
Abstract
Dunaliella salina a halotolerant microalga is well known for a high fatty acid and beta-carotene content, which makes it a potent source at a commercial level. The current study focuses on optimizing commercially known media using Box Behnken design to attain higher yields of biomass, beta-carotene, and lipids simultaneously. The optimal medium conditions as per response surface methodology were glucose, potassium nitrate, sodium chloride at a concentration of 13.23 g/L, 3.145 g/L and 35.6 g/L, respectively while maintaining the concentration of other nutrients unchanged. Maximum yield of biomass, beta-carotene and lipid productivity attained experimentally using the optimized media was 1.24 g/L, 6.07 mg/g and 20.7 mg/L/day, respectively than their original values i.e., 0.571 g/L of biomass, 4.18 mg/g of beta-carotene and 13.2 mg/L/day of lipid content. Biomass yield was increased by 2.17 folds, beta-carotene and lipid were increased by 1.45 folds and 1.56 folds, respectively.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Dunaliella salina synthesizes beta-carotene in chloroplast lipid globules (Rabbani et al. 1998). Carotenoids are known to possess antioxidant activity, thus quenching the free radicals and preventing cell damage. In such condition, the chloroplast begins to shrink and turn yellow-orange in colour from green (Mojaat et al. 2008). Two major beta-carotene isomers present in D. salina are 9-cis–beta carotene and all-trans-beta-carotene. Under stress conditions beta-carotene can contribute up to 10% of dry algal weight (García-González et al. 2005). Open raceway ponds are constructed to culture Dunaliella as chances of contamination are rare since extreme growth conditions do not favour the growth of other microbes (Del Campo et al. 2007; Lamers et al. 2012). Few Dunaliella raceway ponds are set up on industrial scale in the coastal deserts of Israel, Australia, Hawaii (Gary and Masao, 2001). However, there is a limitation to scaling up the mass culture whereas the closed tubular photo bioreactors maintain controlled environments to achieve maximum productivity (Wu et al. 2016; Hosseini and Shariati 2009).
In 2010, worldwide beta-carotene production reached to 233 million dollars with an annual growth rate of 3.6% and is expected to increase up to 309 million dollars by 2018 (Mata-Gómez et al. 2014). Beta-carotene can be produced from various sources including plants, yeast, bacteria, and algae. In case of beta-carotene production from algae, only Dunaliella salina is studied extensively compared to other marine algal species (Bonnefond et al. 2017). Dunaliella salina has the ability to accumulate huge amounts of beta carotene under stress conditions that mainly includes nutrient starvation, salt stress, and high irradiance (Chen et al. 2011; Cheirsilp et al. 2012; Srinivasan et al. 2015). These stress conditions disturb the physiological equilibrium resulting in the synthesis of free radicals and reactive oxygen species (ROS) that can lead to DNA mutations or react with chlorophyll in the chloroplast and lead to cell death (Sun et al. 2018). Response Surface Methodology (RSM) is a statistical tool used for media optimization in cases where multiple variables are varied to achieve an optimal output (Azma et al. 2011). RSM has been successfully used to enhance the productivity of lipids, proteins, lutein, zeaxanthin, and beta-carotene (Azma et al. 2011; Hallenbeck et al. 2015; Singh et al. 2016).
Conventional one factor approach requires setting up of large number of experiments which is an expensive and time-consuming process. RSM is a simple and precise method to optimize the concentration of limiting nutrients in a medium without affecting the cost of production (Kirrolia et al. 2014). However, very few studies using RSM to subsequently enhance growth, beta-carotene and lipid production in Dunaliella salina cultures has been done (Morowvat and Younes 2016). There is a need to explore the interactions between carbon, nitrogen and salinity source to comprehend the significant impact of culture medium.
