
Abstract
Purpose A long-term no-till study was conducted to determine whether offseason intercropping systems with maize as the cash crop increase crop straw inputs and the yield of soybean as the main crop and alter soil P dynamics compared with monocropped offseason maize. Methods The experiment was conducted between 2019 and 2021 in a randomized complete block design with four replications and four cropping systems in the offseason (fall–winter): maize alone; maize + palisade grass (U. brizantha); maize + sunn hemp (C. spectabilis); and maize + palisade grass + sunn hemp. Leaf P content, crop straw, and soybean and maize yields were analyzed, and chemical P fractionation was performed at soil depths of 0–0.05, 0.05–0.1, and 0.1–0.2 m. Results The intercropping systems affected the labile and moderately labile P pools at a depth of 0–0.05 m. On average, the intercropping systems decreased soil inorganic P (Pi) extracted by anion exchange resin (PiAER) by 26% compared with monocropped maize. In addition, the intercropping systems increased moderately labile organic P (Po) while simultaneously reducing the Pi pool. Intercropping palisade grass and/or sunn hemp with maize increased the depletion of labile and moderately labile Pi pools due to higher plant P demand driven by plant biomass. On the other hand, higher root exudates and root detritus enhanced the accumulation of moderately labile organic P under highly weathered soils. Intercropping palisade grass enhanced the yield of the main crop, i.e., soybean, but limited maize yield, presumably by creating an insufficient N supply for maize.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The main sources of phosphorus (P) for fertilizer production are finite and non-renewable (Ashley et al. 2011), which is a growing concern for crop and food security (Chowdhury et al. 2017). In weathered tropical soils, P is the most limiting nutrient for crop production and cropping system sustainability (Pavinato et al. 2020). Forest conversion to cropland mineralizes organic P (Po) species, which are readsorbed in non-labile forms, reducing P cycling in the system (Rigon et al. 2024). Hence, improving P use in cropping systems in tropical regions is essential.
Soil P cycling varies according to the crop species and its straw production (Tiecher et al. 2015). One strategy for enhancing soil P use efficiency (PUE) is to add cover crops in rotation under no-tillage (NT) (Rodrigues et al. 2021; Soltangheisi et al. 2018, 2019). Cover crop species vary in their ability to acquire less labile soil P pools (Hallama et al. 2019; Tiecher et al. 2012). The release of P from crop straw under NT management depends on the forms of P accumulated in plant tissues (de Oliveira et al. 2017) and on straw quality, such as nitrogen (N) content, carbon: nitrogen (C: N) ratio, lignin: N ratio, and cellulose and hemicellulose contents (Rigon et al. 2022). In general, legumes promote the efficient utilization of P from the soil/fertilizer in acidic soils (Hedley et al. 1995), whereas grasses can explore deeper soil layers and recycle more P in the whole system (Almeida and Rosolem 2016). As a result, some cereal/legume intercropping systems may substantially alter soil P availability over time (Tang et al. 2021; Teles et al. 2017; Tiecher et al. 2015).
The positive effects of intercropping on resource use efficiency in agroecosystems (Tang et al. 2021) are driven by two main processes: (i) resource partitioning between crops, which reduces crop competition (Fridley 2001), and (ii) positive interactions that alter the environment and enhance resource availability (Callaway 2007). These processes vary depending on differences in rooting pattern or depth interactions between species or differences in P demand during plant growth (Hinsinger et al. 2011; Yu et al. 2015).
Soil organic C cycling controls biological P availability (Rheinheimer et al. 2019). Because soil microorganisms are dependent on crop inputs, soil management practices and C inputs might enhance biological activity and improve soil P and Po availability (Cherubin et al. 2016; Frasier et al., 2022; Rigon et al. 2022). There is conflicting evidence on the relationships of Po with microbial biomass and soil organic matter (Cherubin 2016). Changes in crop P uptake and P return to soil affect the diversity of P species along the soil profile (Ferrarini et al. 2021). Consequently, soil P dynamics are expected to vary among crop rotation systems comprising different species alone and/or intercropped under long-term NT. In this study, we hypothesized that (i) soil P lability is influenced by intercropping systems and (ii) intercropping maize with palisade grass or sunn hemp in the offseason increases the soil Po pool. To test these hypotheses, we assessed differences in soil P dynamics between offseason intercropping systems under long-term NT.
2 Materials and methods
2.1 Site Characteristics
The experiment was initiated in 2006 at the Lageado experimental farm in Botucatu, São Paulo State, Brazil (48º 23’ W, 22º 51’ S, 765 m above sea level). The soil is a clayey Rhodic Hapludox (Soil Survey Staff 2014). The climate is highland tropical with dry winters and wet rainy summers (Fig. 1). At the time of the establishment of the experiment in 2006, the chemical properties of the soil in the 0–0.2 m layer as analyzed according to Raij et al. (2001) were as follows: soil organic matter = 18 g dm− 3; pH = 4.2 (1:2.5 soil:0.01 mol L− 1 CaCl2 suspension); P resin = 4 mg dm− 3; exchangeable K = 0.8 mmolc dm− 3, Ca = 12 mmolc dm− 3, Mg = 6 mmolc dm− 3; total acidity at pH 7.0 (H + Al) = 54 mmolc dm− 3; and base saturation = 24%.

Average monthly maximum (red line, ˚C), minimum (blue line, ˚C), and medium temperatures (black line, ˚C) and monthly rainfall (blue bars; mm) at the weather station located at Botucatu, Brazil
2.2 Experimental Design
The experiment was arranged as a randomized complete block design with four replications and four different cropping systems cultivated from 2007 to 2017. In any given year, the summer crop was the same in all treatments, but the crop choices in the offseason/winter season differed, as shown in Table 1. The summer grain crop choices were chosen based on the major crops cultivated in the Cerrado region.
In November 2018, soybean was sown in the total area. In March 2019, the cropping systems, i.e., the treatments, were updated to enhance the intensity of agricultural management and match the current scenario of cropping systems in Brazil. From 2018 to 2022, the offseason crops in the crop systems were as follows: (I) monocrop maize; (II) maize + palisade grass (U. brizantha); (III) maize + sunn hemp (C. spectabilis); and (IV) maize + palisade grass + sunn hemp. Soybean was cropped as the main crop in the summer during this period. A graphical scheme of the treatments is shown in Fig. 2.

