
Abstract
This study aimed to investigate the effect of salinity and water stress on the physiological and functional characteristics of winter wheat (Triticum aestivum L.) under the foliar application of plant growth regulators (PGRs). The experiment was carried out as a split plot based on a randomized complete block design with three replications in two environments. In each environment, water stress at two irrigation levels (after 90 and 120 mm of pan evaporation) and with two EC of 1.5 and 10 dS/m in the main plots and spraying of PGRs including salicylic acid (SA), gibberellic acid (GA3), and cytokinins (CK) (purine) content with a concentration of 100 ppm and the control treatment (spraying solution with normal water) were placed in subplots. Results indicated that all treatments caused significant increases in functional and qualitative characteristics and yield of Triticum aestivum L. The saline environment and irrigation level after 120 mm of pan evaporation caused a reduction in grain yield in all traits except for seed proline, seed nitrogen content, and seed protein content. Also, the combined foliar application of GA3 + CK + SA increased yield in most traits. The highest RWC of flag leaves was observed in the foliar application of GA3 + SA (3.36 kg/ha) and then in the foliar application of GA3 + SA + CK (57.87 kg/ha). GA3 interacts with PGR spraying to balance another development under saline and non-saline conditions.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Agricultural science requires the most research in the field of crop adaptation to climate change in the current situation. Winter wheat (Triticum aestivum L.) is one of the most important crops in terms of cultivated area and production rate in the world and plays an important role in providing the food needs of human societies (Kinga et al. 2020). Among the environmental factors, saline stress has received serious attention from researchers in Iran for about 50 years. About eight million hectares of irrigated land are used to produce about 12% of Iran's produce. About 6.8 million hectares of these 8 million hectares have various restrictions, including salinity (Roozitalab et al. 2018). The salinization of water and soil resources has increased attention to research related to ways to improve the yield of agricultural plants in saline conditions (Pirasteh-Anosheh et al. 2015). Salinity causes the phenomenon of physiological dryness and directly damages the internal structure of the plant (El Karamany et al. 2019). In arid and semiarid regions, reducing the quantity and quality of water is one of the issues limiting production (Mosaffa and Sepaskhah 2019). Water stress is another important factor limiting the production of agricultural products, which strongly affects food security (Zhang et al. 2018). It also affects leaf water potential, photosynthetic pigments, stomatal conductance, and absorption of elements, especially phosphorus and nitrogen (Anli et al. 2020; Gowthami 2022; Samad et al. 2023).
Plant growth regulators (PGRs) are one of the solutions to increase the yield of agricultural plants under different stress conditions (Zhuang et al. 2019; Correia et al. 2020; Desta and Amare 2021). Its yield depends on the intensity, genotype, time of application, concentration used, and intensity of the stress level (Quamruzzaman et al. 2021). PGRs include salicylic acid (SA), gibberellic acid (GA3), and cytokinins (CK). GA3 has various effects on plants, inducing grain germination, and stimulating the production of important hydrolyzing enzymes in the germination of leguminous grains (Nagar et al. 2021). Chauhan et al. (2019), Al-Harthi et al. (2021), Ali et al. (2021), and Attia et al. (2022) certified the alleviating effect of GA3 on plant tolerance to salt stress. SA, as one of the most important phytohormones, plays a crucial and emerging role in plants' response to salt stress and supports them against many water stresses. Ma et al. (2017), Xu et al. (2022), and Yang et al. (2023) revealed the protective roles of SA priming in plants' response to salt stress. CK reduces the adverse effects of non-living stresses and also causes cell division, removal of apical dominance, stem differentiation, and delaying aging (Veselov et al. 2017). Hai et al. (2020) acknowledged the role of CK in plant response to water stress.
The vast expanse of dry and salty lands in Isfahan province, especially in the northern plains of Golpayegan city, are harvested in a barren form or with a small area and a negligible yield. Water salinity in the region is one of the limiting factors for agricultural activities, which if reduced and adjusted with the help of growth regulators, will help to increase productivity per unit area, reduce unemployment and job creation, reverse migration, and ultimately increase economy growth at the level of the region and the country. Until now, there has been no solution to the problem mentioned, which was a research gap and the main objective of this study. Modifying the structure of the soil in a large area is practically impossible. In the current situation, it is vital to adopt agricultural management methods and the use of PGRs. The combined use of several PGRs on wheat plants in different environments and with different types and levels of stress, which have not yet experienced any type of PGRs, is an innovative method. In general, investigating the impact of biological stresses on the productivity and quality of crops and also determining the appropriate solution to reduce the negative effects of stresses and improve the quality of the produced product can be considered an effective step in the direction of improving and developing agricultural plants, including Triticum aestivum L. The climate of the Golpayegan region (relatively cold winters (Tmin = −21 °C) and hot and dry summers (Tmax = + 37 °C) and a mean rainfall of about 300 mm) is completely suitable for wheat cultivation. Therefore, the results of this research can be generalized to other places where this product is cultivated. The hypotheses of this research can be stated as follows: a) Foliar spraying with PGRs can cause resistance to environmental stresses (salinity and drought); b) the interaction of PGRs can reduce the effects of environmental stresses (salinity and drought); and c) environmental stresses (salinity and drought) can have a negative effect on the yield and yield components of the wheat plant. This experiment aimed to determine how PGRs affect the functional and qualitative characteristics of winter wheat (Triticum aestivum L.) under conditions of saline and water stress. Also, obtaining the best application rate of these regulators in creating resistance in wheat plants by spraying is another goal of this study.