Media composition plays an important function in microalgal growth and lipid production which subsequently affects the total biomass productivity. In order to find the suitable culture medium conditions to improve the biomass and beta-carotene production, several parameters should be considered. Organic carbon sources, nitrogen and phosphate have influence the production of carotenoids and lipids (Lamers et al. 2012; Mojaat et al. 2008) and they are critical parameters. Organic carbon sources, nitrogen starvation and an increased salinity have contributed in higher accumulation of carotenoids and lipids (Fazeli et al. 2006; Lamers et al. 2008). Organic carbon sources enhance growth and carotegenisis in microalgae. A high carbon/nitrogen ratio elevates accumulation of lipids and carotenoids in microalgae. Nitrogen starvation especially in the late exponential phase contributes to an increase in the biomass production due to a transient increase in the rate of cell division, increase in cell size (Converti et al. 2009). Accumulation of beta-carotene is highest when growth rate is least and that is a major problem in the commercial production of beta-carotene. An increase in salinity levels correspond to a decrease in the production of lutein and xanthine, leading to accumulation of higher levels of beta-carotene (Gómez et al. 2003). In previous studies it was reported that glucose, nitrate and salinity had a significant effect on biomass and beta-carotene production (Chen et al. 2009; Wang et al. 2014; Morowvat and Younes 2016).
The present study devises an optimized mixotrophic medium to achieve maximum biomass, beta-carotene and lipid production from Dunaliella salina which is practically very difficult to achieve using an autotrophic medium. Additionally, mixotrophic mode of medium is comparatively more cost effective in large scale cultivation of microalgae than the other trophic modes. Major problem faced practically in media optimizations is that the production of secondary metabolites occurs under stressed conditions which lead to a proportional decrease in the yields of biomass production. In this study we have accessed the economic feasibility of a mixotrophic medium by using low nitrate and high salinity stress with optimal glucose concentrations to achieve highest lipids and beta-carotene yields with minimal negative impact on the biomass production. Effects of three independent variables i.e., glucose (5–15 g/L), potassium nitrate (0.1–5 g/L) and sodium chloride (10–40 g/L) were studied individually and in combination using BBD. Beta-carotene concentrations were quantified using spectrophotometer at 460 nm and lipid content was measured using gravimetric method. Biomass, beta-carotene, and lipid production results of experiments conducted with different combinations of glucose, potassium nitrate and sodium chloride were compiled to generate interdependency equations and theoretical maximum values of the output. The theoretical optimized values were further confirmed through experiments.
Materials and methods
Strains, culture media, and culture conditions
Dunaliella salina was procured from The Energy and Resources Institute, New Delhi, India (This strain is procured because of its ability to grow in extreme environmental conditions and they are widely utilized for production of important biochemicals, such as carotenoids). The micro-algal strain was cultivated in the Modified Johnson’s Media composed of (per litre): MgCl2·6H2O (1.5 g), MgSO4·7H2O (0.5 g), KCl (0.2 g), CaCl2·2H2O (0.2 g), KNO3 (0.5 g), NaHCO3 (0.043 g), KH2PO4 (0.035 g), NaCl (12 g), Glucose (5 g), 0.9 M FeCl3 solution (10 ml), Trace Metal solution H3BO3 (61 mg/L), (NH4)6Mo7O24·4H2O (38 mg/L), CuSO4·5H2O (6 mg/L), CoCl2·6H2O (5.1 mg/L), ZnCl2 (4.1 mg/L), MnCl2·4H2O (4.1 mg/L)) (10 mL). The experiment was performed at 25 °C and neutral pH for a time span of 21 days starting with an inoculum density of 2 × 105 cells/ml. Cultures were incubated under continuous illumination at a light intensity of 3000 lx.
Media optimization by Box Behnken design
Box Behnken design was applied to achieve the interdependence of three independent variables, glucose (X1), potassium nitrate (X2) and sodium chloride (X3), respectively on algal biomass and beta carotene production. Optimal production of biomass and beta carotene was attained using Statistical Package Design Expert Version 10.0.6 (Stat Ease, Inc. Minneapolis, MN, USA). Optimization study comprised of 15 set of experiments including three replicates for centre point with different combinations as per the design package.
