
Abstract
Water and nutrients scarcity are the two major factors for yield reduction in many wheat-producing areas of the world. Drought stress also restricts micronutrient uptake and thus induces micronutrient deficiency. Zinc is the most important micronutrient for abiotic stress tolerance. A field trial was conducted from 2013 to 2015 to study the effect of foliar application of zinc under normal (100% field capacity) and water deficit (60% field capacity) conditions on quantitative and qualitative aspects of wheat (Triticum aestivum L. var. Faisalabad-08). Three Zn levels (0, 0.1%, and 0.2% by using ZnSO4. 7H2O) were foliar sprayed at the vegetative and reproductive stages of wheat. Obtained results revealed that the growth, yield traits, photosynthetic pigments, and WUE were negatively affected by lower water supply. Zinc foliar application did not affect yield under normal irrigation conditions, however, under water deficit foliar application of 0.2% Zn enhanced grain yield to 25–40%. Zinc foliar spray at both vegetative and reproductive stages increased the chlorophyll contents and WUE. Zinc contents of wheat grain were more when 0.1% Zn was sprayed at the reproductive stage or 0.2% Zn at the vegetative stage. Thus, the foliar application of Zn is a promising short-term approach to improve productivity and grain nutrient content in wheat under water deficit stress. Furthermore, the timing and rate of Zn application could be helpful for the efficient use of water for increasing grain yield.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wheat is one of the most important grain crops produced and traded globally. Consumption of wheat is increasing in the world (Shahbandeh, 2021) as a result of the increasing population (Ma et al. 2017), nutritional quality, and substitution with other cereals (Ram & Govindan, 2020). Therefore, increasing grain yield and improving the nutritional quality of wheat is still the object of numerous investigators. Drought and low availability of nutrients often limit the growth and productivity potential of wheat particularly in arid and semi-arid regions (Shayanmehr et al. 2020; Xue et al. 2019). The severity and duration of drought determine the extent of the yield loss by shortening the life cycle and duration of grain filling. A meta-analysis showed that drought caused a 27.5% reduction in wheat yield from 1980 to 2017 (Zhang et al. 2018). The models’ project showed continued increases in global drought frequency and area by 50–200% (in a relative sense) in the twenty-first century (Zhao & Dai, 2017). The future droughts are predicted to be more severe and frequent with more yield losses due to changing climate and variation in precipitation (Zhang et al. 2018).
There is extensive wheat yield variability in many semi-arid regions, which are owed to water limitation and year-to-year fluctuations in precipitation and temperature. The productivity of wheat growing in these environments not only depends on the water available for growth but also the water use efficiency (WUE) and amount of plant-available nutrients (Liang et al. 2019; Xue et al. 2019).
Drought disturbs the mineral-nutrient relations in plants through their effects on nutrient availability, and partitioning of transport processes in plants (Hu & Schmidhalter, 2005). It is extensively assumed that the mineral nutrient status of plants plays a main role in improving WUE and proper nutrition could alleviate water shortages and mitigate environmental problems (Ma et al. 2017; Waraich et al. 2011). Zinc fertilization improved the water use efficiency by increasing the osmolytes, maintaining membrane stability, and improving stomatal conductance (Hassan et al. 2020). Obtaining nutrient-enhanced grains requires the consideration of many factors, such as grain quality, bioavailability, and most importantly, grain yield (Shi et al. 2010). Accordingly, fertilizer management with macro and micronutrients in plant nutrition is very imperative for crop productivity and plants’ resistance (Janmohammadi et al. 2016).
Although micronutrients are required in small amounts, they play crucial roles in plant production at critical growth stages (Janmohammadi et al. 2016). Zinc fertilizer is widely used to improve yield, Zn concentration, and quality of edible parts of crops (Mousavi, 2011). It is well known that Zn has a great influence on basic plant life processes such as synthesis of chlorophyll, indole-3-acetic acid, nitrogen metabolism, protein quality, and enzyme activation (Zeidan et al. 2010). Zinc has been found useful in improving yield and yield components of wheat (Daneshbakhsh & Hossein, 2013; Ma et al. 2017; Zeidan et al. 2010) and adequately applied Zn has been shown to improve the water use efficiency of wheat plants (Bagci et al. 2007).
Zinc deficiency is more common in developing countries particularly in South Asia affecting human health and declining wheat productivity (Joy et al. 2017). Few studies have examined that the soil and foliar application of Zn increased the grain quality and yield traits of wheat and mobilization of Zn from vegetative tissues into the grains play an important role in biofortification (Cakmak et al. 2010; Ramzan et al. 2020). It has been reported that the concentrations of Zn and other micronutrients in cereal grains are significantly and positively correlated with those in leaves, suggesting the importance of foliar application of micronutrients for biofortification in grains (Xia et al. 2019). In addition, seedlings from Zn-enriched grains have a better ability to withstand environmental stresses at early growth stages (Mousavi, 2011).
Although Zn fertilizer is widely used, the effect of Zn application on the crop yield and Zn concentration are related to the amount and method of Zn fertilizer application (Wang et al. 2012). The forms and rates of fertilizer as well as timings affect the chemical composition of plants and therefore determine the technological utility and their feed value. The Zn remobilization from vegetative tissues into grain can be achieved by foliar applications of Zn (Cakmak et al. 2010; Xue et al. 2012). Foliar fertilization has the advantages of low application rates, uniform distribution, and quick plant responses to enrich essential micronutrients in crop grains (Saadati et al. 2013; Zhao et al. 2014). The time of foliar spray of Zn is the main factor determining the efficacy of Zn in improving grain Zn concentration (Nawaz et al. 2015). Foliar spray of Zn at a late growth stage and an early stage of seed development could be a useful practice to produce a significant increase in grain Zn concentration in wheat (Cakmak et al. 2000, 2010). Also, in young wheat, Zn is moved by phloem from leaves to roots after foliar application (Haslett et al. 2001) but in maturing wheat, zinc transported from leaves to developing grains (Riesen & Feller, 2005).