Materials and methods
Experimental environment
Experiments were carried out in the crop year of 2022–2023 in two non-saline environments (irrigation with EC of 1.5 dS/m) at the agricultural and animal science research station with latitude (33° 28′ north) and longitude (50° 17′ east) and saline environment (irrigation with EC of 10 dS/m) in the fields of the northern plain of Golpayegan city with latitude (33° 30′ north) and longitude (50° 23′ east). The study area represents a semiarid climate based on the Koppen (1936) method. Its annual average temperature and rainfall are 18 °C and 300 mm, respectively (Table 1).
Agricultural operations
Soil samples were taken from the plot of land located in the agricultural research station and farms before planting. To conduct the soil test, 10 samples were taken randomly, one kilogram each, and then the soil samples were completely mixed, and a 1 kg sample was selected as a soil sample and sent to the soil science laboratory of Isfahan Agricultural Research Center. Table 2 presents the physicochemical characteristics of the soil at a depth of 0–30 cm. To strengthen the soil and provide the elements required by the plant and according to the result of the soil fertilizer test, before planting, all the required phosphorus fertilizers were added to the soil from the triple superphosphate source at the rate of (non-saline field: 170 kg/ha; saline field: 190 kg/ha), potash fertilizer in the form of potassium sulfate at the rate (non-saline field: 150 kg/ha; saline field: 170 kg/ha), and one-third of nitrogen fertilizer in the form of urea at the rate of (non-saline field: 120 kg/ha; saline field: 150 kg/ha). The planting operation was done with a density of 400 plants per hectare in the non-saline field on October 23, 2021, and in the salt field on December 6, 2021.
Experimental design and treatments
The experiment was carried out as a split plot based on a randomized complete block design (RCBD) with three replications in two environments. Drought stress was executed at three irrigation levels after evaporation of 90 and 120 mm from Class A evaporation pan as non-stress and severe stress, respectively. In each environment, water stress at two irrigation levels (after 90 and 120 mm of pan evaporation) in the main plots and spraying of PGRs SA, GA3, and CK (purine) with a concentration of 100 ppm, and the control treatment (spraying solution with normal water) were placed in subplots. PGR spraying was done in the pollination stage (to reduce the effects of heat stress on the results of flowering and grain formation of this stage). Foliar application was done with PGRs including GA3, SA, and CK by spraying in two stages: 1—double ridge, 2—terminal spikelet stage in the amount of 25cc of PGRs solution in 5 L of water per plot at the beginning of the morning. The measured traits include the relative water content (RWC) of flag leaves (Soltys-Kalina et al. 2016), proline content in flag leaves, the no. of spikes/m2, the no. of spikelet and grains per spike, the thousand seed weight (PMG), biological yield (Yb), and seed content (nitrogen, protein, and gluten-free) (Fariñas et al. 2019).
Sampling and measurement
Sampling was done in five steps. The first stage of sampling was done in the non-salinity field 135 days after germination and in the saline field 75 days after germination. The second stage and then every 15 days, and in the final harvesting stage, a sampling stage was done. Sampling was done by a 1 × 1 square meter, and the samples were placed in paper envelopes with a code written for each one. After measuring the traits, the envelopes were placed in an oven at 75 °C for 72 h and the results were recorded by a digital scale with a sensitivity of one hundredth. Also, in the final harvest, 1m2 from the middle rows of each plot was removed by removing the marginal effect from the soil surface and the desired traits were measured.
Statistical analysis
All the obtained data were analyzed using statistical software SAS version 9.2 and the MSTAT-C Program version 7.0.1. To compare the means, Duncan's multiple range tests (DMRT) were used at the 5% level. STATGRAPHICS software version 18.1.001 was used to obtain correlation coefficients. Excel software was used to draw graphs. Bartlett's test was used for the combined analysis of traits and the possibility of combining the two environments (Table 3).