Schematic representation of the study timeline showing the cropping systems
All cropping systems were managed under NT. Each plot had an area of 54 m2 (5.4 m x 10 m). One row on each side of the plot and one meter at both edges were considered buffer zones and were not sampled.
Over the first 11 years of the field experiment, the soil was amended with calcium and magnesium silicate (SiO2 = 220 g kg− 1) to increase the base saturation (BS) to 70%. The amendments were applied mechanically to the soil surface in October 2006 (before sowing the first crop), 2011, 2015, and 2018 (before sowing the summer crop) based on the results of BS determination (van Raij et al. 1997). Adequate rates of NPK fertilizers were applied to the summer crops according to the results of soil chemical analyses and crop recommendations.
2.3 Planting and Harvesting
At the time of installation of the experiment in 2018, the chemical properties of the soil in the 0–0.2 m layer as analyzed according to Raij et al. (2001) were as follows: soil organic matter = 27 g dm− 3; pH = 4.5 (1:2.5 soil:0.01 mol L− 1 CaCl2 suspension); resin P = 20 mg dm− 3; exchangeable K = 3.30 mmolc dm− 3, Ca = 31 mmolc dm− 3, Mg = 21 mmolc dm− 3; total acidity to pH 7.0 (H + Al) = 42 mmolc dm− 3; base saturation = 57%, CEC = 98 mmol dm− 3.
The experiment began in November 2018 with the sowing of soybean (CD 2728 IPRO) in the total area after the application of calcium and magnesium silicate. The soybean was harvested in March 2019, and soybean cultivar TMG 7062 IPRO was planted on 12 November 2019 and 26 October 2020 and harvested on 15 March 2020 and 25 February 2021. Both soybean crops were fertilized at sowing with 20 kg ha− 1 N, 50 kg ha− 1 P2O5 and 150 kg ha− 1 K2O. After the soybean harvest, maize hybrid P3707 VYW was sown on 17 March 2019 and 23 March 2020 and harvested on 8 May 2019 and 6 May 2020. For both maize crops, 22 kg ha− 1 N, 78 kg ha− 1 P2O5 and 45 kg ha− 1 K2O were supplied in the maize sowing furrow. When the maize plants had six fully expanded leaves, N fertilization was performed at 90 kg ha− 1 with ammonium sulfate. In system I, maize was not intercropped, and the area was fallow during the offseason. In system II, forage was sown simultaneously with the maize grain harvest at 8 kg of seeds ha− 1; the seeds were mixed together with the fertilizer a few hours before sowing. To sow the green fertilizer Crotalaria spectabilis in system III, the third box of the seeder (additional box of seeds) was used, and the seeds were sown at 17 kg ha− 1 simultaneously with maize. In system IV, i.e., the maize + palisade grass + sunn hemp triple intercropping system, sowing was performed under the same conditions as for maize alone. Palisade grass was mixed with the fertilizer and sown at 8 kg ha− 1, and sunn hemp was sown at 17 kg ha− 1 using the additional seed box.
2.4 Crop Analyses
To evaluate the contributions of the crop straw from the cover crops intercropped with maize, samples of the crop straw were collected at the time of desiccation before soybean sowing, i.e. 25 days before soybean sowing. Three subsamples were taken randomly from each subplot using a wooden square (0.25 × 0.25 m). The collected material was dried in a forced-air circulation oven at 65 °C until reaching a constant weight and was used to analyze dry matter and cumulative P content (Malavolta et al. 1997).
During the flowering periods of soybean and maize, leaf P levels were determined following the methodology proposed by Malavolta et al. (1997). For soybean, 30 samples of the third trefoil completely expanded from the apex were collected from each plot in the R2 stage. For maize, 30 samples of the leaf below and opposite the main ear were collected at the beginning of flowering.
Maize and soybean were harvested mechanically with a plot combine (NM Elite, Wintersteiger Seed Mech), and the crop yields were adjusted to 13% moisture. The P levels in the harvested maize and soybean grains (2019/2020 and 2020/2021 harvests) were analyzed according to the methodology of Malavolta et al. (1997). Soybean and maize grain P outputs were calculated by averaging the grain yields from both seasons (2019/2020, and 2020/2021) and multiplying the average grain yield by the P content.
2.5 Soil Analysis
Three soil subsamples were collected in October 2021 from each experimental unit at depths of 0–0.05, 0.05–0.1, and 0.1–0.2 m, air dried, and sieved (2 mm). Chemical P fractionation was performed according to Hedley et al. (1982) as modified by Rheinheimer et al. (2000), i.e., the original sonication step was replaced with extraction with 0.5 M NaOH. The soil P fractions were grouped into pools according to their potential lability (Cross and Schlesinger 1995): labile P (PiRTA + PiNaHCO3 + PoNaHCO3), moderately labile P (PiHID0.1 + PoHID0.1 + PiHCl), and non-labile P (PiHID0.5 + PoHID0.5 + P residual), and the data for the organic and inorganic pools were plotted in graphs.
2.6 Statistical Analysis
All statistical analyses were performed using SAS (Inc. 2009). All data were tested for normality with the Shapiro–Wilk test in the UNIVARIATE procedure. Data were analyzed using the MIXED procedure (mixed linear model), while the Satterthwaite approximation was used to determine denominator degrees of freedom to test fixed effects. The cropping system was considered a fixed effect. Blocks were treated as a random effect. The least significant difference (LSD) test was used to separate means through the LSMEANS procedure at p ≤ 0.1.
3 Results
3.1 Crop Straw Inputs from Intercropping
The cumulative crop straw on the soil surface and the straw P content differed significantly (p < 0.1) depending on the crop species intercropped with maize (Table 2). The average crop straw on the soil surface was 10 Mg ha− 1. Surprisingly, intercropping sunn hemp with maize did not significantly affect the crop straw input on the soil surface, whereas intercropping palisade grass with maize increased the crop straw input by an average of 144% compared with maize alone. Intercropping both palisade grass and sunn hemp with maize increased the crop straw input by 109% (p < 0.1) compared with intercropping only palisade grass with maize.
*Means followed by different letters differed by the t test (p ≤ 0.1).
†Pg: palisade grass; ‡Sh: sunn hemp.
On average, the crop straw P content was approximately 75% higher (p < 0.1) when maize was intercropped with palisade grass or sunn hemp than when maize was intercropped with both crops. Intercropping maize and palisade grass resulted in a cumulative P input of 23.82 kg ha− 1, 76% higher than the average of the other two intercropping systems.