Combining improved tolerance to drought and increased Zn concentration and content in seeds is a high priority research topic, especially in Zn-deficient areas. However, finding a solution to Zn deficiency requires a better understanding and evaluation of the effect of foliar application of Zn and their role to enhance nutritional quality and yield of wheat. Therefore, this work aimed to evaluate the effects of different amounts and time of zinc spray on the quantitative and qualitative aspects of wheat under water stress conditions.
Materials and Methods
Experimental Conditions
A 2-year field experiment was conducted during 2013–15 in the research area at Faisalabad (31°N, 73.06°E) where the climate is semi-arid (Fig. 1). Seeds of high-yielding winter wheat variety (Triticum aestivum L. cv Faisalabad-08) were obtained from Ayub Agriculture Research Institute Faisalabad and sown by dibbling method on 15th October of the respective study years. The experiment was arranged in a randomized complete block design having four replications and each treatment was applied to the area of 1.5 m × 5 m. Physico-chemical properties of soil were determined before sowing as follows: sand = 49.5%, silt = 28.21%, clay = 22.29, saturation % = 40, EC = 1.01 dS m−1, pH = 7.06, organic matter = 0.25%, available nitrogen = 30 mg kg−1, available phosphorous = 7.04 mg kg−1, available potassium = 8 mg kg−1, and DTPA extractable Zn = 1.01 mg kg−1. Before sowing, basic doses of N (urea, 200 kg ha−1), phosphorous (KH2PO4, 60 kg ha−1), and potassium (K2SO4, 60 kg ha−1) were mixed in the soil. Additional 100 kg ha−1 nitrogen dose was also applied at tillering.

Meteorological condition of the research area during 2-year wheat growing season
Irrigation
Plants were subjected to two irrigation levels i.e., normal or fully irrigated (almost > 70% FC) and water deficit. The field capacity (FC) was measured before sowing at 30 cm soil depth for which the bare soil area was filled with excess water and coved with a plastic cover, then after 2 days the soil samples were collected from 30 cm soil depth and moist soil was weighed, dried in the oven at 105 °C till to constant weigh (after about 24 h) and then the dry soil was weighed to calculate soil moisture at 100% field capacity. For water deficit, the water available to the crop was limited (< 60% FC) by limiting irrigation.
Foliar Application of Zinc
Three levels of zinc, 0% (control), 0.1% (0.44 g /100 ml) and 0.2% (0.88 g/100 ml) were foliarly applied using zinc sulphate solutions (ZnSO4.7H2O, Sigma-Aldrich, USA) and adding 0.1% Tween-20. The foliar spraying was applied at 10 am at the vegetative (30 days after sowing) or reproductive stage (60 days after sowing).
Data Collection
Plants were harvested after 20 days of foliar application of zinc and at maturity. To record grain yield and yield-related traits, plants were harvested at maturity from 1 m2. Then samples were dried and digested with sulphuric acid and hydrogen peroxide at 350 °C (Lowther, 1980). Iron (Fe), zinc (Zn), and potassium (K) contents of grains and shoots were analyzed by atomic absorption (Jenway, PFP).
Water use efficiency (kg m−2) was calculated according to the method by Hussain and Al-Jaloud (1995) by using the formula:
WUE = grain yield (kg m–2)/amount of total water use for irrigation (m).
Data Analysis
The collected data was analyzed by Statistix 8.1 to determine the significance of treatment with analysis of variance (ANOVA), whereas, the significant differences between treatments were tested (P < 0.05) with the least significant difference (LSD). The figures were drawn by using Microsoft excel 2010.
Results
Yield and Biomass
The yield and yield-related traits were significantly affected by the drought and Zn foliar spray but the difference between the foliar spray at the vegetative and reproductive stage was not significant (Table 1). There was no significant difference in the yield and yield components between the control and Zn sprayed plants under full irrigation. Although, under water deficit, the grain yield was increased by increasing Zn foliar spray from 0.1 to 0.2%, the difference between 0.1 and 0.2% Zn was non-significant. The foliar application of 0.1 and 0.2% Zn at the vegetative stage showed a 10 and 38% increase in the first year and a 32 and 56% increase in the second year as compared to drought. Similarly, foliar application of 0.1 and 0.2% Zn at the reproductive stage caused a 24 and 34% increase in the first year and a 16 and 36% increase in the second year as compared to drought.
The grain weight, number of grains per spike, spikes per plant, and spikelets per spikes were significantly reduced under water deficit stress which showed 34%, 39%, 23%, and 28% reduction as compared to control (Table 1). Under drought, the foliar application of 0.2% Zn at the vegetative stage showed maximum increase in grains weight i.e., 24 and 27% increase in the first and second year respectively, but the effect was significantly similar to 0.1% Zn.
In the first year, a higher number of grains were obtained by foliar spray of 0.2% Zn at the vegetative stage both under full irrigation and drought conditions. Although, in the second year, the maximum number of grains were produced by 0.2% Zn foliar spray at the reproductive stage but it was statistically similar to 0.1% Zn. The number of spikes was not significantly affected by the Zn foliar spray. The number of spikelets was highest at 0.2% Zn at the vegetative stage both under control and drought conditions.