Results
The relative water content of flag leaves
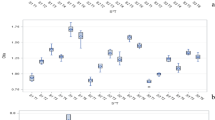
The interaction effect of environment × irrigation levels × PGRs on the RWC of the flag leaf was significant at the 5% probability level (Table 4). In the current research, the highest RWC of flag leaves was obtained in the condition of non-saline, irrigation level after 90 mm of pan evaporation, and PGRs including GA3 + CK + SA at the rate of 74.77%. Its difference with other foliar applications was significant except for CK + SA and GA3 + SA (Fig. 1). The interaction effect of environmental levels × PGRs caused a 29% increase in the RWC of the flag leaf.

Interaction effect of environment × irrigation levels × PGRs on relative leaf water content in both saline and non-saline environments
Proline content in flag leaves
The interaction effect of environmental levels × PGRs on the proline content of flag leaves was significant at the 5% level (Table 4). The highest content of proline in flag leaf was obtained in the conditions of a saline environment, irrigation level after 120 mm of pan evaporation, and PGRs including foliar application of GA3 + CK + SA at the rate of 18.59 mg/g. The difference between this treatment with a non-saline environment, irrigation level after 90 mm of pan evaporation, and other PGRs and no PGRs was significant except for foliar application of GA3 + SA (Fig. 2). The interaction effect of environment × irrigation levels × PGRs caused a 69% increase.

Interaction effect of environment × irrigation levels × PGRs on flag leaf proline content in both saline and non-saline environments
The number of spikes per square meter
The interaction effect of environment × irrigation levels on the no. of spikes/m2 was significant at the 5% probability level (Table 4). The highest no. of spikes/m2 was obtained in non-salinity, and the irrigation level after 90 mm of pan evaporation was equal to 436.26. It was significantly different from other treatments (Table 7). The interaction effect of environment × irrigation levels caused a 30% increase in the no. of spikes/m2. The interaction effect of environment × PGRs on the no. of spikes/m2 was significant at the 5% probability level (Table 4). The highest no. of spikes/m2 was obtained in non-salinity and PGRs including GA3 + CK + SA at the rate of 452.19. It was significantly different from other treatments (Table 8). The interaction of environment × PGRs caused a 43% increase in the no. of spikes/m2.
No. of spikelet per spike
The effect of the environment on the no. of spikelet per spike was significant at the probability level of one percent (Table 4). The maximum no. of spikelet in the spike was obtained in the condition of non-salinity equal to 20.21. It was significantly different from the salinity environment (Table 5). Saline stress caused a 5% decrease in the no. of spikelet per spike. The effect of PGRs on the no. of spikelet per spike was significant at the 5% probability level (Table 4). The highest no. of spikelet per spike was obtained by PGRs including GA3 + CK + SA at the rate of 20.63. It showed a significant difference with other PGRs and non-foliar spraying treatments (Table 5). PGRs including GA3 + CK + SA increased the no. of spikelet per spike by 14% compared to other foliar application treatments and the control treatment.
The number of grains per spikelet
The effect of PGRs on the number of grains per spikelet was significant at the 5% probability level (Table 4). The highest number of grains per spikelet was obtained by PGRs including GA3 + CK and GA3 + CK + SA, equal to 55.19 and 55.05, respectively. This treatment, except for GA application, showed a significant difference from other PGRs and non-foliar spraying treatments (Table 5). PGRs including GA3 + CK and GA3 + CK + SA increased the no. of grains per spike by 25% compared to the control treatment. The interaction effect of environment × irrigation levels on the no. of grains per spike was significant at the 5% probability level (Table 4). The highest no. of grains per spike in the condition of no salinity and irrigation level after 90 mm of pan evaporation was obtained at the rate of 59.00, and the difference with other treatments was significant (Table 6). The interaction of environment × irrigation levels increased the no. of grains per spike by 28%.
The thousand seed weight (PMG)
The interaction effect of environment × irrigation levels on PMG was significant at the 5% probability level (Table 4). The maximum PMG in the condition of no salinity and irrigation level after 90 mm of pan evaporation was obtained at the rate of 41.71 gr (Table 6). The interaction of environment × irrigation levels increased the RWC of flag leaves by 20%. The interaction effect of irrigation levels × PGRs on PMG was significant at the 5% probability level (Table 4). The highest weight of PMG was obtained at the irrigation level after 90 mm of pan evaporation and foliar application with PGRs including GA3 + CK at the rate of 40.90 gr (Table 7). The interaction between irrigation levels and PGRs caused an 18% increase in PMG.
