and salinity. Am J Plant Sci 7(9):1357–1371. https://doi.org/10.4236/ajps.2016.79129
Morgun VV, Dubrovna OV, Morgun BV (2016) Suchasni biotehnologiji otrymannia stijkyh do stresiv roslyn pshenyci [Modern biotechnologies of obtaining stress-resistant wheat plants]. Fiziologija Rastenij i Genetika 48:196–214 (in Ukrainian)
Mühlenbock P, Szechy’nska-Hebda M, Płaszczyca M, Baudo M, Mateo A, Mullineaux PM, Parker JE, Karpinska B, Karpinski S (2008) Chloroplast signaling and LESION SIMULATING DISEASE1 regulate crosstalk between light acclimation and immunity in Arabidopsis. Plant Cell 20(9):2339–2356. https://doi.org/10.1105/tpc.108.059618
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Ann Rev Plant Biol 59(1):51–81. https://doi.org/10.1146/annurev.arplant.59.032607.092911
Nelissen H, Sun XH, Rymen B, Jikumaru Y, Kojima M, Takebayashi Y, Abbeloos R, Demuynck K, Storme V, Vuylsteke M, De Block J, Herman D, Coppens F, Maere S, Kamiya Y, Sakakibara H, Beemster GTS, Inzé D (2018) The reduction in maize leaf growth under mild drought affects the transition between cell division and cell expansion and cannot be restored by elevated gibberellic acid levels. Plant Biotechnol J 16(2):615–627. https://doi.org/10.1111/pbi.12801
Nishiyama R, Watanabe Y, Fujita Y, Le DT, Kojima M, Werner T, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Kakimoto T, Sakakibara H, Schmülling T, Tran L-SP (2011) Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and ABA responses, and ABA biosynthesis. Plant Cell 23(6):2169–2183. https://doi.org/10.1105/tpc.111.087395
Park S-H, Lee B-R, La VH, Mamun MA, Bae D-W, Kim T-H (2021) Characterization of salicylic acid- and abscisic acid-mediated photosynthesis, Ca2+ and H2O2 accumulation in two distinct phases of drought stress intensity in Brassica napus. Environ Exp Bot 186:e104434. https://doi.org/10.1016/j.envexpbot.2021.104434
Prerostova S, Dobrev PI, Gaudinova A, Knirsch V, Körber N, Pieruschka R, Fiorani F, Brzobohatý B, Cerný M, Spichal L, Humplik J, Vanek T, Schurr U, Vankova R (2018) Cytokinins: their impact on molecular and growth responses to drought stress and recovery in Arabidopsis. Front Plant Sci 9:655. https://doi.org/10.3389/fpls.2018.00655
Rivero RM, Kojima M, Gepstein A, Sakakibara H, Mittler R, Gepstein S, Blumwald E (2007) Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc Natl Acad Sci USA 104(49):19631–19636
Rivero RM, Shulaev V, Blumwald E (2009) Cytokinin-dependent photorespiration and the protection of photosynthesis during water deficit. Plant Physiol 150(3):1530–1540. https://doi.org/10.1104/pp.109.139378
Sadok W, Schoppach R (2019) Potential involvement of root auxins in drought tolerance by modulating nocturnal and daytime water use in wheat. Ann Bot 124(6):969–978. https://doi.org/10.1093/aob/mcz023
Sakhabutdinova AR, Fatkhutdinova DR, Bezrukova MV, Shakirova FM (2003) Salicylic acid prevents the damaging action of stress factors on wheat plant. Bulg J Plant Physiol 314–319 (special edition)
Schachtman DP, Goodge JQ (2008) Chemical root to shoot signaling under drought. Trends Plant Sci 13(6):281–287. https://doi.org/10.1016/j.tplants.2008.04.003
Shohat H, Ceriker H, Vasuki H, Illouz-Eliaz N, Blum S, Amsellem Z, Tarkowska D, Aharoni A, Eshed Y, Weiss D (2021) Inhibition of gibberellin accumulation by water deficiency promotes fast and long-term ‘drought avoidance’ responses in tomato. New Phytol 232(5):1985–1998. https://doi.org/10.1111/nph.17709
Ullah A, Manghwar H, Shaban M, Khan AH, Akbar A, Ali U, Ali E, Fahad S (2018) Phytohormones enhanced drought tolerance in plants: a coping strategy. Environ Sci Pollut Res Int 25(33):33103–33118. https://doi.org/10.1007/s11356-018-3364-5
Van Emden H (2008) Statistics for terrified biologists. Wiley-Blackwell, Oxford, UK
Verma V, Ravindran P, Kumar PP (2016) Plant hormone-mediated regulation of stress responses. BMC Plant Biol 16:86. https://doi.org/10.1186/s12870-016-0771-y
Wang C, Zhao Y, Gu P, Zou F, Meng L, Song W, Yang Y, Wang S, Zhang Y (2018) Auxin is involved in lateral root formation induced by drought stress in tobacco seedlings. J Plant Growth Regulat 37(3):539–549. https://doi.org/10.1007/s00344-017-9752-0
Wani AB, Chadar H, Wani AH, Singh S, Upadhyay N (2017) Salicylic acid to decrease plant stress. Enviorn Chem Lett 15(1):101–123. https://doi.org/10.1007/s10311-016-0584-0
Werner T, Nehnevajova E, Kollmer I, Novak O, Strnad M, Kramer U, Schmulling T (2010) Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 22(12):3905–3920. https://doi.org/10.1105/tpc.109.072694
Xie Z, Jiang D, Cao W, Dai T, Jing Q (2003) Relationships of endogenous plant hormones to accumulation of grain protein and starch in winter wheat under different post-anthesis soil water statuses. Plant Growth Regul 41(2):117–127. https://doi.org/10.1023/A:1027371906349
Xu W, Jia L, Shi W, Liang J, Zhou F, Li Q, Zhang J (2013) Abscisic acid accumulation modulates auxin transport in the root tip to enhance proton secretion for maintaining root growth under moderate water stress. New Phytol 197(1):139–150. https://doi.org/10.1111/nph.12004
Zhang Y, Du H, Gui Y, Xu F, Liu J, Zhang J, Xu W (2020) Moderate water stress in rice induces rhizosheath formation associated with abscisic acid and auxin responses. J Exp Bot 71(9):2740–2751. https://doi.org/10.1093/jxb%2Feraa021
Acknowledgements
This publication contains the results of research conducted within the project funded by the National Academy of Sciences of Ukraine III-90-19.463 "Hormonal regulation of growth and development of cereals under the influence of negative climatic factors" (2019-2023).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Human and animal rights
This article does not contain any studies involving animals or human participants performed by any of the authors.
Additional information
Communicated by Gabor Kocsy.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kosakivska, I.V., Vasyuk, V.A., Voytenko, L.V. et al. The effects of moderate soil drought on phytohormonal balance of Triticum aestivum L. and Triticum spelta L.. CEREAL RESEARCH COMMUNICATIONS 51, 627–638 (2023). https://doi.org/10.1007/s42976-022-00332-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42976-022-00332-8