The aboveground dry biomass was significantly decreased under drought (Table 2). Under drought stress, shoot dry biomass was reduced to 3.3-fold and fourfold as compared to control in the first year and second year. In both years, shoot dry weight was not significantly increased with Zn foliar spray under fully irrigated conditions. Under drought stress, the shoot dry weight was significantly increased by the application of 0.1% Zn at the vegetative stage. The Zn spray at the vegetative stage showed more increase in shoot dry biomass than at the reproductive stage.
Water Use Efficiency
Water use efficiency was significantly reduced under drought conditions and was 27% and 33% less as compared to control in the first and second studied years. The effect of foliar application of Zn was significant in the first year, while non-significant in the second year. There was a 37% increase in WUE by the foliar application of 0.2% Zn at the vegetative stage and a 39% increase by 0.2% Zn at the reproductive stage as compared to drought.
Total Chlorophyll and Carotenoid Content
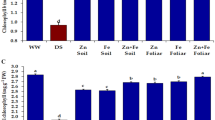
The water deficit caused a 24% and 26% reduction in total chlorophyll content of wheat flag leaves in the first and second year of study as compared to full irrigated plants (Fig. 2a). The total chlorophyll content was significantly increased by 0.2% Zn foliar application at the reproductive stage in the first year, while no significant increase was observed in the second year. While at water deficit stress, all applications of Zn increased the chlorophyll content and a maximum increase was observed at 0.2% Zn at the reproductive stage which showed 34% in the first year and 25% in the second year as compared to drought alone. The carotenoid content was significantly affected by drought and more reduction was observed in 2014–15 (Fig. 2b). Under normal irrigation, no significant effect of Zn spray was observed. While under drought, the carotenoid content was increased by the Zn spray, and a maximum increase was observed at 0.2% Zn at the reproductive stage in both years.

Effect of zinc foliar spray at the vegetative and reproductive stages of wheat on the a total chlorophyll and b carotenoid content after under drought stress. Each bar is a mean (n = 4) ± standard error and different lowercase letters on bars indicate significant differences among various irrigation and foliar treatments according to Fisher’s LSD test (p < 0.05)
Zinc Content in Shoot and Grain
The concentration of Zn in shoot and grains of wheat was increased by foliar application of Zn especially at the vegetative stage (Fig. 3). A notably higher Zn content in shoot and grains was found at 0.2% Zn at the vegetative stage in both fully irrigated and water deficit conditions. Under drought, the application of 0.2% Zn increased the Zn content in the shoot by 2.2-fold and 1.8-fold in the first year and 1.71-fold and 1.5-fold in the second year at the vegetative and reproductive stage, respectively (Fig. 3a). Under full irrigation, the Zn content in wheat grain was improved by increasing the Zn rate from 0.1 to 0.2% at the vegetative and reproductive stages (Fig. 3b). Under drought conditions, increasing Zn rate had increased the Zn content in grains at the vegetative stage while reduced at the reproductive stage. Under drought stress, the foliar application of 0.1% and 0.2% Zn at the vegetative stage caused a 17% and 19% increase in the first year and 35% and 44% increase in the second year and foliar application of 0.1% and 0.2% Zn at the reproductive stage caused 56% and 22% increase in the first year and 25% and 23% increase in the second year.

Effect of zinc foliar spray at the vegetative and reproductive stages on the zinc content in shoot and grain of wheat under drought. Each bar is a mean (n = 4) ± standard error and different lowercase letters on bars indicate significant differences among various irrigation and foliar treatments according to Fisher’s LSD test (p < 0.05)
Potassium Content in Shoot and Grain
Potassium (K) concentration in the shoot of fully irrigated plants was not affected by Zn spray at the vegetative spray, but 0.1% Zn at the reproductive stage in the first year and 0.2% Zn at the reproductive stage in the second year had significantly reduced the shoot potassium (Fig. 4a). Potassium content in shoot showed a 1.5-fold and 1.25-fold decrease under water deficit conditions as compared to control. The potassium content in grains was not significantly affected by Zn spray and drought stress (Fig. 4b). Under drought stress, only the application of 0.1% Zn spray at the reproductive stage in the second year showed a significant increase in the potassium content of the grains while other treatments showed a non-significant effect.

Effect of zinc foliar spray at the vegetative and reproductive stages on the potassium content in shoot and grain of wheat under drought. Each bar is a mean (n = 4) ± standard error and different lowercase letters on bars indicate significant differences among various irrigation and foliar treatments according to Fisher’s LSD test (p < 0.05)
Iron Content in Shoot and Grain
The shoot and grain iron (Fe) content were statistically similar for fully irrigated and drought stress (Fig. 5). There was no significant difference in shoot Fe content by Zn foliar application in the first year (Fig. 5a). While during the second year, 0.2% Zn sprayed at the reproductive stage had significantly increased the Fe content in shoot under drought stress. Under full irrigation, the Fe content in grains was significantly enhanced by the foliar spray of 0.1% and 0.2% Zn at the reproductive stage in first year, while not affected in the second year (Fig. 5b). Under drought, the application of 0.2% Zn at the reproductive stage caused a 33% increase in Fe content of grain in the first year, and 0.1% and 0.2% Zn at the reproductive stage caused a 32% and 55% increase in grain Zn content in the second year.

