
Abstract
We examined if restored connectivity within a freshwater coastal wetland, dominated by invasive Typha x glauca, can increase the ecological condition, as assessed by a Floristic Quality Index (FQI), and improve habitat to benefit northern pike (Esox lucius) reproduction. Using an aquatic excavator to simulate natural rivulets, channels were excavated within monotypic Typha stands to create a link to upslope wet meadows. Following excavation, we found channels were rapidly colonized by native vegetation, resulting in higher FQI scores and percent cover of short-emergent vegetation compared to natural reference channels. Excavated channels also retained a community of vegetation with more native species five years following excavation. Fish emigration surveys revealed that excavated channels functioned as early-life habitat and had higher abundances of young-of-the-year northern pike than reference channels. A seven-year dataset (2011–2017) of northern pike outmigration at channel connectivity and pothole complexes was strongly related to water level conditions during the spawning and emigration period. We conclude that the enhancement methods increased the ecological condition of the vegetation community (i.e., FQI) suppressed by invasive Typha while the associated northern pike reproductive success highlights the importance of habitat connectivity and periodic flooding within coastal wetlands.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ecological evaluations of enhanced wetlands are well documented for salt marshes (Minello and Webb 1997; Dionne et al. 1999; Williams and Zedler 1999; Havens et al. 2002; Larkin et al. 2008; Weinstein and Litvin 2016). Less research, however, has been applied to habitat restoration within coastal wetlands in freshwater systems, especially for wetland-dependent fishes (e.g., Oele et al. 2015; Landress 2016), within a context of hydrodynamic regulation (Poff et al. 1997; Freeman et al. 2001) and invasive Typha (Boers et al. 2007). The regulated hydrology of Lake Ontario and the St. Lawrence River (LOSLR) and its coastal wetlands provides an opportunity to examine these management influences in a large freshwater system. The Robert Moses Saunders Power Dam in the upper St. Lawrence River is used to regulate discharge to manage water levels in adherence of policies set by the International Joint Commission (IJC) through its St. Lawrence River Board of Control. Dam operations from 1958 to 2017 followed the regulation mandate of Plan 1958D until Plan 2014 was enabled in January of 2017. Past regulation is known to have suppressed the magnitude and periodicity of water level fluctuations (Farrell et al. 2010) and system-wide ramifications to multiple interests have been demonstrated (IJC 2014). Coastal wetlands throughout the region experienced a shift in vegetation composition to those dominated by invasive cattail Typha x glauca (Godr.) resulting in reduction of wet meadow habitats (Wilcox et al. 2008; Farrell et al. 2010; Rippke et al. 2010; Wilcox and Bateman 2018). Typha x glauca is the hybrid of the native broad-leaved (T. latifolia (L.)) and the introduced narrow-leaved cattail (T. angustifolia (L.)) (Smith 1967). Because T. x glauca exhibits hybrid heterosis and spreads vegetatively, it can produce large dense mats (Galatowitsch et al. 1999) and may flourish with water level fluctuation suppression (Boers and Zedler 2008; Wilcox et al. 2008). Thus, under Plan 1958D, T. x glauca has spread and accumulated litter (Vaccaro et al. 2009).
The expansion and dominance of Typha within coastal wetlands, along with system hydro-pattern alteration, has demonstrated impacts on wetland dependant fishes, as connectivity and accessibility to coastal wetland habitat is critical to their life-cycle (Jude and Pappas 1992). Northern pike (Esox lucius), a top predator in the St. Lawrence River, (Farrell et al. 2006) preferentially spawns over flooded wet meadow habitats during spring before juveniles emigrate to deeper waters (Farrell 2001; Cooper et al. 2008). Although northern pike will spawn in deeper water when access to preferred spawning habitat is blocked (Farrell 2001; Cooper et al. 2008), early-life survival and recruitment is low from these habitats (Farrell et al. 2006). Thus, lost access to historical reproductive habitats (e.g. wet meadows; Farrell 2001; Cooper et al. 2008) from the expansion of T. x glauca and changes to hydrodynamics are thought to be associated with reduced reproductive success, contributing to a population decline of northern pike in the upper St. Lawrence River (Smith et al. 2007).