3.2 Grain and Leaf P Contents
The cropping systems influenced grain and leaf P contents only in the maize crop grown in 2020 (Table 3). In this season, the leaf P content in maize intercropped with palisade grass was 31% higher than the average of the other intercropped systems. By contrast, the grain P content in maize intercropped with sunn hemp was 30% higher than the average of the other cropping systems.
3.3 Soybean and Maize Yields
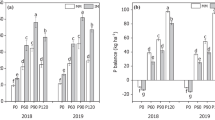
The average maize yield was ~ 4,800 kg ha− 1 in 2019 and 2020. As the main crop, the average soybean yield was 3.547 and 5.084 kg ha− 1 in the 2019/2020 and 2020/2021 seasons, respectively. The long-term cultivation system significantly affected (p < 0.1) maize yield in the 2019 and 2020 offseasons, soybean yield in the 2019/2020 season, and the cumulative yields of both crops (Fig. 3).

Yields of soybean as the main crop and maize as the offseason crop as a function of cropping system in the 2019 and 2020 seasons. *Means followed by different letters differ by the t test (p ≤ 0.1). The capital letters correspond to the cumulative data
In the 2019 and 2020 offseasons, maize yield was influenced by cropping system. In both maize harvests, the grain yield was highest when maize was intercropped with sunn hemp; in this cropping system, grain production was 19% and 53% higher than in the system with maize alone in 2019 and 2020, respectively. Compared with maize alone, intercropping maize with palisade grass increased the cumulative grain yield by 30%, and triple intercropping increased the cumulative grain yield by 22%. However, compared with the average of the other intercropped systems, triple intercropping reduced the cumulative grain yield by 10%.
For soybean grown in succession to maize, the different cropping systems influenced the grain yield in the 2019/2020 harvest and the cumulative grain yield. In the 2019/2020 season, soybean grain yield was highest after maize intercropped with palisade grass and maize intercropped with sunn hemp; in these cropping systems, soybean grain yield was 13% higher than in the system with maize alone and 15% higher than in the triple intercropping system. By contrast, the soybean grain yield in the 2020/2021 harvest did not differ between cropping systems. Compared with the system with maize alone, the cumulative soybean yield was 10% higher after maize intercropped with palisade grass.
3.4 Soil P Pools
The cropping systems affected most of the soil P fractions in the labile and moderately labile pools at a soil depth of 0–0.05 m (Supplementary table). In the deeper soil layers, the cropping systems only affected PoNaHCO3 at 0.5–0.1 m and PiAER and PiNaOH0.1 at 0.1–0.2 m. On average, intercropping maize with palisade grass and/or sunn hemp significantly (p < 0.1) decreased soil PiAER by 26% compared with cropping maize alone. Interestingly, PiAER (p < 0.1) and PiNaHCO3 (p = 0.21) decreased by 37% when maize was intercropped with sunn hemp. Among the moderately labile fractions, PiNaOH0.1 decreased (p < 0.1) by 25%, on average, at a soil depth of 0–0.05 m when maize was intercropped, but opposite trends were observed in deeper layers. Compared with triple intercropping, cropping maize alone decreased PiNaOH0.1 by 35% and 25% at soil depths of 0.05–0.1 and 0.1–0.2 m, respectively, but only the difference in the surface layer was significant. At a soil depth of 0–0.05 m, triple intercropping increased (p < 0.1) ) PoNaOH0.1 by 22% and 16% compared with intercropping maize with palisade grass or sunn hemp, respectively, whereas PoNaOH0.1 did not differ between maize intercropped with palisade grass or sunn hemp. At this depth, PoNaOH0.1 was significantly higher when maize was intercropped with palisade grass or sunn hemp than when maize cropped alone.
3.5 Soil P Lability
The total soil P content did not differ between the cropping systems and averaged 936, 806, and 765 mg kg− 1 in the 0–0.05, 0.05–0.1, and 0.1–0.2 m soil layers, respectively. In the surface soil layer, only 11.3% and 22.6% of the soil P stocks were allocated in the labile and moderately labile P pools, respectively (Fig. 4). As expected, the cropping systems impacted (p < 0.1) soil P only in the upper soil layer (0–0.05 m). Regardless of the cropping system, intercropping palisade grass or sunn hemp decreased (p < 0.1) soil labile and moderately labile P at a soil depth of 0–0.05 m (Fig. 4A), driven mostly by decreases in the inorganic P fractions (Fig. 4B). A similar trend was observed in the inorganic non-labile pool (Fig. 4D) but was not significant (p = 0.32). Overall, the effects of the cropping systems differed depending on P pool lability: intercropping increased (p < 0.1) moderately labile Po but slightly reduced (p = 0.12) moderately labile Pi.

Soil P pools by (A) lability, (B) labile organic and inorganic P, (C), moderately labile organic and inorganic P, (D) and non-labile organic and inorganic P at a soil depth of 0–0.05 m as a function of cropping system. *Means followed by different letters differ by the t test (p ≤ 0.1). ns: not significant
The contribution of soil labile P to P stocks was smaller in deeper soil layers but was not affected by cropping system. Intercropping maize with palisade grass reduced (< 0.1) the labile Po fraction by 58% at 0.05–0.1 m compared with intercropping maize with sunn hemp (Fig. 5) and moderately labile Pi by 30% at 0.1–0.2 m (Fig. 6) compared with triple intercropping.

Soil P pools by (A) lability, (B) labile organic and inorganic P, (C) moderately labile organic and inorganic P, and (D) non-labile organic and inorganic P at a soil depth of 0.05–0.1 m as a function of cropping system. *Means followed by different letters differ by the t test (p ≤ 0.1). ns: not significant

Soil P pools by (A) lability, (B) labile organic and inorganic P, (C) moderately labile organic and inorganic P, and (D) non-labile organic and inorganic P at a soil depth of 0.1–0.2 m as a function of cropping system. *Means followed by different letters differed by the t test (p ≤ 0.1). ns: not significant
4 Discussion
4.1 Crop Straw Inputs from Intercropping
Intercropping systems are widely used to increase biomass production in the offseason in weathered tropical soil, especially in regions with dry conditions (Crusciol et al. 2015). Under NT, crop straw on the surface of tropical soils decomposes quickly, which increases the importance of enhancing crop biomass production (Ferrari Neto et al. 2020; Lal 2002; Rigon and Calonego 2020). Palisade grass has high biomass production capacity (Almeida et al. 2020; Capstaff and Miller 2018), and in this study, intercropping with palisade grass greatly increased crop straw inputs compared with intercropping with sunn hemp. However, the legume sunn hemp produces high-quality crop straw (Rigon et al. 2021a), and intercropping sunn hemp with maize may increase the supply of N to intensive production systems through biological fixation by the legume itself or the release of nitrogenous compounds from the decomposition of the legume’s nodules and roots. However, legumes have low C: N ratios, which leads to rapid decomposition of their plant straw and less persistent soil cover (Rigon et al. 2021b).