Effect of zinc foliar spray at the vegetative and reproductive stages on the iron content in shoot and grain of wheat under drought. Each bar is a mean (n = 4) ± standard error and different lowercase letters on bars indicate significant differences among various irrigation and foliar treatments according to Fisher’s LSD test (p < 0.05)
Discussion
Nutrient deficiencies are a worldwide nutritional problem for cereal production and the nutritional quality of the grains in many types of soil in many arid and semi-arid countries (Alloway, 2008). In this study, we observed drought caused a marked reduction in grain yields of wheat mainly due to less grain weight and number of grains. The zinc (Zn) foliar spray significantly increased the grain yield under drought stress particularly by increasing the number of grains under drought conditions. The increase in grain yield was 10–56% by 0.1% Zn spray, and 16–36% by 0.2% Zn under drought stress as compared to drought alone. This indicates that the Zn foliar application enhanced drought tolerance by maintaining leaf water status or by higher expression of aquaporins by Zn foliar spray (Fatemi et al. 2020). Previous studies also showed that Zn foliar application improved yield (Farooq et al. 2014; Karim et al. 2012). Since Zn is known as a critical micronutrient in plant growth and production, wheat dry weight may be improved through the Zn application under water stress and normal conditions (Saeidnejad et al. 2016; Xue et al. 2012). The effect of Zn treatment on shoot dry matter production (Table 2) was very similar to the effect on total chlorophyll (Fig. 2a). It seems that the availability of Zn resulted in higher photosynthetic activity, which in turn improved plant growth and ultimately production. Carbonic anhydrase, a Zn-containing metalloenzyme, is an essential enzyme for photosynthesis. The increase in carbonic anhydrase activity due to Zn application may be a factor contributing to higher photosynthesis, biomass, and yield (Bhat et al. 2017; Cakmak et al. 2000).
These findings are supported by Singh et al. (2012) who observed significantly higher plant growth by the application of Zn. Arough et al. (2016) reported that Zn could enhance the chlorophyll content and photosynthetic activity along with improved the growth of plants. It seems that a decrease in wheat yield under drought conditions related to increasing grain Zn and Fe concentration in the second year. These findings are reported by Peleg et al. (2008) who observed a negative association between wheat yield and grain mineral concentrations. The present study has also shown that the timing of the foliar application is an important factor in achieving the highest increases in shoot dry matter and grain Zn, Fe, and K through foliar Zn spray.
The need to improve agricultural water use efficiency (WUE) is rising due to the demand for enhanced grain production with using fewer resources (Waraich et al. 2011). The foliar application of Zn improved the WUE of the plants under water deficit conditions. Similar results have been made by Khan et al. (2003) in chickpea and Bagci et al. (2007) in wheat plants. It has been noticed that the foliar application of Zn alleviated the negative effects of water stress and significantly increased the WUE by activating the metabolic processes (Waraich et al. 2011) and by the osmotic adjustment in the plants (Hu & Schmidhalter, 2005). It seems that improvement of K content helps in stomatal regulation (Hu & Schmidhalter, 2005) and improves the chlorophyll and carotenoid content and lead to increase WUE under water deficit condition (Karim et al. 2012).
The reduction in chlorophyll and carotenoid content under water deficit is the most important anabolic process in plants resulting in reducing growth and yield (Hammad & Ali, 2014) which ultimately leads to a reduction in WUE (Waraich et al. 2011). Zinc plays an important role in shoot dry weight (Cakmak, 2008; Kaya & Higgs, 2002), chlorophyll production (Kaya & Higgs, 2002), and wheat yield (Farooq et al. 2014; Karim et al. 2012). Reduction in chlorophyll content at low Zn may be related to low Fe and Zn because Zn does not directly affect chlorophyll formation, but it can affect the concentration of Fe which are part of the chlorophyll molecule (Kaya & Higgs, 2002).
The Zn and Fe deficiency in cereal grains is the most common problem throughout the world. Water shortage could change the accumulation of micronutrients in grains (Singh et al. 2012). Our study showed that the Zn contents in grains were slightly increased while K and Fe contents were decreased by drought, but results were non-significant with the well-irrigated plants. Some researchers reported a decreased level of Zn under stress conditions (El-fouly et al. 2011), whereas others showed that Zn accumulation was improved due to stress (Saeidnejad et al. 2016). The concentrations of Zn in shoot and grains were stimulated through Zn foliar applications. Some previous results are in confirmation with the present results (Cakmak et al. 2010; Kutman et al. 2011). These results indicate that the foliar application of Zn at the vegetative and reproductive stages is translocated from vegetative organs to grain by phloem (Zhang et al. 2010). In addition, higher Zn in shoots also resulted in higher translocation of Zn to grains (Cakmak, 2008).
Our results showed that under drought stress, the foliar application of 0.1% Zn at the vegetative stage caused a 17–35% increase and 0.2% Zn caused a 19–44% increase, while foliar application of 0.1% Zn at the reproductive stage caused 25–56% increase and 0.2% Zn caused 22–23% increase in grain Zn. In our study, the grain Zn ranges from 29 to 34 mg kg−1 under full-irrigation which was increased to 41–51 mg kg−1 by foliar application of 0.1% Zn and 44 to 67 mg kg−1 by foliar application of 0.2% Zn. While under drought the grain Zn content ranges from 40 to 63 mg kg−1 by foliar application of Zn. Similarly, Gomez-Coronado et al. (2016) reported that total grain Zn in wheat varies from 11 to 52 mg kg−1 which was increased by 156% by the foliar application of Zn. To meet the recommended dietary allowance of Zn (15 mg kg−1), the total Zn level in grains should be in a range from 45 and 60 mg kg−1 (Ram & Govindan, 2020). Thus, increasing the Zn concentration in wheat grains to 40–60 mg kg−1 would meet the Zn requirement for people who consume cereal grains as a staple food (Cakmak, 2008).