The upper and lower limit of glucose (X1), potassium nitrate (X2) and sodium chloride (X3) were set as 5–15 g/L, 0.1–5 g/L, and 10–40 g/L, respectively (Table 1). Optimal output was recorded as yields of algal biomass (Y1) and beta carotene (Y2), respectively. A generalized equation for output in terms of an equation comprising a linear, quadratic and an interactive term is represented as:
In the above equation, βo represents a constant term, βi is a linear coefficient, βii is a quadratic coefficient and βij is a cross term coefficient. The terms Xi and Xj represent the upper and lower limit of the independent variables, and ɛ represents the residual error.
Statistical analysis by ANOVA
Regression models as per response surface methodology of Box Behnken design were determined in terms of linear, quadratic and interaction terms. Regression model was solved using Newton Ralphson method. Models were validated to be significant using Analysis of variance (ANOVA). Experiments were conducted in triplicates and mean values of replicates were presented. F value at probability P, were quantified to be 0.0135, 0.0117 and 0.0014 thus validating the models to be significant. Significance of linear and quadratic regression coefficients was studied using Students t test at a probability of 0.05.
Analytical methods
Dried weight of algal biomass was measured every alternate day for 21 days. Culture flasks were harvested and centrifuged at 4500 rpm, for 15 min. Pellet was washed twice with distilled water and centrifuged again to remove impurities and dried overnight at 95 °C in a hot air oven over night in a pre-weighed glass petri-plate. Optical density of D. salina cultures was noted every alternate day for 21 days using a UV/vis Spectrometer at 680 nm.
Total lipid content of culture samples for every set of experiment was determined using standard Folch Method. Dried micro algal biomass (1 g) was homogenized using an ultrasonicator for 5 min in a mixture of chloroform–methanol (1:2). For 1 g of biomass 20 ml of solvent was added. Mixture was vortexed at every 10-min interval for 30 min so that the solvent gets absorbed and biomass turns colourless. The mixture was then centrifuged at 2000 rpm for 5 min. Supernatant was collected in clean glass tubes and 1/4th its volume of distilled water was added to allow phase separation. Bottom organic phase was washed with 1/4th its volume of distilled water: methanol (1:1). Two phases were allowed to settle and separate. Solvent phase was transferred in pre weighed glass vials and dried using nitrogen gas.
Beta carotene extraction was performed using hexane: ethanol (1:2) and estimated at 450 nm using a UV/vis spectrometer. Following 21 days of experiment 1 ml of D. salina culture was centrifuged for 10 min at 3000 rpm. Supernatant was discarded and pellet was suspended in 3 ml of hexane: ethanol (1:2). Solution was vortexed to allow a uniform mixture and centrifuged at 3000 rpm for 10 min. There were two phases of supernatant, the upper hexane phase retained beta-carotene compounds that were determined spectrophotometrically at 450 nm. Total amount of beta-carotene was calculated as:
where, A450 nm = absorbance of the hexane phase sample at 450 nm. Beta-carotene values were then divided by the microalgal biomass content to attain the values in mg/g.
Results and discussions
Data analysis using Box Behnken design
Effect of three independent variables glucose (X1), potassium nitrate (X2), sodium chloride (X3) on biomass productivity (Y1), beta-carotene productivity (Y2) and lipid productivity (Y3) were assessed using a three level and three factors as per the response surface methodology of the Box Behnken design.
The design comprises of 15 experiments arranged randomly consisting of three centre points and twelve factorial points. The three replicate centre points (runs 1, 6 and 7) were applied to access the pure error sum of squares.
A regression model was built to evaluate the biomass productivity (Y1), presented through a second order polynomial equation:
where, X1, X2, X3 define the actual concentrations of glucose, potassium nitrate and sodium chloride, respectively.
The regression model to predict the beta-carotene productivity (Y2) is represented through a second order polynomial equation as follows:
The second order polynomial equation using the actual values as per the regression model to evaluate the lipid productivity (Y3) is presented as:
Coded values Y2 and Y3 in equations represented beta carotene and lipid productivity. Statistical significance of regression model was analyzed using F distribution value. Model can be validated if P value in F distribution analysis was less than 0.05. F value of the model was 8.86, 9.47 and 24.42 for biomass, beta-carotene and lipid productivity which imply that model was significant with chances of F value being this large due to noise in each case was almost negligible i.e., 1.35%, 1.17%, and 0.13%, respectively. Analysis of variance (ANOVA) for biomass, beta-carotene and lipid productivity was presented in Table 2. Results imply that model was significant as p-values are less than 0.05.
