
Abstract
Plant feeding by omnivorous predators may affect their development and survival rate, as well as their dispersal behavior after release in the target patch. In this study, we determined whether soybean or corn seedlings could be used as insectary plants to enhance the establishment of Arma chinensis and improve its suitability as a natural enemy of pest insects. We tested the development, nymphal survival rate, predation rate, number of eggs laid, and dispersal of A. chinensis on soybean, corn, and tobacco in laboratory and field experiments. Results showed that A. chinensis reared on soybean developed faster and had greater survival rates during the nymphal stage as compared to those reared on corn and tobacco. Adult A. chinensis fed least on Spodoptera litura larvae and laid the greatest number of eggs on soybean seedlings. Arma chinensis dispersed faster and spent less time in the greenhouse when provided with corn as a host, while the retention time of A. chinensis in the target field was extended when provided with soybean as a host. In addition, a more uniform distribution of A. chinesis on a higher proportion of plants was observed when soybean was provided as a host. These results indicate that soybean is a potential insectary plant that could be used during the early stages of A. chinensis release when prey is scarce or absent.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Augmentative biological control using natural enemies has been practiced for decades to control the main insect pests in greenhouses and outdoor crops (van Lenteren et al. 2018). One important factor affecting biological control efficiency is how well a population of released natural enemies establishes in a crop. Many programs have failed because natural enemies did not successfully establish a population in the target patch (Collier and Van Steenwyk 2004). Crop diversification increases natural enemy abundance and therefore reduces pest populations, and intercropping is an important tool to achieve this spatial diversity of crops (Smith & McSorley 2000). Rational intercropping can also improve crop yield (Alarcón‐Segura et al. 2022) as well as decrease the occurrence of pests and diseases. Thus, studies on potential intercropping patterns to promote the natural enemies’ efficacy are essential for planning effective biological control programs.
Omnivorous natural enemies can feed on both prey and plants (Eubanks et al. 2003) and have been used successfully in the biological control of many crop pests (Blubaugh et al. 2016; Leman et al. 2020). Studies have shown that plant feeding positively affected the performance of omnivorous predators (Limburg and Rosenheim 2001, Portilloet al. 2012, Pumariño et al. 2012, Oveja et al. 2016). The ability of omnivorous Hemiptera (bugs) to feed on plants may allow them to survive and establish populations when prey is scarce or absent and thus prevent pest densities from reaching economic thresholds. However, previous studies on how plant feeding affects prey consumption have given inconsistent results. In most cases, prey feeding was reduced when high quality plant food was provided (Eubanks and Denno 2000; Shakya et al. 2009; Lykouressis et al. 2014; Stephan et al. 2017); in other systems, a positive or no relationship between plant feeding and prey consumption was reported (Gillespie and McGregor 2000; Blubaugh et al. 2016).
Dispersal is another key factor that affects the establishment of natural enemies. A high rate of dispersal can lead to an Allee effect whereby too few individuals remain in the target area after release for a population to persist (Heimpel and Asplen 2011). Host plant attributes such as plant structure and nutrition affected the dispersal of natural enemies (Coll et al. 1997; Eubanks and Denno 1999). For omnivorous predators, availability of high-quality plant resources within the target patch may reduce dispersal and increase the likelihood of establishing a resident population.
Arma chinensis Fallou (Hemiptera: Pentatomidae: Asopinae) is an omnivorous predator found in China, Japan, and North Korea. This predator has a broad prey range (Pan et al. 2019) that makes it a promising biological control agent in ephemeral and disturbed agroecosystems. Arma chinensis successfully developed and reproduced feeding on prey only; however, the availability of plants was crucial for its establishment (Pan, unpublished data).
We have used A. chinensis to control Lepidoptera pests, including Spodoptera litura and S. exigua, on tobacco which is an important economic crop in China (Shen et al. 2018). However, the density of A. chinensis decreased quickly as individuals dispersed from the release sites and the efficiency of pest control was not good due to the slow establishment of A. chinensis in annual crops. In this case, early population establishment on the crop is needed to use A. chinensis as a biological control agent. Dispersed A. chinensis were found on both tobacco and non-crop plants. Soybean and corn which are commonly intercropped with tobacco are species on which A. chinensis was found feeding and egg laying (Tu et al. 2015; Zhou et al. 2015). We observed that starved A. chinensis preferred feeding on these plants even when prey was available. We hypothesize that intercropping with soybean and corn could function to retain released A. chinensis within tobacco crops, enhance the establishment of A. chinensis populations, and improve its suitability as a biological control agent.