The technical considerations for designing an intercropping system include competition for light, water and nutrients between plants, which can harm not only the productivity of the main crop but also the development of the cover crop (MacLaren et al. 2023). Because maize is a tall C4 crop with rapid initial growth, it is highly competitive with C3 species such as sunn hemp. In the intercropping systems evaluated in this work, straw accumulation was lowest when maize was intercropped with sunn hemp and similar to that in the system with maize alone. The lower accumulation of plant straw in the sunn hemp intercropping system may reflect not only competition with maize for natural resources but also the sensitivity of sunn hemp to photoperiod. For example, late sowing in the autumn–winter period advances sunn hemp flowering by a few days compared with summer sowing (Dzvene et al. 2022). Interestingly, a similar study found that intercropping maize with sunn hemp or palisade grass increased straw production without reducing maize grain yield compared with monocropping maize (Souza et al. 2022). In the present study, biomass production was highest in the triple intercropping system, i.e., maize intercropped with both palisade grass and sunn hemp. Intercropping two or more species, particularly with legume species, may improve soil structure and fertility (Garland et al. 2017) because the biological fixation of atmospheric N increases the N supply and consequently favors biomass production by grasses.
Although P content was high in both sunn hemp and palisade grass tissues, palisade grass had the highest cumulative P contribution due to its higher biomass production. Palisade grass has high P uptake efficiency, particularly in P-restricted soils, and improves plant-available P in long-term cultivation systems (Almeida et al. 2019, 2020). Our results show that plant P acquisition strategies and uptake capacities differ between cover crop species. By clearly highlighting the differences in P inputs between crop species intercropped with maize, this study provides a better understanding of the potential nutrient-cycling capacity of intercropping systems in tropical soils.
4.2 Grain and Leaf P Contents
In the 2019 season, maize leaf P content was within the range proposed for maize in all cropping systems and did not differ between cropping systems (Raij et al. 2001). By contrast, the cropping systems affected maize leaf P content in the 2020 season, but the values were below the ideal range for maize. The differences between seasons are likely attributable to differences in climatic conditions.
In 2020, maize leaf P content was highest when maize was intercropped with palisade grass and differed significantly between this cropping system and the other systems. This result is consistent with the high P accumulation and biomass production in this intercropping system. P is a component of protein modifications, and a portion of the P in soluble compounds may become available to successive crops, impacting leaf P levels (Hedley et al. 1995). The P-cycling efficiency of palisade grass is further confirmed by the marked decrease in P export in maize grains in the system with maize intercropped with palisade grass. We assume that P export was lowest in this system because leaf P content was highest. However, increases in P content in maize grain are not always accompanied by corresponding increases in P content in straw (Xia et al. 2019).
4.3 Soybean and Maize Yields
In southern Brazil, dry conditions are typical during the fall/winter offseason (Cunningham 2020). In the 2019 season, water deficits may have intensified the competition between maize and palisade grass, as these crops are physiologically similar (Friedman 2024). Split application of N can increase the responses of both maize and forage grasses to N fertilization (Crusciol et al. 2020). Grass–grass rotations without N fertilization can promote significant N immobilization due to competition between plants and microorganisms (Kuzyakov and Xu 2013), resulting in an insufficient N supply for maize intercropped with palisade grass. These effects may explain the increased yield of maize intercropped with sunn hemp; the introduction of this legume may have guaranteed a greater supply of N through biological fixation, ensuring greater N acquisition by the grass (i.e., maize).
The cumulative soybean yield was higher after maize intercropped with palisade grass than after monocropped maize, although soybean yield did not differ between the intercropping systems. Higher biomass production and P accumulation in straw were expected in the intercropping systems because intercropping mutually improves crop P nutrition for all crops (Duchene et al. 2017). According to Yang et al. (2022), the crops in intercropping systems differ in their preferences for soil P pool composition, P bioavailability, and P uptake. These differences lead to resource partitioning of soil P (Turner and Lambert 2008). The contrasting effects of the cropping systems on the yields of the offseason and main crops highlight the importance of a holistic approach to cropping systems that does not focus solely on a specific crop.
4.4 Soil P Pools
Long-term cropping with annual P fertilization (± 50 kg P ha− 1) under NT in the weathered tropical soils of the Cerrado intensifies P stratification in topsoil, offsets the high P immobilization potential (Withers et al. 2018), and ensures adequate soil available P (PAER averaged at a soil depth of 0–0.1 m) for main crops (Raij, B. et al. 2001; Souza et al. 2004). The soil P pools at the soil surface (0–0.05 m soil depth) in this study confirmed the high soil P immobilization in the upper layers and the lack of downward movement (Soltangheisi et al. 2018). Compared with maize alone, the intercropping systems depleted PAER at a soil depth of 0–0.05 m, which is attributable to higher crop uptake of P from fertilizer and temporary accumulation in crop tissues, as confirmed by the crop straw inputs (Table 2). Increasing P uptake reduces the availability of soil P forms (Rigon et al. 2022; Rodrigues et al. 2021). Hence, higher soil P availability is associated with lower biomass production, as observed in the straw input in the monocropped maize system. The intercropping systems also depleted PiNaHCO3, indicating the transformation of relatively labile Po into Pi (Almeida and Rosolem 2016; Rodrigues et al. 2021). These results suggest that the intercropping systems increased Pi immobilization to aid straw decomposition, as confirmed by the low labile Po (Rodrigues et al. 2016). Crop straw mineralization under NT occurs via transient microbial immobilization, which prevents rapid Pi adsorption (Martinazzo et al. 2007), an important process in Oxisols.