In case of biomass yield (Y1), lack of fit of F-value is 3.36 which are insignificant relative to pure error and chances of this lack of fit being due to noise are 23.78%. An insignificant lack of fit indicates that the model perfectly fits the experimental values. Lack of Fit of F-values for beta-carotene (Y2) and lipid (Y3) productivity of 5.89 and 3.14 with the chances of this lack of fit being due to noise is only 14.86% and 25.06%, respectively which implies that the lack of fit is insignificant and the regression model proposed fits experimental values.
Lack of fit of regression model is insignificant and to determine if model fits experimental values the R2 coefficient was measured. As per ANOVA Table 2, R2 value, adjusted R2 value and predicted R2 values of 0.9410, 0.8348 and 0.1905, respectively exhibit that regression model for biomass productivity fits experimental values. R2 value, adjusted R2 value and predicted R2 values for beta carotene (Y2) and lipid productivity respectively (Y3) were 0.9446, 0.8448, 0.1908 and 0.9778, 0.9377, 0.6977, respectively which implies that regression model for Y2 and Y3 fits the experimental values.
Numerical optimization
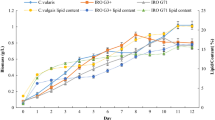
The results for 15 experimental runs were compiled using Box Behnken design. Three independent variables carbon, nitrogen and sodium chloride were altered to values as stated in Table 3, to attain theoretically possible maximal output responses. Numerical solution to get an optimized result for all three output results as per Response Surface Methodology based on Box Behnken design with a desirability of 93.8% is stated in Table 4. Theoretical maximum yield of biomass, beta-carotene and lipids as per Table 4, is 1.426 g/L, 6.707 mg/g and 21.67 mg/L/day, respectively and when glucose, potassium nitrate and sodium chloride were used at 13.23 g/L, 3.145 g/L and 35.6 g/L, respectively. Maximum yield of biomass, beta-carotene and lipids were 0.571 g/L, 4.18 mg/g and 13.2 mg/L/day, respectively. The numerical solution to get an optimized result for all the three output results as per Response Surface Methodology of Box Behnken design with a desirability of 93.8% is stated in the Table 4. Theoretical optimized culture conditions when performed experimentally exhibited 2.17 folds increase in biomass yield, 1.45 folds increase in beta-carotene yield and 1.56 folds increase in lipid yields respectively. Theoretical optimized medium was validated experimentally. The results of theoretical optimized medium were almost similar to the predictive optimized medium (1.42 g/L biomass, 5.32 mg/g beta-carotene, and 23.40 mg/L/day lipid) yields as exhibited in Run 8 of Table 5. This study validates the robustness in the use of Box Behnken design and response surface methodology as an important tool to optimize nutrient composition to overproduce biomass, beta-carotene and lipids simultaneously in D. salina.
Graphical representation
The optimal value for each independent variable to achieve maximal production of biomass, beta-carotene, and lipids, respectively, were derived by plotting Response surface 3D curves and contour curves. The surface 3D plots determined the interaction amidst two independent variables while keeping the third independent variable constant. As represented in Fig. 1a, b, higher glucose concentration results in a higher biomass yield whereas a moderate to high glucose concentration with a higher concentration of sodium chloride favours beta-carotene production as represented in Fig. 1e. This observation can be explained by the fact that a higher accumulation of beta-carotene defends the cell from adverse effects of salinity (Gómez et al. 2003). Wang et al. (2014) also observed that during mixotrophic cultivation, organic compounds such as glucose are accumulated through aerobic respiration and this phenomenon is affected, by the concentration of available organic carbon in the culture medium in case of Neochloris oleoabundans. Moreover, during aerobic respiration organic carbon sources are assimilated leading to a higher production of secondary metabolites (Smith et al. 2016). Morowvat and Younes (2016) also found the similar observations in case of Dunaliella salina. Nutrient stress imposed due to a lower nitrogen concentration allows an increase in beta carotene but affects the biomass yield as evident from Fig. 1d–g, respectively. A previous study reported that a decrease in nitrogen concentration results in an increase in cell size and a higher rate of proliferation thus exhibiting a higher cell density (Dominguez Teles 2016). As shown in Fig. 1f, g nitrogen limitation results an increase in lipid content. This can be justified by previous studies that show the conversion of nitrogen depleted cell components to lipids to release nitrogen (Fakhry and El Maghraby 2015). Skorupskaite et al. (2015) also found that increase concentration of ammonium nitrogen (more than 20 mg/L) might have adverse impacts on microalgae. Ahmed et al. 2017 reported maximum growth of Dunaliella salina obtained at 1.5–2 M NaCl. A higher amount of salinity imposes a nutrient stress on the microalgae Dunaliella salina and as evident from Fig. 1b, c, e, f, h, i, it favours beta-carotene and lipid content but affects biomass yield. This is evident by a study which reported, that an increase in salt stress increases the size of cell leading to a rapid uptake of the nutrients (Oren 2014). This leads to an increase in concentration of lipids per cell.