Biological yield
The interaction effect of environment × irrigation levels on Yb was significant at the 5% probability level (Table 4). The highest Yb was achieved in the absence of salinity and the irrigation level after 90 mm of pan evaporation at the rate of 22,625.33 kg/ha, which was significantly different from the saline and irrigation environment at the irrigation level after 120 mm of pan evaporation (Table 5). The interaction of environment × irrigation levels increased the Yb by 35%. The interaction effect of irrigation levels × PGRs on Yb was significant at the probability level of 1% (Table 4). The highest Yb was achieved under irrigation conditions at the irrigation level after 90 mm of pan evaporation and PGRs including GA3 + CK + SA at the rate of 22,114.5 kg/ha. The difference between this treatment with the irrigation level after 120 mm of pan evaporation and other treatments of foliar spraying and no foliar spraying was significant (Table 8). The irrigation levels × PGRs increased the Yb by 38%. The foliar application of SA and GA (100 ppm) priming led to an increase in Yb under full irrigation conditions.
Grain nitrogen percentage
The effect of the environment on grain nitrogen percentage was significant at the probability level of 1% (Table 4). The highest percentage of grain nitrogen in saline conditions was 4.09%, which was significantly different from the non-saline environment (Table 5). Saline stress caused an increase of 18% in grain nitrogen. The interaction effect of irrigation levels × PGRs was significant at the 5% probability level (Table 4). The highest percentage of grain nitrogen at the irrigation level after 120 mm of pan evaporation was achieved by evaporation and PGRs including GA3 + CK + SA at the rate of 5.04%. This treatment showed a significant difference between evaporation and other foliar spraying and non-foliar spraying treatments with irrigation level after 90 mm of pan evaporation (Table 8). The irrigation level after 120 mm of pan evaporation and PGRs including GA3 + CK + SA increased grain nitrogen by 63%.
Grain protein percentage
The effect of the environment on grain protein percentage was significant at the probability level of one percent (Table 4). The highest percentage of grain protein was obtained in saline conditions at the rate of 23.42%, which was significantly different from the non-saline environment (Table 5). The results show that saline stress increased grain protein by 19%. The interaction effect of irrigation levels + PGRs was significant at the 5% probability level (Table 4). The highest percentage of grain protein was obtained at the irrigation level after 120 mm of pan evaporation, and foliar spraying of PGRs including GA3 + CK + SA at the rate of 28.83%. This showed a significant difference between evaporation and other foliar spraying and non-foliar spraying treatments with an irrigation level after 90 mm of pan evaporation (Table 8). The irrigation level after 120 mm of pan evaporation and PGRs including GA3 + CK + SA caused a 63% increase in grain protein.
Grain gluten
The interaction effect of environment × irrigation levels × PGRs on grain gluten content was significant at the 5% probability level (Table 4). The highest amount of grain gluten was obtained in non-saline conditions, the irrigation level after 90 mm of pan evaporation, and PGRs including GA3 + CK + SA at the rate of 33.50%. Its difference with the non-saline environment level, the irrigation level after 120 mm of pan evaporation, and other foliar spraying and no foliar spraying treatments was significant (Fig. 3). The interaction effect of environment × irrigation levels × PGRs caused a 54% increase in grain gluten.

Interaction effect of environment × irrigation levels × PGRs on grain gluten percentage in both saline and non-saline environments
Discussion
The effect of the environmental stresses on the studied traits
The reduction of wheat grain yield in a saline environment is due to the reduction of three components: the no. of spikes, the number of grains per spikelet, and PMG (Zorb et al. 2018). In saline conditions, the acceleration of terminal spike growth reduces the no. of spikelets per spike. Also, the decrease in PMG in saline conditions can be justified by reducing the length of the grain-filling period. Water salinity, as a growth-inhibiting factor, with its negative oxidation effect, destroys the cell structure and reduces the osmotic potential of the plant tissue, affecting yield. RWC is the most appropriate method for measuring the amount of water in plant tissues. For this reason, its importance is greater than other methods. There is a positive and significant correlation between the RWC and the no. of spikes (r = 0.846**), the no. of spikes (r = 0.595**), the no. of grains (r = 0.755**), 1000-grain weight (r = 0.398**), Yb (r = 0.807**), gluten index (r = 0.783**), and flag leaf proline content (r = 0.520**) (Table 9). The no. of spikelets is dependent on suitable water and soil conditions, and the salinity condition reduces the no. of spikelets in the spike. In the yellow stage, the living flowers that will produce grains are mature and close to pollination. The fifth and higher flowers are aborted. At this stage, the no. of spikelets was determined to be fertile and will produce grains. A decrease in the irrigation level leads to an increase in water consumption efficiency (Mosaffa and Sepaskhah 2019). Also, by conducting another experiment, it was shown that the RWC of the flag leaves was 76% in the control treatment and 5.47% in the saline treatment (10 dS). In the present experiment, there was a significant correlation between the no. of spikes and the no. of spikelets (r = 0.745**), the no. of grains (r = 0.818**), and PMG (r = 0.412**), which indicates the trend between these traits (Table 10). The application of different irrigation levels and salinity treatments has caused spikelets to be infertile and also reduced the transfer of photosynthetic materials to grains. Salinity affects grain yield components depending on when the stress is applied to the plant (Mashi et al. 2018). The no. of grains is directly related to the environmental conditions, duration, quality, and time of the stress peak, which in this experiment is the transition from the vegetative phase to the reproductive phase and also the flowering time from the stress-sensitive stages of the plant in terms of grain formation. Therefore, salinity stress hurts the vegetative growth stages and thereby decreases the grain yield and its components.