The objective of this study was to evaluate the potential of soybean and corn as insectary plants to enhance the performance of A. chinensis in biological control programs. We tested the influence of soybean, corn, and tobacco on the performance and dispersal of A. chinensis. Experiments were conducted to determine (1) the effects of plant species on the development, survival, and prey feeding of A. chinensis, (2) the dispersal of A. chinensis on three plant species, and (3) the effects of soybean on the dispersal of A. chinensis in tobacco fields.
Materials and methods
Experimental plants and insects
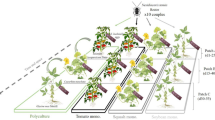
Soybean, corn, and tobacco plants were grown from seeds. The seeds were sown in plastic plates (length 50 cm × width 35 cm × height 10 cm) filled with vermiculite.
Arma chinensis was collected in Langfang, Hebei province, and reared on Bombyx mori pupae in the laboratory at 25 ± 2 °C, 60 ± 10% RH, and a photoperiod of 16: 8 (L: D). First instar nymphs of A. chinensis from one egg mass were placed in a transparent plastic cup and provided with water via a piece of moist absorbent cotton. One Chinese oak silk moth pupa was provided as food from the second instar and replaced every 4 to 5 d.
Spodoptera litura larvae were reared on artificial diet at 25 ± 2 °C, 70 ± 10% RH, and a photoperiod of 14: 10 (L: D). Adults were provided with 10% honey-water solution, and absorbent gauze was placed in the cages for oviposition.
Development and survival of larval A. Chinensis
Twenty newly hatched A. chinensis were put in a transparent plastic cage (length 14.3 cm × width 8.5 cm × height 5.5 cm) with a mesh top and provided with fresh plant leaves. The leaves were replaced every two days. Nymphs of A. chinensis that developed to the second instar were provided with both plant leaves and S. litura larvae of third to fifth instar as food. The survival and development stage of A. chinensis nymphs were recorded every day until they developed to adults. Five replicates were used for each plant species (100 newly hatched nymphs were used in total for each plant treatment).
Prey feeding and oviposition of A. Chinensis
Soybean, corn, and tobacco seedlings were planted individually into plastic pots (12 cm diam.) of soil mix and grown to 30 cm high for experiments. Five pots of the same species were placed in a cage (60 cm × 60 cm × 60 cm), and 15 adults of A. chinensis (10 females and 5 males) that had emerged within five days were then released in the cage. The A. chinensis were provided with 30 third- to fourth-instar larvae of S. litura every day. The number of S. litura killed and eggs laid by A. chinensis was recorded each day for continuous five days. Five replicates were used for each plant species.
Dispersal and distribution of A. Chinensis in greenhouse experiments
The dispersal of A. chinensis on the three plant species was tested in a 35 m2 greenhouse (7× 5 m). A total of 70 potted plants of each species were placed in seven rows in the greenhouse (10 plants per row) with 60 cm between each plant. Plants were approximately 25 cm high. Individuals of A. chinensis could pass freely in and out of the greenhouse through a narrow gap between the window and roof. At 7 pm, 150 A. chinensis adults were released in the center of the greenhouse. One corner of the greenhouse was set as the coordinate origin, and the position of each A. chinense was measured with a Handheld Laser Rangefinder (Leica D2, Switzerland) at 7 am, 11 am, 3 pm, and 7 pm daily for three days. Three replicate experiments were conducted for each plant species.
The diffusion constants were calculated with the formula (Heinz 1998):
where x and y are the planar coordinates describing the position of an individual in the greenhouse; xo and yo are the coordinates of the release point; n is the number of A. chinensis found in the greenhouse; ∆t is the time after release.
To determine the resultant distribution pattern of released A. chinense, the greenhouse was divided into 35 units of 1 m2. Data of the number of A. chinensis in each unit from the three replicate experiments were combined and used to calculate the distribution patterns of A. chinensis on each of the three plant species at 48 h after release using the index of clumping (IDM) (David and Moore 1954):
where s2 is variance and \(\overline{x }\) is mean number of A. chinensis per unit.