Response Surface 3D plots to study the effect of: a Glucose (X1) and potassium nitrate (X2) on biomass productivity (Y1). b Glucose (X1) and sodium chloride (X3) on biomass productivity (Y1). c Potassium nitrate (X2) and sodium chloride (X3) on biomass productivity (Y1). d Glucose (X1) and potassium nitrate (X2) on beta-carotene productivity (Y2). e Glucose (X1) and sodium chloride (X3) on beta-carotene productivity (Y2). f Potassium nitrate (X2) and sodium chloride (X3) on beta-carotene productivity (Y2). g Glucose (X1) and potassium nitrate (X2) on lipid productivity (Y3). h Glucose (X1) and sodium chloride (X2) on lipid productivity (Y3). i Glucose (X1) and sodium chloride (X3) on lipid productivity (Y3)
The optimal biomass and lipid yield of 1.42 g/L and 23.421.67 mgL/day were achieved in the experimental run 8 with an upper limit of glucose and sodium chloride, respectively. Minimum biomass production was observed in experimental run 15 where glucose to nitrogen ratio was low with a lower limit of carbon and an upper limit of nitrogen. The optimal yield of 7.04 mg/g of beta-carotene was achieved in experimental run 6 with a moderate concentration of carbon, nitrogen and sodium chloride. A nutrient scarcity as imposed in experimental run 14 with lower limits of glucose and nitrogen favours beta carotene production with a yield of 6.9 mg/g of biomass. A combination of lower limit of glucose and sodium chloride gives minimum yield of beta-carotene i.e., 2.54 mg/g. In order to achieve maximal yields of biomass, beta-carotene, and lipid, it is required to consider the concentration of three major elements i.e., carbon, nitrogen and salinity in a nutrient medium to culture the microalgae. The responses noted in Fig. 1a–i exhibit that concentration of three independent variables glucose, potassium nitrate, and sodium chloride, affect the yields of biomass, beta-carotene, and lipids, respectively.
Original concentration of glucose, potassium nitrate and sodium chloride in Modified Johnson’s Medium were 5 g/L, 0.5 g/L, and 12 g/L, respectively. Original media yields a total biomass productivity of 0.571 g/L with a beta-carotene content of 4.18 mg/g at the end of 21 days of experiment. Lipid content after 21 days of experiment in original medium as extracted through standard Folch Method was 13.2 mg/L/day. The optimized media as per theoretical results provided better results with a biomass yield of 1.42 g/L, beta-carotene content of 7.04 mg/g and a lipid content of 21.67 mg/L/day, at the end of 21 days, respectively whereas the maximum yield of biomass, beta-carotene and lipid productivity attained experimentally using the optimized media was 1.426 g/L, beta-carotene was 6.707 mg/g biomass and lipid productivity of 21.67 mg/L/day, respectively. Morowvat and Ghasemi (2016) obtained 8.03 mg/g of beta-carotene and 0.46 g/L/day of biomass production in Dunaliella salina. Mixotrophic cultivation mode has various advantages over other modes for increase production of biomass and lipid. It is almost hard to achieve higher amount of lipid and biomass productivity at the same time using autotrophic mode. However, mixotrophic cultivation attains higher amounts of biomass production with least negative impact on total lipid content of microalgae. To enhance the economic feasibility of beta-carotene production from D. salina it is important to improve the lipid formation and TAG accumulation. Therefore, utilization of environmental stress such as nutrient starvation can be a useful alternative (Converti et al. 2009; Markou and Nerantzis 2013). Here also we had evaluated the possibility of such nutrient starvation using carbon, nitrates and salt concentration in the culture medium.