The effect of irrigation levels
Triticum aestivum L. grain yield components include the no. of spikes/m2, the number of grains per spikelet, and PMG, which develop in a time sequence. Considering that PMG is the last component of grain yield development, if other grain yield components (no. of grains per spike and no. of spikes/m2) decrease due to water stress, increasing PMG can compensate for this deficiency. The no. of spikes/m2 had a direct positive effect (0.38) and an indirect effect (0.27) through PMG on plant grain yield under drought stress conditions (Torabian and Maghsoudi 2012). In water stress conditions, absorption and stabilization of CO2 are reduced due to the partial closing of the stomata or the reduction of their degree of opening, so the total amount of cultivated material for grain filling is reduced. However, water stress does not reduce the transfer of nitrogen from the leaves to the grain, and this causes an increase in grain protein (Sehgal et al. 2018). Desta and Amare (2021) showed that the spraying of PGRs increased the mentioned traits in both normal irrigation conditions and water stress conditions. Water stress lowers all the physiological activities of Triticum aestivum L. This is while it seems that the use of SA priming with a concentration (of 100 ppm) in the conditions of water stress has improved the condition of the plant by increasing the plant's resistance to stress. As a result, it has led to higher grain yield. With increasing stress intensity, the traits of the no. of grains per spike, PMG, leaf area index, Yb, and photosynthetic pigments decreased, while proline content, soluble protein content, and soluble carbohydrates increased (Gowthami 2022). In an experiment, the highest and lowest biological yield of winter wheat were observed in the treatments of 125% of winter wheat water requirement × 1.6 dS/m salinity and 50% of winter wheat water requirement × salinity 7.8 S/m, respectively. Poor treatment compared to superior treatment showed a decrease of about 65% in this trait.
The effect of PGRs
In the present experiment, the effect of PGRs separately and in combination caused an increase in the values of each of the studied traits. PGRs including GA3, CK, and SA priming according to the type of their influence on traits (especially in grains that need cold or light), improving the biochemical characteristics of the plant such as the content of soluble proteins, free proline, photosynthetic pigments, and the no. of plant hormones (Samad et al. 2023). These increments are similar to those reported by Kumar et al. (2018) and El Karamany et al. (2019), on different plant species. Also, as a result of increasing grain yield in saline stress conditions, it causes cell division, stimulates morphogenesis, stimulates the growth of lateral buds, and stimulates the growth of leaves in plants (Forghani et al. 2018; Kumar et al. 2018). These results agree that the foliar application of PGRs had an enhancement effect on growth, as reported in some of the findings of Forghani et al. (2018) and Kumar et al. (2018). PGRs can reduce the damages to a minimum by creating resistance or modulating adverse effects in stressful environments (El Karamany et al. 2019). It delays the leaf senescence in plants. Also, it includes organic compounds that in small amounts can modify the growth (Chaurasiya et al. 2014). Also, it increases the active period of grain growth (grain germination times), controls the vegetative growth, flowering, fruiting, and seed production in plants, and increases PMG and the no. of grains per plant. Finally, PGRs increase the crop grain yield (Dwivedi et al., 2014; Safdari et al. 2014). The increase in Triticum aestivum L. grains PMG with the use of CK has been reported in various studies (Zheng et al. 2016). Cell division is one of the most important roles of CK, and it seems that foliar application of CK has increased biological grain yield by increasing the cell division of vegetative organs, increasing photosynthesis, and increasing the durability of leaf surface index and grain weight. Increasing the Yb of Triticum aestivum L. with the use of CK has also been reported in other studies (Zaheer et al. 2019). The effect of SA priming in the treatment without saline had a lesser effect on the RWC of leaves (4%). In the case of 10dS salinity treatment, the effect of SA priming increased the RWC of leaves by 20%. The physiological efficiency of nitrogen increased with increasing consumption of SA priming. Also, the consumption of 6 mM SA priming with ratios of 124.6 and 132.1 g of total dry matter per gram of absorbed nitrogen had the highest physiological efficiency of nitrogen in both irrigation and non-irrigation conditions, respectively. Irrigation interruption conditions have caused the re-transfer of nitrogen to increase, and the physiological efficiency of nitrogen has increased to 113.95 g of total dry matter per gram of absorbed nitrogen. On the other hand, SA priming moderated the water stress by increasing the physiological efficiency and re-transfer of nitrogen, which improved the yield of barley grains under water stress conditions compared to full irrigation (Abhari and Radman 2020).