The foliar application of Zn at the vegetative stage caused more accumulation of Zn in shoots and grains than at the reproductive stage. This might be because the reproductive stage might be late for the improvement in biomass under stress and translocation of Zn to grains. Therefore, biofortification of Zn contents in shoot and grain has been recommended by the application of Zn at the vegetative stage, particularly under drought condition. In this study, the relative increase in shoot Zn content under drought by 0.1% and 0.2% Zn rate was in a range of 16–119% (Fig. 2), whereas, under drought, the increase in shoot dry matter production by 0.1% and 0.2% Zn treatment were 54% and 154% higher (Table 2). While in well-irrigated plants, the shoot Zn content was increased but shoot dry biomass was not significantly increased by Zn spray. This showed that application of Zn only increased the biomass under drought and increased Zn application can be a major reason for increased shoot Zn content. These results also suggest that most of the grain Zn originates primarily from Zn translocation from vegetative tissues which is similar to the results presented by Haslett et al. (2001).
It seems that the environmental factors and growth conditions have a significant role in remobilization (contribution of phloem transport) and grain Zn accumulation (Boonchuay et al. 2013). High accumulation of Zn by application of Zn at vegetative probably because of a large Zn requirement for the rapidly growing tissue of wheat (Haslett et al. 2001). Zinc remobilization is the dominating supply of grain Zn in cereals even with high or low Zn status under control and water stress condition (Xue et al. 2012). It was assumed that in the second year, plants prefer to take up more Zn from Zn application at vegetable growth stages because of the warmer temperature and high humidity in the winter (Fig. 1). A similar observation was also found under field conditions in central Anatolia (Bagci et al. 2007).
Comparatively more Zn was accumulated in shoot and grains when Zn was sprayed at the vegetative stage as compared to Zn spray at the reproductive stage. This might be because Zn being highly mobile in phloem re-translocates considerably when the plants progress toward the reproductive stage (Riesen & Feller, 2005; Waters & Sankaran, 2011; Waters et al. 2009). In the second year of study (2014–15), an increase in Zn concentration by foliar spray of 0.2% Zn at the vegetative stage was 93% in aboveground dry biomass under full irrigation, whereas Fe was increased by 63% and K concentration increased by 19%. In the same condition, foliar application of 0.2% Zn at the vegetative stage increased Zn uptake by 132%, Fe uptake by 29%, and K uptake only 3% in grains under fully irrigation. These results showed that the element’s content in plant tissues increased with their foliar spray and then elements were redistributed in the grains. Phloem mobility of each element determines the amount of nutrient element remobilized via the phloem (Pahlavan-Rad & Pessarakli, 2009).
The current results indicate that the accumulation of K to the plant decreases with decreasing soil water content due to the decreasing mobility of K under water deficit conditions which was in agreement with Hu and Schmidhalter (2005) and Xue et al. (2012). The grain K concentration was significantly increased by 0.1% Zn spray at the reproductive stage in 2014–2015 at which the amount of grain K was increased by 93% as compared to the control under drought condition (Fig. 4). Whereas, the K content of the grain decreased to 32% by the application of a high Zn level (0.2%) at the reproductive stage as compared with the low-Zn (0.1%) treatment. It seems that the 0.1% Zn treatment was enough to maintain K remobilization from vegetative tissues into grains. The K concentration of shoot was also improved by 0.1% Zn foliar spray. These results showed the grain K contents under drought could be improved by improving the nutritional status of wheat and Zn has a role in the maintenance of a high K concentration in shoot and grain of wheat. It has been well-documented that adequate Zn nutrition is important in controlling the uptake of K and Fe by leaves (Cakmak, 2008; Xue et al. 2012). These findings are supported by Cakmak et al. (2000) who observed that K concentration in guard cells has been affected by Zn application. Potassium improves the drought tolerance in plants through its functions in the stomata regulation, maintains turgor pressure, and reduces transpiration under drought conditions (Hu & Schmidhalter, 2005).
The current results showed that grain Fe content was positively related to shoot Fe content (Fig. 5), indicating that grain Fe concentrations might be improved by increasing shoot Fe concentrations. It seems that foliar application of high-Zn treatment led to increase shoot dry matter and chlorophyll content especially under drought condition and consequently resulted in higher grain Fe content. Many previous studies reported a positive relationship between grain Zn concentration and grain Fe concentration in cereals (Kutman et al. 2011; Peleg et al. 2008; Xia et al. 2019). The synergetic effect of Zn and Fe accumulation in grain might be attributed to the pleiotropic effects or linkage among the genes governing Zn and Fe accumulation in grains (Xia et al. 2019). Notably, the effects of foliar application of Zn on the shoot and grain Fe concentration depend on the irrigation and timing of application. The decline in Fe concentration of shoot in the first year with the addition of high rates of Zn at the reproductive stage suggests reduced Fe availability (Khurana & Chatterjee, 2001) or transportation of Fe to the grains (Mirzapour & Khoshgoftar, 2006).