Conclusion
The results imply that Box Behnken design can be successfully applied to optimize media for Dunaliella salina culture. Optimization study has formulated a media with concentrations of glucose (13.23 g/L), potassium nitrate (3.145 g/L), sodium chloride (35.6 g/L) while maintaining the concentration of other nutrient components unchanged. The current study proves to deliver promising results confirming an optimal yield of three products simultaneously. This study can be utilized for large production of biomass, lipid and beta carotene production.
References
Ahmed RA, He M, Aftab RA, Zheng S, Nagi M, Bakri R, Wang C (2017) Bioenergy application of Dunaliella salina SA 134 grown at various salinity levels for lipid production. Sci Rep 7(1):8118
Azma M, Mohamed MS, Mohamad R, Rahim RA, Ariff AB (2011) Improvement of medium composition for heterotrophic cultivation of green microalgae, Tetraselmis suecica, using response surface methodology. Biochem Eng J 53(2):187–195
Bonnefond H, Moelants N, Talec A, Mayzaud P, Bernard O, Sciandra A (2017) Coupling and uncoupling of triglyceride and beta-carotene production by Dunaliella salina under nitrogen limitation and starvation. Biotechnol Biofuels 10:25
Cheirsilp B, Torpee S (2012) Enhanced growth and lipid production of microalgae under mixotrophic culture condition: effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour Technol 110:510–516
Chen H, Jiang JG, Wu GH (2009) Effects of salinity changes on the growth of Dunaliella salina and its isozyme activities of glycerol-3-phosphate dehydrogenase. J Agric Food Chem 57(14):6178–6182
Chen M, Tang H, Ma H, Holland TC, Ng KY, Salley SO (2011) Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresour Technol 102(2):1649–1655
Converti A, Casazza AA, Ortiz EY, Perego P, Del Borghi M (2009) Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem Eng Process 48:1146–1151
Del Campo JA, García-González M, Guerrero MG (2007) Outdoor cultivation of microalgae for carotenoid production: current state and perspectives. Appl Microbiol Biotechnol 74:1163–1174
Dominguez Teles I (2016) The fatter the better: selecting microalgae cells for outdoor lipid production. Wageningen University. https://library.wur.nl/WebQuery/wurpubs/508992
Fakhry EM, El Maghraby DM (2015) Lipid accumulation in response to nitrogen limitation and variation of temperature in Nannochloropsis salina. Botanical studies 56(1):6
Fazeli MR, Tofighi H, Samadi N, Jamalifar H, Fazeli A (2006) Carotenoids accumulation by Dunaliella tertiolecta (Lake Urmia isolate) and Dunaliella salina (ccap 19/18 & wt) under stress conditions. DARU J Pharm Sci 14(3):146–150
García-González M, Moreno J, Manzano JC, Florencio FJ, Guerrero MG (2005) Production of Dunaliella salina biomass rich in 9-cis-beta-carotene and lutein in a closed tubular photobioreactor. J Biotechnol 115(1):81–90
Gary H, Masao O (2001) Impact of algal research in aquaculture. J Phycol 37:968–974
Gómez PI, Barriga A, Cifuentes AS, González MA (2003) Effect of salinity on the quantity and quality of carotenoids accumulated by Dunaliella salina (strain CONC-007) and Dunaliella bardawil (strain ATCC 30861) Chlorophyta. Biol Res 36(2):185–192
Hallenbeck PC, Grogger M, Mraz M, Veverka D (2015) The use of Design of Experiments and Response Surface Methodology to optimize biomass and lipid production by the oleaginous marine green alga, Nannochloropsis gaditana in response to light intensity, inoculum size and CO2. Bioresour Technol 184:161–168