Confirming our hypothesis, Po NaOH 0.1M was highest and Pi NaOH 0.1M was lowest when maize was intercropped with palisade grass or sunn hemp. Maize leaf P content was also highest in these two systems, demonstrating the influence of P availability on P uptake. The accumulation of moderately labile Po could be attributed to microbial immobilization and the accumulation of root residues in Al and Fe oxides present in highly weathered soils. Greater P immobilization in microbial biomass is expected in weathered soils (Waithaisong et al. 2022). Organic acids exuded from the roots of companion crops can promote the solubility of poorly labile P forms (Pavinato et al. 2008). Intercropping perennial grasses improves the Po pool by increasing root development and the production of organic acid exudates (Almeida et al. 2020; Almeida and Rosolem 2016). Organic acids compete with P for adsorption sites on soil surfaces, reducing P adsorption (Sims et al. 2005). The addition of legumes to rotation systems also affects the soil Po pool (Rigon et al. 2022). Consistent with the effects of intercropping maize with sunn hemp in this study, cereal–legume intercropping systems (Yang et al. 2022) promote P uptake by the intercropped P-inefficient crop (Li et al. 2004; Yang et al. 2022). Our results confirm that crop species impact soil P cycling, P use efficiency, and organic P pools in cropping systems (Hallama et al. 2019; Rigon et al. 2022, 2024; Soltangheisi et al. 2018; Teles et al. 2017).
A low contribution of Ca-precipitated P (PHCL) to total soil P was expected because this P pool is more important in temperate soils (Jin et al. 2021). Presidue, which has been reported to function as a P input sink (Deiss et al. 2016; Rigon et al. 2022; Rodrigues et al. 2021; Soltangheisi et al. 2018), accounted for almost half of the soil P stocks but was not affected by the cropping systems. Similarly, the cropping systems did not affect recalcitrant Pi and PoNaOH0.5 pools.
4.5 Soil P Lability
Our results show that long-term cropping systems can effectively change soil P lability, mainly at a soil depth of 0–0.05 m, thus partially confirming our hypothesis. We anticipated that after 14 years, the cropping systems under NT would substantially increase the labile P pool at the soil surface (~ 11%). This expectation was based on the ability of crop straw on the soil surface to reduce P fixation and stimulate P cycling and crop uptake. Presumably, these effects are restricted to the soil surface (Rigon et al. 2022, 2024; Soltangheisi et al. 2018; Tiecher et al. 2017) and are smaller or absent in deeper layers (Rodrigues et al. 2021; Soltangheisi et al. 2018; Teles et al. 2017). Tropical weathered soils, which are dominated by kaolinite, gibbsite, and hematite (Rigon et al. 2022), have high P adsorption (Schaefer et al. 2008), and most of the P added by fertilization accumulates in moderately labile and non-labile forms (Rodrigues et al. 2016; Withers et al. 2018), as observed in our study. It is possible to infer that the effects of P fertilization under NT and crop straw from intercropping systems are limited to the upper soil layers (Teles et al. 2017).
The higher crop straw inputs from the intercropping systems compared to maize alone explain the temporary P accumulation in tissues and the reduced labile Pi (Rodrigues et al. 2021), which had cascading effects on the labile P pool. In weathered tropical soils, soil P lability depends more on biologically mediated Po turnover processes than on Pi adsorption (Oberson et al. 2001). The organic pool is second only to mineralization as an important source of P for crops in these soils (Vincent et al. 2010), and growing crops in the fall–winter season under NT enhances the importance of the organic pool for P cycling (Rigon et al. 2022; Tiecher et al. 2012). Urochloa grasses, particularly palisade grass, may increase moderately labile Po (Almeida et al. 2019, 2020; Almeida and Rosolem 2016). Under low-P conditions, the roots of Urochloa species exude organic acids, which may affect agronomic PUE. These processes help explain the effects of the intercropping systems, particularly the effects of intercropping maize with palisade grass, on the moderately labile Po fraction. In addition, our results confirm that intensive cropping systems under NT stimulate microbial Po cycling to enhance the biological P pool (Cherubin et al. 2016). The Po pool plays an essential role in soil P bioavailability because it is less susceptible to soil adsorption in tropical soils (Damodar Reddy et al. 2000; Pavinato et al. 2017; Shears and Turner 2007). Although organic P has received little attention, it is essential for P availability to plants in the rhizosphere (Zhang et al. 2023).
Clay minerals strongly adsorb non-labile P fractions with low reversibility; consequently, the cropping systems did not impact this soil P pool. The contribution of the non-labile P pool to the total soil P in tropical weathered soils is greater than those of the labile and moderately labile P pools (Pavinato et al. 2009; Rigon et al. 2022; Rodrigues et al. 2016, 2021; Soltangheisi et al. 2020; Teles et al. 2017). The contribution of this inaccessible pool to soil P in the surface soil layers despite long-term NT demonstrates the ability of soil management strategies to increase the efficiency and sustainability of cropping systems in tropical soils.
5 Conclusion
The differences in crop straw inputs between crop species intercropped with maize in the offseason under long-term NT affected soil P pools and P lability, but these effects were limited to the soil surface. By contrast, the high soil P immobilization capacity of weathered tropical soils did not permit changes in soil P stocks and the non-labile P pool. Intercropping palisade grass and/or sunn hemp with maize increased the depletion of labile and moderately labile Pi pools due to higher plant P demand driven by plant biomass. On the other hand, higher root exudates and root detritus enhanced the accumulation of moderately labile organic P under highly weathered soils of Brazil where the concentrations of Al and Fe oxides are high. The intercropping treatments had opposing effects on crop yields: palisade grass enhanced the yield of the main crop, soybean, but limited maize yield, suggesting that intercropped maize requires a higher N supply.