Increased efforts are underway to enhance freshwater coastal wetlands impacted by invasive Typha to improve lateral floodplain connectivity and provide access for spawning fish. Excavated channels and pool complexes have been created in areas blocked by Typha in an effort to reconnect mainstream tributaries with remnant upslope wet meadow habitat. Channel excavation has also been implemented to promote colonization of native vegetation from the seed bank buried beneath the accumulated Typha litter (Lishawa et al. 2015). Greenhouse experiments previously demonstrated that buried seed banks in coastal wetlands of the St. Lawrence River remain viable despite the spread of T. x glauca (Farrell et al. 2010). What remains uncertain is whether native vegetation can colonize channels once the seed bank is exposed, or if T. x glauca and its accumulated litter will inhibit colonization of native vegetation (Farrer and Goldberg 2009; Larkin et al. 2012). Excavated channels may promote native vegetation colonization that was replaced by T. x glauca expansion and improve the ecological condition from increased cover of beneficial plant species within and along created channels (Schummer et al. 2012). Consequently, improved vegetative condition could enhance spawning and early nursery habitat opportunities for northern pike (Bry 1996).
We therefore examined vegetation response of channel excavation compared with existing reference channels that received no enhancement with emphasis on northern pike reproduction and physical and environmental habitat conditions. We also used a longer term dataset from excavation and reference sites to examine the effect of hydrologic conditions (e.g. water levels) on northern pike emigration abundance.
Methods
Study Area
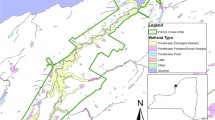
French Creek is a drowned river mouth tributary within the Thousand Islands region of the upper St. Lawrence River (Fig. 1) and serves as important coastal wetland spawning and nursery habitat for northern pike. The main stem of French Creek is characterized by a mix of submersed aquatic vegetation (SAV; e.g., Myriophyllum spicatum, Zizania palustris) and floating-leaved vegetation (e.g., Nymphaea odorata and Hydrocharis morsus ranae). Within the floodplain and along the edges of the main channel, dense stands (~10–20 stems per m2) of robust-emergent vegetation dominate (e.g., primarily T. x glauca and T. angustifolia), while narrow bands of wet meadow habitat, primarily sedges (Carex spp.) occur at groundwater seeps and rivulets upslope that became inaccessible to fish by Typha expansion (Farrell et al. 2010).

French Creek showing excavated channels with reconnected wet meadow habitat; stars indicate locations of juvenile fish emigration nets at exits of enhancement channels (n = 5) and reference channels (n = 2) and locations of vegetation transects are depicted by the circled V. Location of reference channels (R) indicated with an arrow and labelled with whether the reference channels were surveyed for emigrating northern pike (n), vegetation (v) or both (nv). Bottom insets provide 2015 orthoimagery (orthos.dhese.ny.gov) of an excavated channel and spawning pool complex
To restore connectivity and accessibility to wet meadow habitat by fish, enhancement channels were created in French Creek with an amphibious aquatic excavator by the US Fish and Wildlife Service, Partners for Fish and Wildlife Program, New York Field Office, Cortland NY. Channel excavation sites were chosen based on proximity to wet meadow habitat to the main channel and where historical aerial imagery identified previous channels that had been filled by Typha. The first two enhancement channels, 164 and 256 m length, were excavated in fall 2008 and reconnected French Creek with wet meadow approximately 1531 m2 and 3202 m2 respectively (Fig. 1). A substantially longer channel was excavated in fall 2010 measuring 807 m and reconnected 8457 m2 of wet meadow to the main stem (Fig. 1). All enhanced channels were designed to create some meandering and were about 1–2 m width, with an approximate depth of 0.5 m. Width and depth of the excavated channels approximated that at reference sites. Dredge spoil was side-casted along riparian zones. To evaluate excavated channels, existing channels in French Creek that remained unblocked, but bordered by Typha, were selected as reference sites (n = 3; Fig. 1). The reference sites were assumed to be representative of small connecting channels (lengths ranged between 98 and 218 m) available following Typha expansion. One reference channel was selected to survey both vegetation and emigrating northern pike, while two additional reference channels were used for either vegetation or emigrating northern pike sampling (Fig. 1).
Vegetation Sampling
The response of vegetation to channel excavation was compared between the first two excavated channels and two randomly selected reference channels for five consecutive years (2009–2013). Longitudinally, at each site, at least one 30.5 m-length channel segment was established, using a random number generator to determine a vegetation sampling transect location. The number of vegetation sampling transects established at each of four channels was in proportion to its total length. At each segment, a lateral transect that bisected the channel was established with five permanent vegetation plots (1 m2). Vegetation plots were positioned at channel midpoint, each riparian edge and 1.5 m from the riparian edge to capture vegetation response of its immediate floodplain. Overall, 95 permanent vegetation plots were established, 50 plots (10 transects × 5 plots per transect) at enhancement channels and 45 plots (9 transects × 5 plots per transect) at reference channels. All vegetation plots were sampled annually during peak growing season from 2009 to 2013, except in 2009 when the survey was done in September. In each quadrat we identified all vascular plants and visually estimated percent coverage for each species. We also counted individual live Typha stems and recorded maximum Typha height. All plants were identified to the lowest observable taxon and if necessary specimens were brought back to the lab for identification.