Conclusion
Results showed that water deficiency caused low grain yield by declining growth, number of grains, spikes per plant, spikelets per spike, grain weight, and WUE. The wheat plants have a differential response to Zn depending on the time of application and treatment concentration. The foliar application of 0.2% of Zn at the vegetative growth stage increased shoot Zn content by 93% and increased grain Zn concentration by 131%. At water deficit, higher accumulation of K was observed at 0.1% Zn at the vegetative stage while higher grain Fe was observed at 0.2% Zn at the reproductive growth stage. Overall, 0.2% Zn at the vegetative stage of wheat induced drought tolerance with the improved shoot and grain Zn content, chlorophyll content, shoot biomass, grain weight, and grain yield. Although, the total amount of Zn, Fe, and K in grains is not always an important indicator of their bioavailability (utilization), thus more bioavailability of nutrient grain resulting from foliar spray would be an important research topic in the future.
Availability of Data and Material
Not applicable.
Code Availability
Not applicable.
References
Alloway, B.J. (2008). Zinc in soils and crop nutrition (second edn, pp. 15–35). Brussels, Belgium and Paris, France: International Zinc Association (IZA) and International Fertilizer Association (IFA)
Arough, Y. K., Sharifi, R. S., Sedghi, M., & Barmaki, M. (2016). Effect of zinc and bio fertilizers on antioxidant enzymes activity, chlorophyll content, soluble sugars and proline in triticale under salinity condition. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 44(1), 116–124.
Bagci, S. A., Ekiz, H., Yilmaz, A., & Cakmak, I. (2007). Effects of zinc deficiency and drought on grain yield of field-grown wheat cultivars in Central Anatolia. Journal of Agronomy and Crop Science, 193, 198–206. https://doi.org/10.1111/j.1439-037X.2007.00256.x
Bhat, F. A., Ganai, B. A., & Uqab, B. (2017). Carbonic anhydrase: Mechanism, structure and importance in higher plants. Asian Journal of Plant Science Research, 7(3), 17–23.
Boonchuay, P., Cakmak, I., Rerkasem, B., & Prom-U-Thai, C. (2013). Effect of different foliar zinc application at different growth stages on seed zinc concentration and its impact on seedling vigor in rice. Soil Science and Plant Nutrition, 59, 180–188. https://doi.org/10.1080/00380768.2013.763382
Cakmak, I. (2008). Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant and Soil, 302, 1–17. https://doi.org/10.1007/s11104-007-9466-3
Cakmak, I., Kalayci, M., Kaya, Y., Torun, A. A., Aydin, N., Wang, Y., Arisoy, Z., Erdem, H., Yazici, A., Gokmen, O., Ozturk, L., & Horst, W. J. (2010). Biofortification and localization of zinc in wheat grain. Journal of Agricultural and Food Chemistry, 58, 9092–9102. https://doi.org/10.1021/jf101197h
Cakmak, I., Ozkan, H., Braun, H. J., Welch, R. M., & Romheld, V. (2000). Zinc and iron concentrations in seeds of wild, primitive, and modern wheats. Food and Nutrition Bulletin, 21, 401–403. https://doi.org/10.1177/156482650002100411
Daneshbakhsh, B., & Hossein, A. (2013). Effect of zinc nutrition on salinity-induced oxidative damages in wheat genotypes differing in zinc deficiency tolerance. Acta Physiologiae Plantarum, 35(3), 881–889. https://doi.org/10.1007/s11738-012-1131-7
El-fouly, M. M., Mobarak, Z. M., & Salama, Z. A. (2011). Micronutrients (Fe, Mn, Zn) foliar spray for increasing salinity tolerance in wheat Triticum aestivum L. African Journal of Plant Science, 5, 314–322.
Farooq, M., Hussain, M., & Siddique, K. H. M. (2014). Drought stress in wheat during flowering and grain-filling periods. Critical Reviews in Plant Sciences, 33, 331–349. https://doi.org/10.1080/07352689.2014.875291
Fatemi, H., Zaghdoud, C., Nortes, P. A., Carvajal, M., & Martínez-Ballesta, M. D. C. (2020). Differential aquaporin response to distinct effects of two Zn concentrations after foliar application in pak choi (Brassica rapa L.) plants. Agronomy, 10(3), 450.
Gomez-Coronado, F., Poblaciones, M. J., Almeida, A. S., & Cakmak, I. (2016). Zinc (Zn) concentration of bread wheat grown under Mediterranean conditions as affected by genotype and soil/foliar Zn application. Plant and Soil, 401(1–2), 331–346.
Hammad, S. A. R., & Ali, O. A. M. (2014). Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Annals of Agricultural Sciences, 59, 133–145. https://doi.org/10.1016/j.aoas.2014.06.018
Haslett, B. S., Reid, R. J., & Rengel, Z. (2001). Zinc mobility in wheat: Uptake and distribution of zinc applied to leaves or roots. Annals of Botany, 87, 379–386. https://doi.org/10.1006/anbo.2000.1349
Hassan, M. U., Aamer, M., Chattha, M. U., Haiying, T., Shahzad, B., Barbanti, L., Nawaz, M., Rasheed, A., Afzal, A., Liu, Y., & Guoqin, H. (2020). The critical role of zinc in plants facing the drought stress. Agriculture, 10(9), 396.
Hu, Y., & Schmidhalter, U. (2005). Drought and salinity: A comparison of their effects on mineral nutrition of plants. Journal of Plant Nutrition and Soil Science, 168, 541–549. https://doi.org/10.1002/jpln.200420516
Hussain, G., & Al-Jaloud, A.A. (1995). Effect of irrigation and nitrogen on water use efficiency of wheat in Saudi Arabia. Agricultural Water Management, 27(2), 143–153.