Dispersal of A. Chinensis in tobacco fields
To test the dispersal of A. chinensis in tobacco fields with and without soybean plants, we selected two plots of approximately 667 m2 each. Within each plot, the distance between rows was 1 m, and the distance between plants in each row was 0.5 m. The release point of A. chinense was the center of the field. One potted soybean (about 30 cm in height) was placed at the release point of one plot. One hundred adults of A. chinensis were released in each plot at 8 am. We surveyed the distribution of A. chinensis at 12, 36, 60, and 84 h after release and recorded the numbers of A. chinensis at release point and on the tobacco seedlings and weeds within 10 m (we observed the maximum dispersal distance of A. chinensis was 8.1 m). The experiment was conducted 3 times (replicates).
Data analysis
All data were analyzed using statistics package SPSS v. 19.0 (SPSS Inc., Chicago, IL, USA). An analysis of variance (one-way ANOVA) was used to compare the survival and development time during nymphal stages, host feeding, and the number of eggs laid by adult A. chinensis, when reared on different host species; means were separated using Tukey–Kramer HSD test at P < 0.05. The effects of host plant species on diffusion constants, the proportion of A. chinensis on plants, and the proportion of A. chinensis remaining in the greenhouses were determined by Kruskal–Wallis nonparametric tests. The Mann–Whitney U test was used to test for a significant difference in the number of A. chinensis collected from tobacco fields with and without soybean at the release site.
Results
Development and survival
The development time of A. chinensis varied greatly among plant species. The first-instar nymphs which do not feed on prey developed fastest on soybean and slowest on tobacco (F = 104.492, df = 2, 274, P < 0.0001); the duration from second instar to both male (F = 3.254, df = 2, 78, P = 0.044) and female (F = 5.981, df = 2, 64, P = 0.004) adults was also affected by plant species. Arma chinensis reared on soybean developed fastest and those reared on corn slowest; no significant difference in development time from second instar to adult was found between individuals reared on tobacco and those reared on the other two plant species. The mean survival rate of A. chinensis nymphs reared on soybean and corn was 64% and 49%, respectively, significantly higher than that of nymphs reared on tobacco (36%); there was no significant difference in survival rate between individuals reared on the former two plant species (F = 5.583, df = 2, 12, P = 0.019) (Table 1).
Prey feeding and oviposition
Arma chinensis on tobacco and corn fed significantly more S. litura than those on soybean (F = 6.564, df = 2, 12, P = 0.012); however, the difference in average number of prey consumed by one bug in one day was 0.2. The average number of eggs laid by A. chinensis reared on soybean was twofold greater than those reared on tobacco, but the difference in the number of eggs laid by A. chinensis on the three plant species was not significant in statistics (F = 3.375, df = 2, 12, P = 0.069), (Table 2).
Dispersal and distribution of A. Chinensis in greenhouse experiments
Host plant species significantly affected diffusion constants of A. chinensis. Arma chinensis dispersed much more quickly on corn than on the other two plant species at 16 h (χ2 = 7.200, df = 2, 7, P = 0.027), 20 h (χ2 = 7.200, df = 2, 7, P = 0.027), 36 h (χ2 = 7.200, df = 2,7, P = 0.027), 40 h (χ2 = 7.200, df = 2, 7, P = 0.027), 68 h (χ2 = 6.489, df = 2, 7, P = 0.039), and 72 h (χ2 = 7.200, df = 2, 7, P = 0.027) (Fig. 1A). There was a significant difference in the percentage of A. chinensis collected on the three plant species only at 24 h after release (χ2 = 6.543, df = 2, 7, P = 0.038) (Fig. 1B). The proportion of A. chinensis on soybean seedlings was in the range of 58–76%, while only 7–34% of A. chinensis were on tobacco; the differences were significant at 12 h (χ2 = 6.489, df = 2, 7, P = 0.039), 16 h (χ2 = 7.200, df = 2, 7, P = 0.027), 20 h (χ2 = 7.200, df = 2, 7, P = 0.027), and 24 h (χ2 = 6.489, df = 2, 7, P = 0.039) after release (Fig. 1C).

Effects of host plant species on (A) diffusion constants, (B) proportion of A. chinensis collected, and (C) proportion of A. chinensis distributed on plants. (Kruskal–Wallis nonparametric test, *P < 0.05)
The distribution of A. chinensis was affected by host plant species. Although A. chinensis always showed a clumped distribution on the three plant species, a more uniform distribution of A. chinensis was found when soybean was provided as a host. The IDM of A. chinensis on tobacco (18.15) at 48 h after release was 2.3 and 4 times greater than on corn (7.85) and soybean (4.53), respectively. About 67% of individuals were distributed on the window when corn was provided as host. When tobacco was provided, A. chinensis aggregated around the release point; about 40% of individuals were distributed within one-meter radius of the release point at 72 h after release (Fig. 2).