References
Almeida DS, Rosolem CA (2016) Ruzigrass grown in rotation with soybean increases soil labile phosphorus. Agron J 108:2444–2452. https://doi.org/10.2134/agronj2015.0478
Almeida DS, Menezes-blackburn D, Zhang H, Haygarth PM, Rosolem CA (2019) Geoderma Phosphorus availability and dynamics in soil affected by long-term ruzigrass cover crop. Geoderma 337:434–443. https://doi.org/10.1016/j.geoderma.2018.09.056
Almeida DS, Delai LB, Sawaya ACHF, Rosolem CA (2020) Exudation of organic acid anions by tropical grasses in response to low phosphorus availability. Sci Rep 10:1–8. https://doi.org/10.1038/s41598-020-73398-1
Ashley K, Cordell D, Mavinic D (2011) A brief history of phosphorus: from the philosopher’s stone to nutrient recovery and reuse. Chemosphere 84:737–746
Callaway RM (2007) Direct mechanisms for Facilitation. Positive interactions and interdependence in Plant communities. Springer Netherlands, Dordrecht, pp 15–116
Capstaff NM, Miller AJ (2018) Improving the yield and nutritional quality of forage crops. Front Plant Sci 9:1–18. https://doi.org/10.3389/fpls.2018.00535
Cherubin MR, Franco ALC, Cerri CEP, Karlen DL, Pavinato PS, Rodrigues M, Davies CA, Cerri CC (2016) Phosphorus pools responses to land-use change for sugarcane expansion in weathered Brazilian soils. Geoderma 265:27–38. https://doi.org/10.1016/j.geoderma.2015.11.017
Chowdhury RB, Moore GA, Weatherley AJ, Arora M (2017) Key sustainability challenges for the global phosphorus resource, their implications for global food security, and options for mitigation. J Clean Prod 140:945–963
Cross AF, Schlesinger WH (1995) A literature review and evaluation of the. Hedley fractionation: applications to the biogeochemical cycle of soil phosphorus in natural ecosystems. Geoderma 64:197–214. https://doi.org/10.1016/0016-7061(94)00023-4
Crusciol CAC, Nascente AS, Borghi E, Soratto RP, Martins PO (2015) Improving soil fertility and crop yield in a tropical region with palisadegrass cover crops. Agron J 107:2271–2280. https://doi.org/10.2134/agronj14.0603
Crusciol CAC, Mateus GP, Momesso L, Pariz CM, Castilhos AM, Calonego JC, Borghi E, Costa C, Franzluebbers AJ, Cantarella H (2020) Nitrogen-Fertilized systems of Maize Intercropped with Tropical grasses for enhanced yields and estimated Land Use and Meat Production. Front Sustainable Food Syst 4:1–13. https://doi.org/10.3389/fsufs.2020.544853
Cunningham C (2020) Characterization of dry spells in Southeastern Brazil during the monsoon season. Int J Climatol 40:4609–4621. https://doi.org/10.1002/joc.6478
Damodar Reddy D, Subba Rao A, Rupa TR (2000) Effects of continuous use of cattle manure and fertilizer phosphorus on crop yields and soil organic phosphorus in a Vertisol. Bioresour Technol 75:113–118. https://doi.org/10.1016/S0960-8524(00)00050-X
de Oliveira RA, Comin JJ, Tiecher T, Piccin R, Somavilla LM, Loss A, Lourenzi CR, Kürtz C, Brunetto G (2017) Release of phosphorus forms from cover crop residues in agroecological no-till onion production. Revista Brasileira De Ciencia do Solo 41:1–16. https://doi.org/10.1590/18069657rbcs20160272
Deiss L, de Moraes A, Dieckow J, Franzluebbers AJ, Gatiboni LC, Sassaki G, lanzi, Carvalho PCF (2016) Soil phosphorus compounds in integrated crop-livestock systems of subtropical Brazil. Geoderma 274:88–96. https://doi.org/10.1016/j.geoderma.2016.03.028
Duchene O, Vian J-F, Celette F (2017) Intercropping with legume for agroecological cropping systems: complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric Ecosyst Environ 240:148–161. https://doi.org/10.1016/j.agee.2017.02.019
Dzvene AR, Tesfuhuney W, Walker S, Ceronio G (2022) Effects of intercropping sunn hemp into maize at different times and densities on productivity under rainwater harvesting technique. Front Sustain Food Syst 6:1009443. https://doi.org/10.3389/fsufs.2022.1009443
Ferrari Neto J, Franzluebbers AJ, Crusciol CAC, Rigon JPG, Calonego JC, Rosolem CA, do Nascimento CAC, Ribeiro LC (2020) Soil carbon and nitrogen fractions and physical attributes affected by soil acidity amendments under no-till on Oxisol in Brazil. Geoderma Reg 24:e00347. https://doi.org/10.1016/j.geodrs.2020.e00347
Ferrarini A, Martani E, Fornasier F, Amaducci S (2021) High C input by perennial energy crops boosts belowground functioning and increases soil organic P content. Agric Ecosyst Environ 308:107247. https://doi.org/10.1016/j.agee.2020.107247
Fridley JD (2001) The influence of species diversity on ecosystem productivity: how, where, and why? Oikos 3
Friedman SP (2024) Relationships between combined and individual field crops’ biomass and planting density. Field Crops Res 305:109188. https://doi.org/10.1016/j.fcr.2023.109188
Garland G, Bünemann EK, Oberson A, Frossard E, Six J (2017) Plant-mediated rhizospheric interactions in maize-pigeon pea intercropping enhance soil aggregation and organic phosphorus storage. Plant Soil 415:37–55. https://doi.org/10.1007/s11104-016-3145-1
Hallama M, Pekrun C, Lambers H, Kandeler E (2019) Hidden miners– the roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 434:7–45. https://doi.org/10.1007/s11104-018-3810-7
Hedley MJ, Stewart JWB, Chauhan BS (1982) Changes in Inorganic and Organic Soil Phosphorus fractions Induced by Cultivation practices and by Laboratory Incubations. Soil Sci Soc Am J 46:970–976. https://doi.org/10.2136/sssaj1982.03615995004600050017x
Hedley JM, Mortvedt JJ, Bolan NS, Syers JK (1995) Phosphorus fertility management in agroecosystems. In: Tiessen H (ed) Phosphorus in the global environment. Transfers, cycles and management. Scope 54. Wiley, New York, pp 59–92