Conclusion
The results of this research indicated the negative effect of salinity and water stress on the physiological and functional traits of the Triticum aestivum L. plant. Increasing the irrigation water salinity to 10 dS/m and irrigation levels after evaporation of 120 mm led to a significant decrease in the RWC of the flag leaf, the no. of spikes/m2, the no. of spikelets per spike, the number of grains per spikelet, PMG, the biological yield, and the amount of grain gluten. It also decreased the proline content of flag leaves and the percentage of nitrogen and grain protein. Foliar application of different PGRs has a significant effect on the growth, photosynthetic pigments, yield, and yield attributing characters significantly. Based on our study, the spraying of PGRs in combination (GA3 × SA × CK, GA3 × CK, and GA3 × SA) with a recommended 100-ppm concentration was the most effective foliar application for increasing growth and yield of Triticum aestivum L. Due to the specialized effects of each sprayed in two stages: 1—double ridge and 2—terminal spikelet, all traits modify the effects of environmental stress and increase yield in traits. Interaction of environment × irrigation level × PGRs showed the most RWC of the flag leaf in the non-stress of salinity under the influence of the spraying of PGRs at the irrigation level after 90 mm of pan evaporation in GA3 + CK + SA was observed at 74.77 kg/ha. At the irrigation level after 120 mm of pan evaporation in GA3 + CK, GA3 + SA, and CK + SA, the amount of 70.51 kg/ha was observed. The highest RWC was influenced by the spraying of PGRs at the irrigation level after 90 mm of pan evaporation in the foliar application of GA3 + SA at 73.36 kg/ha. It was also observed at the irrigation level after 120 mm of pan evaporation in the foliar application of GA3 + SA + CK the amount of 57.87 kg/ha. Our results reveal the novel finding that GA3 interacts with the spraying of PGRs to balance another development under saline and non-saline conditions in Triticum aestivum L. According to the results, salicylic acid priming and PGR spraying can partially reduce the negative effects of deficient irrigation and soil salinity stress. In general, it seems that by using these compounds, it is possible to reduce the number of times of irrigation and, in addition to managing water resources, reduce the adverse effects of drought stress in plants. Since our results are based on the data from a single year, at least an additional year hassle of study is recommended. The results will be useful for the local farmers as well as future research initiatives. In the area where this experiment is carried out, salinity and drought stress are the two dominant limitations compared to other cases in the climatic conditions of this region. Therefore, carrying out drought stress in the region in the range of 90 and 120 mm of evaporation allows us to investigate the plant's reaction to these environmental stresses in saline and non-saline environments (control) under the influence of foliar spraying of PGRs. In general, the results showed that the spraying of PGRs can be effective in increasing the quantitative and qualitative yield of wheat by improving the production components and reducing the salinity effect of irrigation water, and it can be suggested to researchers and farmers.
Availability of data and materials
All data and materials as well as a software application or custom code support our published claims and comply with field standards.