Distribution of A. chinensis in greenhouse on three plant species at 24, 48 and 72 h after release
Dispersal of A. Chinensis in tobacco fields
Soybean plants significantly extended the retention time of A. chinensis in the tobacco fields. Although the difference in the total number of A. chinensis collected at different times after release between fields with and without soybean was not statistically significant (12 h: U = 0.5, Z = − 1.771, P = 0.077; 36 h: U = 0, Z = − 1.964, P = 0.05; 60 h: U = 0, Z = − 1.964, P = 0.05; 84 h: U = 0, Z = − 1.964, P = 0.05), many more individuals were collected from fields with soybean than from fields without soybean (Fig. 3).

Mean (± SE) number of A. chinensis collected at different times after release in tobacco fields with and without soybean plants. (Mann–Whitney U test, *P < 0.05)
Discussion
Our results show that the species provided as insectary plants influenced the suitability of A. chinensis as a biological control agent. Although A. chinensis is a prey-feeding omnivore, plants are also an important nutritional resource; therefore, plant attributes could directly affect their suitability as we have found. Arma chinensis reared on soybean developed faster and survived more often during the nymphal stage as compared to those reared on corn and tobacco. These results are not surprising. Previous studies indicated that soybean is a highly suitable host for both nymphal development and adult reproduction of several insect species, including Nezara viridula (Velasco and Walter 1992) and Bemisia tabaci (Musa and Ren 2005). Conversely, tobacco was shown to be an unsuitable host. Previous studies have shown this is because it contained the alkaloid nicotine which was highly toxic to herbivores as well as their natural enemies (Barbosa et al. 1986; Thorpe and Barbosa 1986). Plants such as tobacco may indirectly affect the suitability of A. chinensis by influencing the quality of S. litura as prey as herbivores can sequester defensive compounds from plants to resist their natural enemies (Ode 2006). Similarly, the average number of eggs laid by adult A. chinensis fed on soybean was about twofold higher than by those fed on tobacco.
The differences in development and survival of A. chinensis were most pronounced in the first instar; the development time of first instar nymphs on tobacco was one day longer than those developed on soybean, and more than 20% of individuals died on tobacco during first instar stage. This is probably because A. chinensis feeds on plants only during the first instar, while they feed on both plants and prey from the second instar. Plants may play a bigger role in affecting the suitability of A. chinensis as a biological control agent during the first instar.
Many studies have indicated that providing omnivorous predators with a high-quality plant-based resource reduced prey consumption because of the additional nutrition from the plants (Eubanks and Denno 2000; Shakya et al. 2009; Frank et al. 2011; Lykouressis et al. 2014; Stephan et al. 2017). We also found in this study that adult A. chinensis consumed significantly less prey on soybean than on corn and tobacco. Both plant morphology and chemistry affect predation by natural enemies. Results from our first experiment suggest that soybean is a good nutritional resource for A. chinensis, and soybean-provided nutrients resulted in reduced need of nutrients from prey. However, as Kaplan and Thaler (2011) reviewed, many highly resistant and low-quality plants also decreased prey consumption by omnivorous predators, and this was probably due to these plants having glandular trichomes which negatively affected predation efficiency.
Although A. chinensis reared on soybean consumed less S. litura larvae than those reared on corn and tobacco, the difference in average number of prey consumed by one bug in one day was 0.2, which is a very small number. However, as we found in this study, soybean as alternative nutrient resources had significantly positive effects on the development, larval stage survival and reproduction of A. chinensis, and these positive effects would enhance its suitability as a biological control agent. How soybean-feeding affects pest control efficiency of A. chinensis needs further study.