Hosseini Tafreshi A, Shariati M (2009) Dunaliella biotechnology: methods and applications. J Appl Microbiol 107(1):14–35
Kirrolia A, Bishnoi NR, Singh R (2014) Response surface methodology as a decision-making tool for optimization of culture conditions of green microalgae Chlorella spp. for biodiesel production. Ann Microbiol 64(3):1133–1147
Lamers PP, Janssen M, De Vos RCH, Bino RJ, Wijffels RH (2008) Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol 26:631–638
Lamers PP, Janssen M, De Vos RCH, Bino RJ, Wijffels RH (2012) Carotenoid and fatty acid metabolism in nitrogen-starved Dunaliella salina, a unicellular green microalga. J Biotechnol 162(1):21–27
Markou G, Nerantzis E (2013) Microalgae for high-value compounds and biofuels production: a review with focus on cultivation under stress conditions. Biotechnol Adv 31(8):1532–1542. https://doi.org/10.1016/j.biotechadv.2013.07.011
Mata-Gómez LC, Montañez JC, Méndez-Zavala A, Aguilar CN (2014) Biotechnological production of carotenoids by yeasts: an overview. Microb Cell Fact 13:12
Mojaat M, Pruvost J, Foucault A, Legrand J (2008) Effect of organic carbon sources and Fe2+ ions on growth and β-carotene accumulation by Dunaliella salina. Biochem Eng J 39(1):177–184
Morowvat MH, Younes G (2016) Culture medium optimization for enhanced β-carotene and biomass production by Dunaliella salina in mixotrophic culture. Biocatal Agric Biotechnol 7:217–223
Oren A (2014) The ecology of Dunaliella in high-salt environments. J Biol Res 21(1):23
Rabbani S, Beyer P, Lintig J, Hugueney P, Kleinig H (1998) Induced beta-carotene synthesis driven by triacylglycerol deposition in the unicellular alga Dunaliella bardawil. Plant Physiol 116(4):1239–1248
Singh G, Jawed A, Paul D, Bandyopadhyay KK, Kumari A, Haque S (2016) Concomitant production of lipids and carotenoids in Rhodosporidium toruloides under osmotic stress using response surface methodology. Front Microbiol 7:1686
Skorupskaite V, Makareviciene V, Levisauskas D (2015) Optimization of mixotrophic cultivation of microalgae Chlorella sp. for biofuel production using response surface methodology. Algal Res 7:45–50
Smith RT, Bangert K, Wilkinson SJ, Gilmour DJ (2016) Synergistic carbon metabolism in a fast growing mixotrophic freshwater microalgal species Micractinium inermum. Biomass Bioenerg 2015(82):73–86
Srinivasan R, Kumar VA, Kumar D, Ramesh N, Babu S, Gothandam KM (2015) Effect of dissolved inorganic carbon on β-carotene and fatty acid production in Dunaliella sp. Appl Biochem Biotechnol 175(6):2895–2906
Sun XM, Ren LJ, Zhao QY, Ji XJ, Huang H (2018) Microalgae for the production of lipid and carotenoids: a review with focus on stress regulation and adaptation. Biotechnol Biofuels 11:272
Wang J, Yang H, Wang F (2014) Mixotrophic cultivation of microalgae for biodiesel production: status and prospects. Appl Biochem Biotech 172:3307–3329
Wu Z, Duangmanee P, Zhao P, Ma C (2016) The effects of light, temperature, and nutrition on growth and pigment accumulation of three Dunaliella salina strains isolated from saline soil. Jundishapur J Microbiol 9(1):1–9
Acknowledgements
This study was supported by Department of Biotechnology, Delhi Technological University, New Delhi.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ahuja, S., Roy, A., Kumar, L. et al. Media optimization using Box Behnken design for enhanced production of biomass, beta-carotene and lipid from Dunaliella salina. Vegetos 33, 31–39 (2020). https://doi.org/10.1007/s42535-019-00079-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42535-019-00079-4