Janmohammadi, M., Amanzadeh, T., Sabaghnia, N., & Dashti, S. (2016). Impact of foliar application of nano micronutrient fertilizers and titanium dioxide nanoparticles on the growth and yield components of barley under supplemental irrigation. Acta Agriculturae Slovenica, 107, 265–276. https://doi.org/10.14720/aas.2016.107.2.01
Joy, E. J., Ahmad, W., Zia, M. H., Kumssa, D. B., Young, S. D., Ander, E. L., Watts, M. J., Stein, A. J., & Broadley, M. R. (2017). Valuing increased zinc (Zn) fertiliser-use in Pakistan. Plant and Soil, 411(1–2), 139–150.
Karim, M. R., Zhang, Y. Q., Zhao, R. R., Chen, X. P., Zhang, F. S., & Zou, C. Q. (2012). Alleviation of drought stress in winter wheat by late foliar application of zinc, boron, and manganese. Journal of Plant Nutrition and Soil Science, 175, 142–151. https://doi.org/10.1002/jpln.201100141
Kaya, C., & Higgs, D. (2002). Response of tomato (Lycopersicon esculentum L.) cultivars to foliar application of zinc when grown in sand culture at low zinc. Scientia Horticulturae, 93, 53–64. https://doi.org/10.1016/S0304-4238(01)00310-7
Khan, H. R., McDonald, G. K., & Rengel, Z. (2003). Zn fertilization improves water use efficiency, grain yield and seed Zn content in chickpea. Plant and Soil, 249, 389–400. https://doi.org/10.1023/A:1022808323744
Khurana, N., & Chatterjee, C. (2001). Influence of variable zinc on yield, oil content, and physiology of sunflower. Communications in Soil Science and Plant Analysis, 32, 3023–3030.
Kutman, U. B., Yildiz, B., & Cakmak, I. (2011). Improved nitrogen status enhances zinc and iron concentrations both in the whole grain and the endosperm fraction of wheat. Journal of Cereal Science, 53, 118–125. https://doi.org/10.1016/j.jcs.2010.10.006
Liang, Y. F., Khan, S., Ren, A. X., Lin, W., Anwar, S., Sun, M., & Gao, Z. Q. (2019). Subsoiling and sowing time influence soil water content, nitrogen translocation and yield of dryland winter wheat. Agronomy, 9(1), 37. https://doi.org/10.3390/agronomy9010037
Lowther, J. R. (1980). Use of a single sulphuric acid-hydrogen peroxide digest for the analysis of Pinus radiata needles. Communications in Soil Science and Plant Analysis, 11(2), 175–188.
Ma, D., Sun, D., Wang, C., Ding, H., Qin, H., Hou, J., Huang, X., Xie, Y., & Guo, T. (2017). Physiological responses and yield of wheat plants in zinc-mediated alleviation of drought stress. Frontiers in Plant Science, 8, 1–12. https://doi.org/10.3389/fpls.2017.00860
Mirzapour, M. H., & Khoshgoftar, A. H. (2006). Zinc application effects on yield and seed oil content of sunflower grown on a saline calcareous soil. Journal of Plant Nutrition, 29, 1719–1727. https://doi.org/10.1080/01904160600897430
Mousavi, S. R. (2011). Zinc in crop production and interaction with phosphorus. Australian Journal of Basic and Applied Sciences, 5, 1503–1509.
Nawaz, F., Ashraf, M. Y., Ahmad, R., Waraich, E. A., Shabbir, R. N., & Bukhari, M. A. (2015). Supplemental selenium improves wheat grain yield and quality through alterations in biochemical processes under normal and water deficit conditions. Food Chemistry, 175, 350–357. https://doi.org/10.1016/j.foodchem.2014.11.147
Pahlavan-Rad, M. R., & Pessarakli, M. (2009). Response of wheat plants to zinc, iron, and manganese applications and uptake and concentration of zinc, iron, and manganese in wheat grains. Communications in Soil Science and Plant Analysis, 40, 1322–1332. https://doi.org/10.1080/00103620902761262
Peleg, Z., Saranga, Y., Yazici, A., Fahima, T., Ozturk, L., & Cakmak, I. (2008). Grain zinc, iron and protein concentrations and zinc-efficiency in wild emmer wheat under contrasting irrigation regimes. Plant and Soil, 306, 57–67. https://doi.org/10.1007/s11104-007-9417-z
Ram, S., & Govindan, V. (2020). Improving wheat nutritional quality through biofortification. Wheat quality for improving processing and human health (pp. 205–224). Springer.
Ramzan, Y., Hafeez, M. B., Khan, S., Nadeem, M., Batool, S., & Ahmad, J. (2020). Biofortification with zinc and iron improves the grain quality and yield of wheat crop. International Journal of Plant Production, 14, 510.
Riesen, O., & Feller, U. (2005). Redistribution of nickel, cobalt, manganese, zinc, and cadmium via the phloem in young and maturing wheat. Journal of Plant Nutrition, 28, 421–430. https://doi.org/10.1081/PLN-200049153
Saadati, S., Moallemi, N., Mortazavi, S. M. H., & Seyyednejad, S. M. (2013). Effects of zinc and boron foliar application on soluble carbohydrate and oil contents of three olive cultivars during fruit ripening. Scientia Horticulturae, 164, 30–34. https://doi.org/10.1016/j.scienta.2013.08.033
Saeidnejad, A. H., Kafi, M., & Pessarakli, M. (2016). Interactive effects of salinity stress and Zn availability on physiological properties, antioxidant activity, and micronutrients content of wheat (Triticum aestivum) plants. Communications in Soil Science and Plant Analysis, 47, 1048–1057. https://doi.org/10.1080/00103624.2016.1165831
Shahbandeh, M. (2021). Global wheat cinsumption. https://www.statista.com/statistics/1094056/total-global-rice-consumption/. Accessed 10 Feb 2021
Shayanmehr, S., Henneberry, R. S., Sabouni, S. M., & Foroushani, S. N. (2020). Drought, climate change, and dryland wheat yield response: An econometric approach. International Journal of Environmental Research and Public Health, 17(14), 5264.