Apart from affecting the development, survival, number of eggs laid, and predation efficiency, plant species also affected the dispersal rate and distribution of A. chinensis after release. Arma chinensis dispersed faster and spent less time in the greenhouse when provided with corn as a host. This behavior of A. chinensis may reduce the probability that its population will establish in corn patch. A more uniform distribution was observed when soybean was provided as a host, and there were always about 60% of individuals distributed on soybean plants. A more uniform distribution and higher proportion distributed on plants could increase the probability of encountering prey and further improve predation efficiency (Alatawi et al. 2011; Put et al. 2012). In addition, soybean plants significantly extended the retention time of A. chinensis in the tobacco field. These results indicate that soybean could function as insectary plants and enhance population establishment of A. chinensis in target patches. As we found in the first experiment, tobacco was not a suitable host for A. chinensis, and the predator rarely fed on tobacco leaves. Moreover, as we observed, the young leaves of tobacco secrete mucus that would hinder the movement of A. chinensis. This may explain why we found only a small proportion on the tobacco plants.
In conclusion, soybean plants could improve the suitability of A. chinensis as a biological control agent by reducing its reliance on prey and extending its retention time in the target field. Thus, tobacco could be intercropped with soybean to enhance population establishment of A. chinensis, especially at the early stages after introduction of A. chinensis when prey is scarce or absent. However, further research is needed to determine how intercropped soybean plants affects pest suppression by A. chinensis in the field.
References
Alarcón-Segura V, Grass I, Breustedt G, Rohlfs M, Tscharntke T (2022) Strip-intercropping of wheat and oilseed rape enhances biodiversity and biological pest control in a conventionally managed farm scenario. J Appli Ecol 21:1–11
Alatawi F, Nechols JR, Margolies DC (2011) Spatial distribution of predators and prey affect biological control of two spotted spider mites by Phytoseiulus persimilis in greenhouses. Bio Control 56:36–42
Barbosa P, Saunders JA, Kemper J, Trumbule R, Olechno J, Martinat P (1986) Plant allelochemicals and insect parasitoids effects of nicotine on Cotesia congregata (Say) (Hymenoptera: Braconidae) and Hyposoter annulipes (Cresson) (Hymenoptera: Ichneumonidae). J Chem Ecol 12:1319–1328
Blubaugh CK, Hagler JR, Machtley SA, Kaplan I (2016) Cover crops increase foraging activity of omnivorous predators in seed patches and facilitate weed biological control. Agr Ecosyst Environ 231:264–270
Coll M, Smith LA, Ridgway RL (1997) Effect of plants on the searching efficiency of a generalist predator: the importance of predator-prey spatial association. Entomol Exp Appl 83:1–10
Collier T, Van Steenwyk R (2004) A critical evaluation of augmentative biological control. Biol Control 31:245–256
David FN, Moore PG (1954) Notes on contagious distribution in plant populations. Ann Bot 18:47–53
Eubanks MD, Denno RF (1999) The ecological consequences of variation in plants and prey for an omnivorous insect. Ecology 80:1253–1266
Eubanks MD, Denno RF (2000) Host plants mediate omnivore–herbivore interactions and influence prey suppression. Ecology 81:936–947
Eubanks MD, Styrsky JD, Denno RF (2003) The evolution of omnivory in heteropteran insects. Ecology 84:2549–2556
Frank SD, Shrewsbury PM, Denno RF (2011) Plant versus prey resources: influence on omnivore behavior and herbivore suppression. Biol Control 57:229–235
Gillespie DR, McGregor RR (2000) The functions of plant feeding in the omnivorous predator Dicyphus hesperus: water places limits on predation. Ecol Entomol 25:380–386
Heimpel GE, Asplen MK (2011) A ‘Goldilocks’ hypothesis for dispersal of biological control agents. Biocontrol 56:441–450
Heinz KM (1998) Dispersal and dispersion of aphids (Homoptera: Aphididae) and selected natural enemies in spatially subdivided greenhouse environments. Envir Entomol 27:1029–1038
Kaplan I, Thaler JS (2011) Do plant defenses enhance or diminish prey suppression by omnivorous Heteroptera? Biol Control 59:53–60
Leman A, Ingegno BL, Tavella L, Janssen A, Messelink GJ (2020) The omnivorous predator Macrolophus pygmaeus, a good candidate for the control of both greenhouse whitefly and poinsettia thrips on gerbera plants. Insect Science 27:510–518