References
Abhari A, Radman AR (2021) The effect of salicylic acid on the physiological efficiency of nitrogen and atmospheric water consumption in the condition of interruption of irrigation. Plant Prod 27(2):187–200
Abhari A, Radman AR (2020) Effect of salicylic acid on physiological N efficiency and water use efficiency of barley in cut irrigation condition. J Plant Prod Res 27(2):187–200
Al-Harthi MM, Bafeel SO, El-Zohri M (2021) Gibberellic acid and jasmonic acid improve salt tolerance in summer squash by modulating some physiological parameters symptomatic for oxidative stress and mineral nutrition. Plants (basel) 10(12):2768. https://doi.org/10.3390/plants10122768
Ali AYA, Ibrahim MEH, Zhou G et al (2021) Gibberellic acid and nitrogen efficiently protect early seedlings’ growth stage from salt stress damage in Sorghum. Sci Rep 11:6672. https://doi.org/10.1038/s41598-021-84713-9
Anli M, Baslam M, Tahiri A, Raklami A, Symanczik S, Boutasknit A, Meddich A (2020) Biofertilizers as strategies to improve photosynthetic apparatus, growth, and drought stress tolerance in the date palm. Front Plant Sci 11:516818
Attia H, Alamer K, Algethami B et al (2022) Gibberellic acid interacts with salt stress on germination, growth and polyamine gene expression in fennel (Foeniculum vulgare Mill) seedlings. Physiol Mol Biol Plants 28:607–622. https://doi.org/10.1007/s12298-022-01140-4
Chauhan A, AbuAmarah BA, Kumar A, Verma JS, Ghramh HA, Khan KA, Ansari MJ (2019) Influence of gibberellic acid and different salt concentrations on germination percentage and physiological parameters of oat cultivars. Saudi J Biol Sci 26(6):1298–1304. https://doi.org/10.1016/j.sjbs.2019.04.014
Chaurasiya J, Meena ML, Singh HD, Adarsh A, Mishra PK (2014) Effect of GA3 and NAA on growth and yield of cabbage (Brassica oleracea var Capitata L.) cv. Pride of India. Bioscan 9(3):1139–1141
Correia S, Queirós F, Ferreira H, Morais MC, Afonso S, Silva AP, Gonçalves B (2020) Foliar application of calcium and growth regulators modulate sweet cherry (Prunus avium L.) tree performance. Plants 9(4):410. https://doi.org/10.3390/plants9040410
Desta B, Amare G (2021) Paclobutrazol as a plant growth regulator. Chem Biol Technol Agric 8:1–15. https://doi.org/10.1186/s40538-020-00199-z
Dwivedi SK, Arora A, Singh SD, Singh GP, Nagar S, Kumar S (2014) PGRs improve carbohydrate metabolism and yield attributes in wheat (Triticum aestivum L.) under water deficit condition. J. Cereal Res 5(2)
El Karamany MF, Sadak MS, Bakry BA (2019) Improving quality and quantity of mungbean plant via foliar application of plant growth regulators in sandy soil conditions. Bull Natl Res Centre 43:61. https://doi.org/10.1186/s42269-019-0099-5
Fariñas MD, Jimenez-Carretero D, Sancho-Knapik D, Peguero-Pina JJ, Gil-Pelegrín E, Álvarez-Arenas TG (2019) Instantaneous and non-destructive relative water content estimation from deep learning applied to resonant ultrasonic spectra of plant leaves. Plant Methods 15:128. https://doi.org/10.1186/s13007-019-0511-z
Forghani AH, Almodares A, Ehsanpour A (2018) Potential objectives for gibberellic acid and paclobutrazol under salt stress in sweet sorghum (Sorghum bicolor [L.] Moench cv. Sofra). Appl Biol Chem 61:113–124. https://doi.org/10.1007/s13765-017-0329-1
Gowthami R (2022) Foliar application of plant growth regulators in bhendi and its influence over growth parameters under moisture stress (2022). Available at SSRN: https://doi.org/10.2139/ssrn.4158254
Hai NN, Chuong NN, Tu NHC, Kisiala A, Hoang XLT, Thao NP (2020) Role and regulation of cytokinins in plant response to drought stress. Plants (Basel) 9(4):422. https://doi.org/10.3390/plants9040422
Kinga M, Idzi S, Roman K, Jolanta Roman K (2020) Effect of foliar applied acetylsalicylic acid on wheat (Triticum aestivum L.) under field conditions. Agron 10:1–19
Koppen W (1936) Das geographische system der Klimate. In: Koppen W, Geger R (eds) Handbuch der Klimatologie. Gebruder Borntraeger, Berlin, pp 1–44
Kumar M, Sarangi A, Singh DK et al (2018) Modelling the grain yield of wheat in irrigated saline environment with foliar potassium fertilization. Agric Res 7:321–337. https://doi.org/10.1007/s40003-018-0310-1
Ma X, Zheng J, Zhang X, Hu Q, Qian R (2017) Salicylic acid alleviates the adverse effects of salt stress on Dianthus Superbus (caryophyllaceae) by activating photosynthesis, protecting morphological structure, and enhancing the antioxidant system. Front Plant Sci 8:600. https://doi.org/10.3389/fpls.2017.00600
Mashi AS, Galeshi S, Zeinali A, Nourinia A (2018) The effect of salinity stress on yield and yield components of four barley genotypes. Agric Sci Nat Resour 14(5):16–31