Hinsinger P, Betencourt E, Bernard L, Brauman A, Plassard C, Shen J, Tang X, Zhang F (2011) P for two, sharing a scarce resource: soil phosphorus acquisition in the rhizosphere of intercropped species. Plant Physiol 156:1078–1086. https://doi.org/10.1104/pp.111.175331
Inc. SI (2009) The SAS System for Windows
Jin X, Hu C, Khan A, Zhang S, Yang X, Jia L, Sun R (2021) Comparison of fractionation methods for soil phosphorus with soils subjected to various long-term fertilization regimes on a calcareous soil. PeerJ Inorg Chem 3:e3. https://doi.org/10.7717/peerj-ichem.3
Kuzyakov Y, Xu X (2013) Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytol 198:656–669. https://doi.org/10.1111/nph.12235
Lal R (2002) The potential of soils of the tropics to sequester carbon and mitigate the greenhouse effect. Adv Agron 76:1–30. https://doi.org/10.1016/s0065-2113(02)76002-1
Li SM, Li L, Zhang FS, Tang C (2004) Acid phosphatase role in chickpea/maize intercropping. Ann Botany 94:297–303. https://doi.org/10.1093/aob/mch140
MacLaren C, Waswa W, Aliyu KT, Claessens L, Mead A, Schöb C, Vanlauwe B, Storkey J (2023) Predicting intercrop competition, facilitation, and productivity from simple functional traits. Field Crops Res 297:108926
Malavolta E, Vitti GC, de Oliveira SA (1997) Avaliação do estado nutricional das plantas: principios e aplicações. Associação Brasileira para Pesquisa da Potassa e do Fosfato
Martinazzo R, Dos Santos DR, Gatiboni LC, Brunetto G, Kaminski J (2007) Microbial phosphorus in a soil under no-tillage as affected by soluble phosphorus addition. Revista Brasileira De Ciencia do Solo 31:563–570. https://doi.org/10.1590/s0100-06832007000300016
Oberson A, Friesen DK, Rao IM, Bühler S, Frossard E (2001) Phosphorus transformations in an Oxisol under contrasting land-use systems: the role of the soil microbial biomass. Plant Soil 237:197–210. https://doi.org/10.1023/A:1013301716913
Pavinato PS, Merlin A, Rosolem CA (2008) Organic compounds from plant extracts and their effect on soil phosphorus availability. Pesquisa Agropecuaria Brasileira 43:1379–1388. https://doi.org/10.1590/S0100-204X2008001000017
Pavinato PS, Merlin A, Rosolem CA (2009) Phosphorus fractions in Brazilian Cerrado soils as affected by tillage. Soil Tillage Res 105:149–155. https://doi.org/10.1016/j.still.2009.07.001
Pavinato PS, Rodrigues M, Soltangheisi A, Sartor LR, Withers PJA (2017) Effects of cover crops and phosphorus sources on maize yield, phosphorus uptake, and phosphorus use efficiency. Agron J 109:1039–1047. https://doi.org/10.2134/agronj2016.06.0323
Pavinato PS, Rocha GC, Cherubin MR, Harris I, Jones DL, Withers PJA (2020) Map of total phosphorus content in native soils of Brazil. Scientia Agricola 78:1–5. https://doi.org/10.1590/1678-992x-2020-0077
Raij BV, Andrade JC, Cantarella H, Quaggio JA (2001) Chemical analysis for evaluation of the fertility of tropical soils. Instituto Agronômico, Campinas
Rheinheimer DS, Anghinoni I, Kaminski J (2000) Depletion of inorganic phosphorus fractions by successive extraction with Resin in different soils and Management systems. R Bras Ci Solo 24:345–354
Rheinheimer Ddos, Fornari S, Bastos MR, Fernandes MC, Santanna G, Calegari MA, dos Santos Canalli A, Caner LB, Labanowski L, Tiecher J T (2019) Phosphorus distribution after three decades of different soil management and cover crops in subtropical region. Soil Tillage Res 192:33–41. https://doi.org/10.1016/j.still.2019.04.018
Rigon JPG, Calonego JC (2020) Soil carbon fluxes and balances of crop rotations under long-term no-till. Carbon Balance Manag 15:19. https://doi.org/10.1186/s13021-020-00154-3
Rigon JPG, Calonego JC, Capuani S, Franzluebbers AJ (2021a) Soil organic C affected by dry-season management of no‐till soybean crop rotations in the tropics. Plant Soil 462:577–590. https://doi.org/10.1007/s11104-021-04878-0
Rigon JPG, Calonego JC, Pivetta LA, Castoldi G, Raphael JPA, Rosolem CA (2021b) Using cover crops to offset greenhouse gas emissions from a tropical soil under no-till. Exp Agric 57. https://doi.org/10.1017/S0014479721000156
Rigon JPG, Crusciol CAC, Calonego J, Pavinato PS, Azevedo AC, Rosolem CA (2022) Intensive crop rotations and residueSoil & Tillage Research quality increase soil phosphorus lability under long-term no-till in tropical soils. Soil Tillage Res. https://doi.org/10.1016/j.still.2022.105446. 223:
Rigon JPG, Crusciol CAC, Calonego JC, Gatiboni LC, Pavinato PS, Colzato M, Capuani S, Rosolem CA (2024) Phosphorus speciation under long-term crop rotation management in a tropical soil. Soil Use Manag 40:e13006. https://doi.org/10.1111/sum.13006
Rodrigues M, Pavinato PS, Withers PJA, Teles APB, Herrera WFB (2016) Legacy phosphorus and no tillage agriculture in tropical oxisols of the Brazilian savanna. Sci Total Environ 542:1050–1061. https://doi.org/10.1016/j.scitotenv.2015.08.118
Rodrigues M, Withers PJA, Soltangheisi A, Vargas V, Holzschuh M, Pavinato PS (2021) Tillage systems and cover crops affecting soil phosphorus bioavailability in Brazilian Cerrado Oxisols. Soil Tillage Res 205:104770. https://doi.org/10.1016/j.still.2020.104770
Schaefer CEGR, Fabris JD, Ker JC (2008) Minerals in the clay fraction of Brazilian latosols (Oxisols): a review. Clay Miner 43:137–154. https://doi.org/10.1180/claymin.2008.043.1.11
Shears SB, Turner BL (2007) Nomenclature and terminology of inositol phosphates: clarification and a glossary of terms. Inositol Phosphates Linking Agriculture and the Environment. 1–6
Sims JT, Sharpley AN, Condron LM, Turner BL, Cade-Menun BJ (2005) Chemistry and Dynamics of Soil Organic Phosphorus. https://doi.org/10.2134/agronmonogr46.c4
Soil Survey Staff (2014) Keys to Soil Taxonomy, 12th edn. USDA-Natural Resources Conservation Service, Washington, DC