Limburg DD, Rosenheim JA (2001) Extrafloral nectar consumption and its influence on survival and development of an omnivorous predator, larval Chrysoperla plorabunda (Neuroptera: Chrysopidae). Envir Entomol 30:595–604
Lykouressis D, Perdikis D, Charalampous P (2014) Plant food effects on prey consumption by the omnivorous predator Macrolophus pygmaeus. Phytoparasitica 42:303–309
Musa PD, Ren SX (2005) Development and reproduction of Bemisia tabaci (Homoptera: Aleyrodidae) on three bean species. Insect Sci 12:25–30
Ode PJ (2006) Plant chemistry and natural enemy fitness: effects on herbivore and natural enemy interactions. Annu Rev Entomol 51:163–185
Oveja MF, Riudavets J, Arnó J, Gabarra R (2016) Does a supplemental food improve the effectiveness of predatory bugs on cucumber? Biocontrol 61:47–56
Pan MZ, Zhang HP, Zhang LS, Chen HY (2019) Effects of starvation and prey availability on predation and dispersal of an omnivorous predator Arma chinensis Fallou. J Insect Behav 32:134–144
Portillo N, Alomar O, Wäckers F (2012) Nectarivory by the plant-tissue feeding predator Macrolophus pygmaeus Rambur (Heteroptera: Miridae): Nutritional redundancy or nutritional benefit? J Insect Physiol 58:397–401
Pumariño L, Alomar O (2012) The role of omnivory in the conservation of predators: Orius majusculus (Heteroptera: Anthocoridae) on sweet alyssum. Biol Control 62:24–28
Put K, Bollens T, Wäckers FL, Pekas A (2012) Type and spatial distribution of food supplements impact population development and dispersal of the omnivore predator Macrolophus pygmaeus (Rambur) (Hemiptera: Miridae). Biol Control 63:172–180
Shen S, Xu G, Chen F, Gu X, Ji S, Zhang L, Wu D (2018) Effects of aphidius gifuensis release on insect communities and diversity in tobacco fields of Yunnan Province, China. Pak J Biol Sci: PJBS 21(6):284–291
Shakya S, Weintraub PG, Coll M (2009) Effect of pollen supplement on intraguild predatory interactions between two omnivores: the importance of spatial dynamics. Biol Control 50:281–287
Smith HA, McSorley R (2000) Intercropping and pest management: a review of major concepts. Am Entomol 46(3):154–161
Stephan JG, Stenberg JA, Björkman C (2017) Consumptive and nonconsumptive effect ratios depend on interaction between plant quality and hunting behavior of omnivorous predators. Ecol Evol 7:2327–2339
Thorpe KW, Barbosa P (1986) Effects of comsumption of high and low nicotine tobacco by Manduca sexta (Lepidoptera: Sphingidae) on survival of gregarious endoparasitoid Cotesia congregata (Hymenoptera: Braconidae). J Chem Ecol 12:1329–1337
Tu Y, Yang WY, Liu WG, Yong TW, Wang XC (2015) Effects of relay strip intercropping years between flue-cured tobacco and soybean on rhizospheric microbes quantities. Acta Agron Sin 41(5):733–742 ((in Chinese))
van Lenteren JC, Bolckmans K, Köhl J, Ravensberg WJ, Urbaneja A (2018) Biological control using invertebrates and microorganisms: plenty of new opportunities. Biocontrol 63:39–59
Velasco LRI, Walter GH (1992) Availability of different host plant species and changing abundance of the polyphagous bug Nezara viridula (hemiptera: pentatomidae). Environ Entomol 4:751–759
Zhou G, Yin X, Li Y, Zhao Z, Xu L, Ding J (2015) Optimal planting timing for corn relay intercropped with flue-Cured tobacco. Crop Sci 55:2852–2862
Ting Z, Banjie L, Binghui Z, Shun X, Gang Gu (2020) The research progress of tobacco pest control by intercropping. Chin Tob Sci 41(5):105–112
Acknowledgements
We are grateful for the help of Haiping Zhang from the Sino-American Biological Control Laboratory, Institute of Plant Protection, Beijing, China, in conducting the experiments. We also thank Zhaozhi Lu, Lynda Perkins and anonymous reviewers for their valuable comments and improving the English text. This research was supported by the Projects of Guizhou Tobacco Corporation (201936, 201937, and 201941) and the Qingdao Agricultural University High-level Talent Fund (665-1117002 and 663-1119003).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Pan, MZ., Fu, ZX., Li, YY. et al. Role of host plants in the suitability and dispersal of an omnivorous predator Arma chinensis Fallou (Hemiptera: Pentatomidae: Asopinae) in a biological control context. J Plant Dis Prot 129, 861–868 (2022). https://doi.org/10.1007/s41348-022-00624-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41348-022-00624-5