Mosaffa HR, Sepaskhah AR (2019) Performance of irrigation regimes and water salinity on winter wheat as influenced by planting methods. Agric Water Manag 216:444–456. https://doi.org/10.1016/j.agwat.2018.10.027
Nagar S, Singh VP, Arora A, Dhakar R, Singh N, Singh GP, Meena S, Kumar S, Shiv Ramakrishnan R (2021) Understanding the role of gibberellic acid and paclobutrazol in terminal heat stress tolerance in wheat. Front Plant Sci 12:692252. https://doi.org/10.3389/fpls.2021.692252
Pirasteh-Anosheh H, Emam Y, Sepaskhah AR (2015) Improving barley performance by proper foliar applied salicylic acid under saline conditions. Int J Plant Prod 9(3):467–486
Quamruzzaman MD, Nuruzzaman SM, ManikShabala S, Zhou M (2021) Improving performance of salt-grown crops by exogenous application of plant growth regulators. Biomolecules 11(6):788. https://doi.org/10.3390/biom11060788
Roozitalab MH, Siadat H, Farshad A (2018) The soils of Iran. World soils book series. Springer, Switzerland
Safdari M, Dardar A, Khaniki GB (2014) The Independent effect of time and hormonal concentration treatments on reproductive traits in onion. Indian J Fundam Appl Life Sci 4(4):3009–3015
Samad A, Shaukat K, Ansari M, Nizar M, Zahra N et al (2023) Role of foliar spray of plant growth regulators in improving photosynthetic pigments and metabolites in Plantago ovata (Psyllium) under salt stress–a field appraisal. Biocell 47(3):523–532
Sehgal A, Sita K, Siddique KHM, Kumar R, Bhogireddy S, Varshney RK, Hanumantha Rao B, Nair RM, Prasad PVV, Nayyar H (2018) Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front Plant Sci 9:1705. https://doi.org/10.3389/fpls.2018.01705
Soltys-Kalina D, Plich J, Strzelczyk-Żyta D, Śliwka J, Marczewski W (2016) The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ’Katahdin’-derived potato cultivars. Breed Sci 66(2):328–331. https://doi.org/10.1270/jsbbs.66.328
Torabian A, Maghsoudi K (2012) Study on the relationship between yield and yield components of wheat under normal irrigation and drought stress conditions by path analysis method. Agron J 104:47–53
Veselov DS, Kudoyarova GR, Kudryakova NV, Kusnetsov VV (2017) Role of cytokinins in stress resistance of plants. Russ J Plant Physiol 64(1):15–27
Xu L, Chen H, Zhang T, Deng Y, Yan J, Wang L (2022) Salicylic acid improves the salt tolerance capacity of Saponaria officinalis by modulating its photosynthetic rate, osmoprotectants, antioxidant levels, and ion homeostasis. Agronomy 12(6):1443. https://doi.org/10.3390/agronomy12061443
Yang W, Zhou Z, Chu Z (2023) Emerging roles of salicylic acid in plant saline stress tolerance. Int J Mol Sci 24(4):3388. https://doi.org/10.3390/ijms24043388
Zaheer MS, Raza MAS, Saleem MF, Erinle KO, Iqbal R, Ahmad S (2019) Effect of rhizobacteria and cytokinins application on wheat growth and yield under normal vs drought conditions. Commun Soil Sci Plant Anal 50(20):2521–2533
Zhang J, Zhang S, Cheng M, Jin J (2018) Effect of drought on agronomic traits of rice and wheat: a meta-analysis. Int J Environ Res Public Health 15(5):839–853
Zheng C, Zhu Y, Wang C, Guo T (2016) Wheat grain yield increase in response to pre-anthesis foliar application of 6-benzyl amino purine is dependent on floret development. PLoS ONE 11(6):1–14
Zhuang L, Ge Y, Wang J, Yu J, Yang Z, Huang B (2019) Gibberellic acid inhibition of tillering in tall fescue involving crosstalks with cytokinins and transcriptional regulation of genes controlling axillary bud outgrowth. Plant Sci 287:110168. https://doi.org/10.1016/j.plantsci.2019.110168
Zorb C, Geilfus CM, Dietz KJ (2018) Salinity and crop yield. Plant Biol 21(S1):31–38
Acknowledgements
We are grateful to all of those with whom we have had the pleasure to work during this and other related research.
Funding
The authors did not receive support from any organization for the submitted work.
Author information
Authors and Affiliations
Contributions
Hamid Reza Tajdari and Ali Soleymani conceived and designed the study. Ali Soleymani and Nosratolah Montajabi conducted data gathering. Mohammad Reza Naderi Darbaghshahi performed statistical analyses. Hamid Reza Javanmard, Ali Soleymani, and Naderi Darbaghshahi wrote the article.
Corresponding author
Ethics declarations
Conflict of interest
There is no conflict of interest among authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tajdari, H.R., Soleymani, A., Montajabi, N. et al. The effect of foliar application of plant growth regulators on functional and qualitative characteristics of wheat (Triticum aestivum L.) under salinity and drought stress conditions. Appl Water Sci 14, 126 (2024). https://doi.org/10.1007/s13201-024-02203-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13201-024-02203-5