Shi, R., Zhang, Y., Chen, X., Sun, Q., Zhang, F., Römheld, V., & Zou, C. (2010). Influence of long-term nitrogen fertilization on micronutrient density in grain of winter wheat (Triticum aestivum L.). Journal of Cereal Science, 51, 165–170. https://doi.org/10.1016/j.jcs.2009.11.008
Singh, S., Gupta, A. K., & Kaur, N. (2012). Influence of drought and sowing time on protein composition, antinutrients, and mineral contents of wheat. The Scientific World Journal. https://doi.org/10.1100/2012/485751
Wang, J., Mao, H., Zhao, H., Huang, D., & Wang, Z. (2012). Different increases in maize and wheat grain zinc concentrations caused by soil and foliar applications of zinc in Loess Plateau, China. Field Crops Research, 135, 89–96. https://doi.org/10.1016/j.fcr.2012.07.010
Waraich, E. A., Ahmad, R., Yaseen Ashraf, M., Saifullah, S., & Ahmad, M. (2011). Improving agricultural water use efficiency by nutrient management in crop plants. Acta Agriculturae Scandinavica Section B, 61, 291–304. https://doi.org/10.1080/09064710.2010.491954
Waters, B. M., & Sankaran, R. P. (2011). Moving micronutrients from the soil to the seeds: Genes and physiological processes from a biofortification perspective. Plant Science, 180, 562–574. https://doi.org/10.1016/j.plantsci.2010.12.003
Waters, B. M., Uauy, C., Dubcovsky, J., & Grusak, M. A. (2009). Wheat (Triticum aestivum) NAM proteins regulate the translocation of iron, zinc, and nitrogen compounds from vegetative tissues to grain. Journal of Experimental Botany, 60, 4263–4274. https://doi.org/10.1093/jxb/erp257
Xia, H., Kong, W., Wang, L., Xue, Y., Liu, W., Zhang, C., Yang, S., & Li, C. (2019). Foliar Zn spraying simultaneously improved concentrations and bioavailability of Zn and Fe in maize grains irrespective of foliar sucrose supply. Agronomy, 9(7), 386.
Xue, L., Khan, S., Sun, M., Anwar, S., Ren, A., Gao, Z., Lin, W., Xue, J., Yang, Z., & Deng, Y. (2019). Effects of tillage practices on water consumption and grain yield of dryland winter wheat under different precipitation distribution in the loess plateau of China. Soil and Tillage Research, 191, 66–74.
Xue, Y. F., Yue, S. C., Zhang, Y. Q., Cui, Z. L., Chen, X. P., Yang, F. C., Cakmak, I., McGrath, S. P., Zhang, F. S., & Zou, C. Q. (2012). Grain and shoot zinc accumulation in winter wheat affected by nitrogen management. Plant and Soil, 361, 153–163. https://doi.org/10.1080/01904169709365267
Zeidan, M. S., Mohamed, M. F., & Hamouda, H. A. (2010). Effect of foliar fertilization of Fe, Mn and Zn on wheat yield and quality in low sandy soils fertility. World Journal of Agricultural Sciences, 6, 696–699.
Zhang, J., Zhang, S., Cheng, M., Jiang, H., Zhang, X., Peng, C., Lu, X., Zhang, M., & Jin, J. (2018). Effect of drought on agronomic traits of rice and wheat: A meta-analysis. International Journal of Environmental Research and Public Health, 15(5), 839.
Zhang, Y., Shi, R., Rezaul, K. M., Zhang, F., & Zou, C. (2010). Iron and zinc concentrations in grain and flour of winter wheat as affected by foliar application. Journal of Agricultural and Food Chemistry, 58, 12268–12274. https://doi.org/10.1021/jf103039k
Zhao, A., Tian, X., Cao, Y., Lu, X., & Liu, T. (2014). Comparison of soil and foliar zinc application for enhancing grain zinc content of wheat when grown on potentially zinc-deficient calcareous soils. Journal of the Science of Food and Agriculture, 94, 2016–2022.
Zhao, T., & Dai, A. (2017). Uncertainties in historical changes and future projections of drought. Part II: Model-simulated historical and future drought changes. Climate Change, 144, 535–548. https://doi.org/10.1007/s10584-016-1742-x
Funding
This work received no external funding.
Author information
Authors and Affiliations
Contributions
R.B, ZUN: Data curation, Formal analysis and Investigation; S.A: Writing-Original draft preparation; S.K: Methodology; A.M: Resources, conceptualization; R.K: Software; A.P, R.K: Writing-review and editing.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no conflict of interest.
Rights and permissions
About this article

Cite this article
Anwar, S., Khalilzadeh, R., Khan, S. et al. Mitigation of Drought Stress and Yield Improvement in Wheat by Zinc Foliar Spray Relates to Enhanced Water Use Efficiency and Zinc Contents. Int. J. Plant Prod. 15, 377–389 (2021). https://doi.org/10.1007/s42106-021-00136-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42106-021-00136-6