Soltangheisi A, Rodrigues M, Coelho MJAJA, Gasperini AMM, Sartor LRR, Pavinato PSS (2018) Changes in soil phosphorus lability promoted by phosphate sources and cover crops. Soil Tillage Res 179:20–28. https://doi.org/10.1016/j.still.2018.01.006
Soltangheisi A, de Moraes MT, Cherubin MR, Alvarez DO, de Souza LF, Bieluczyk W, Navroski D, Bettoni Teles AP, Pavinato PS, Martinelli LA, Tsai SM, de Camargo PB (2019) Forest conversion to pasture affects soil phosphorus dynamics and nutritional status in Brazilian Amazon. Soil Tillage Res 194:104330. https://doi.org/10.1016/j.still.2019.104330
Soltangheisi A, Teles APB, Sartor LR, Pavinato PS (2020) Cover cropping may alter legacy Phosphorus Dynamics under Long-Term Fertilizer Addition. Front Environ Sci 8:1–12. https://doi.org/10.3389/fenvs.2020.00013
Souza DMG de;, Lobato E, Souza DMG de;, Lobato E (2004) Calagem e adubação para culturas anuais e semiperenes. In: SOUSA DMG de;, Lobato E (eds) Cerrado: correção do solo e adubação, 2nd edn. Embrapa Informação Tecnológica, Brasília, DF, DF, pp 283–315
Souza SSD, Couto Júnior PA, Flôres JDA, Coelho AP, Lemos LB (2022) Do the intercropping with Crotalaria spectabilis and Urochloa ruziziensis reduce the maize agronomic performance? Pesqui Agropecu Trop 52:e70513. https://doi.org/10.1590/1983-40632022v5270513
Tang X, Zhang C, Yu Y, Shen J, van der Werf W, Zhang F (2021) Intercropping legumes and cereals increases phosphorus use efficiency; a meta-analysis. Plant Soil 460:89–104. https://doi.org/10.1007/s11104-020-04768-x
Teles APB, Rodrigues M, Bejarano Herrera WF, Soltangheisi A, Sartor LR, Withers PJA, Pavinato PS (2017) Do cover crops change the lability of phosphorus in a clayey subtropical soil under different phosphate fertilizers? Soil Use Manag 33:34–44. https://doi.org/10.1111/sum.12327
Tiecher T, dos Santos DR, Calegari A (2012) Soil organic phosphorus forms under different soil management systems and winter crops, in a long term experiment. Soil Tillage Res 124:57–67. https://doi.org/10.1016/j.still.2012.05.001
Tiecher TTL, Oliveira LB, De, Caner L, Brunetto G, Bortoluzzi EC, Rheinheimer DS, Casali CA, Zafar M, Tiecher TTL (2015) Cover crops affecting soil phosphorus dynamics in Brazilian highly weathered soils. In: Reuter J (ed) Cover crops: cultivation, management and benefits. Nova Science, New York, pp 23–52
Tiecher T, Calegari A, Caner L, dos Rheinheimer D S (2017) Soil fertility and nutrient budget after 23-years of different soil tillage systems and winter cover crops in a subtropical Oxisol. Geoderma 308:78–85. https://doi.org/10.1016/j.geoderma.2017.08.028
Turner J, Lambert MJ (2008) Nutrient cycling in age sequences of two Eucalyptus plantation species. For Ecol Manag 255:1701–1712. https://doi.org/10.1016/j.foreco.2007.11.038
van Raij B, Cantarella H, Quaggio JA, Furlani AMC (1997) Recomendações de adubação e calagem para o estado de São Paulo
Vincent AG, Turner BL, Tanner EVJ (2010) Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest. Eur J Soil Sci 61:48–57. https://doi.org/10.1111/j.1365-2389.2009.01200.x
Waithaisong K, Robin A, L’Huillery V, Abadie J, Sauvage F-X, Chemardin P, Mareschal L, Bouillet J-P, Laclau J-P, Gonçalves JLM, Plassard C (2022) Organic phosphorus immobilization in microbial biomass controls how N2-fixing trees affect phosphorus bioavailability in two tropical soils. Environ Adv 8:100247. https://doi.org/10.1016/j.envadv.2022.100247
Withers PJA, Rodrigues M, Soltangheisi A, De Carvalho TS, Guilherme LRG, Benites VDM, Gatiboni LC, De Sousa DMG, Nunes RDS, Rosolem CA, Andreote FD, Oliveira A, De, Coutinho ELM, Pavinato PS (2018) Transitions to sustainable management of phosphorus in Brazilian agriculture. Sci Rep 8:1–13. https://doi.org/10.1038/s41598-018-20887-z
Xia H, Wang L, Jiao N et al (2019) Luxury absorption of phosphorus exists in maize when intercropping with legumes or oilseed rape-covering different locations and years. Agronomy 9. https://doi.org/10.3390/agronomy9060314
Yang Z, Zhang Y, Wang Y, Zhang H, Zhu Q, Yan B, Fei J, Xiangmin R, Peng J, Luo G (2022) Intercropping regulation of soil phosphorus composition and microbially-driven dynamics facilitates maize phosphorus uptake and productivity improvement. Field Crops Res 287. https://doi.org/10.1016/j.fcr.2022.108666
Yu Y, Stomph TJ, Makowski D, van der Werf W (2015) Temporal niche differentiation increases the land equivalent ratio of annual intercrops: a meta-analysis. Field Crops Res 184:133–144. https://doi.org/10.1016/j.fcr.2015.09.010
Zhang K, Zheng D, Gu Y, Xu J, Wang M, Mu B, Wen S, Tang T, Rengel Z, Shen J (2023) Utilizing soil organic phosphorus for sustainable crop production: insights into the rhizosphere. Plant Soil. https://doi.org/10.1007/s11104-023-06136-x
Acknowledgement
This study was financed by the Coordination of Higher Level Personal Improvement (CAPES) Finance Code 001.The first author thanks the PROPe– Unesp edital 13/2022 for the scholarship granted. To the National Council for Scientific and Technological Development (CNPq), for the scholarship granted to the 2nd author. In addition, fifth, sixth, and seventh authors thanks the CNPq for an award for excellence in research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
We have no conflicts of interest to disclose.
Statements
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Rigon, J.P.G., Vilela, R.G., Teles, A.P.B. et al. Soil Phosphorus Dynamics under Long-Term No-Till with Offseason Intercropping Systems. J Soil Sci Plant Nutr (2024). https://doi.org/10.1007/s42729-024-01918-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42729-024-01918-3