Vegetation Analyses
The vegetation response in riparian edge and floodplain plots, and channel habitat was analyzed separately. We calculated an overall importance value (IV) for each species as:
where for species i, IVi was defined as the sum of the relative frequency (Rfi) and the relative cover (RCi) of species i (Wilcox and Meeker 1991). Relative frequency (Rfi) was the sum of species incidence i in all plots divided by the sum of species occurrences in all plots. Relative cover was defined as the sum of cover of species i in all plots divided by the sum of cover of all species in all plots. Importance value for each species was averaged across the five years, ranked in order of highest to lowest and qualitatively compared between treatments.
Channel Vegetation
For each year and channel type (excavated and reference), we compared species proportions among six structural categories: algae (filamentous), macroalgae, floating-leaved, submersed fine-leaved, submersed broad-leaved, and emergent. An “other” category was included for plants not fitting a category. We also compared percent cover of combined submersed categories between treatments to infer fish habitat in the excavated channels.
Riparian Vegetation
The riparian “edge habitat” plots on each side of channels were averaged by transect and two “floodplain habitat” plots located 1.5 m landward from the channel’s edge were averaged. Each riparian habitat type was analyzed separately to determine if habitat responded differently based on distance from the channel. We also pooled data between edge and floodplain habitats to provide a representative riparian zone measure between channel types. Three metrics were used to evaluate vegetation response following excavation to infer habitat quality from a fisheries perspective, and included percent cover of robust-emergent vegetation, percent cover of short-emergent vegetation, and the Floristic Quality Index (FQI) score.
Percent cover of robust-emergent vegetation was evaluated as a physical barrier that impedes access to preferred spawning habitat for northern pike and consisted of Typha and Phragmites australis. Percent cover of short-emergent vegetation included sedges, grasses (e.g., Poaceae spp.) and rushes (e.g., Juncaceae spp.) (n = 33 taxa total) that were historically dominant prior to T. x glauca expansion (Rippke et al. 2010) and considered preferred spawning habitat for northern pike (Farrell 2001). Finally, the FQI was used to measure the overall ecological condition of wetlands that accounts for the entire vegetation species assemblage of a site within Great Lakes coastal wetlands (Bourdaghs et al. 2006).
The FQI was determined using coefficients of conservatism (C) that range in value between 0 and 10, and represent the estimated probability that a plant is likely to occur in an unaltered pre-European settlement landscape condition (Wilhelm 1977; Swink and Wilhelm 1994; Taft et al. 1997). Coefficients of conservatism of 0 are assigned to ubiquitously distributed plants, while a C of 10 is applied to plants that are nearly restricted to un-impacted habitat (pre-settlement remnant). FQI was calculated following:
where \( \overline{C} \) was the mean C score and S was the number of species. We used C scores from Michigan (Herman et al. 2001) because well-established C scores do not exist for New York State (Bried et al. 2012). Species without a listed C value were given the average C score for that genus, while exotic species received a score of 0. An FQI score was computed for each habitat type per transect and higher FQI scores represent higher quality habitat based on the product of community composition and species richness (i.e., ecological condition; Bourdaghs et al. 2006).
Vegetation metrics were compared between excavated and reference channels with linear mixed effect models (LMM) in lmerTest (version 3.0–1; Kuznetsova et al. 2017) (R package lme4 version 1.1–17; Bates et al. 2015). Channel type and year, and their interaction, were considered fixed effects, while specific transects (i.e., intercept) and year (i.e., slope) were set as random factors. Likelihood ratio tests were used in our selection of transect-by-year random slopes and intercept models over random intercept-only models due to their significantly better fit to the data for some habitat metrics (i.e., FQI scores and SAV cover).
Satterthwaite approximation procedure (lmerTest, R package lme4) was used to test for significant main effects and their interaction for all habitat variables at α = 0.05, with a primary focus to examine differences between the natural and restored channels. Channel type by year interactions, however, were plotted and visually inspected to evaluate general patterns in response of vegetation to excavation among years. Differences between channel type for each year was evaluated with independent mixed-effect ANOVAs where transects were a random factor nested within channel type.
Finally, to assess vegetation similarity between channel types, we used nonmetric multidimensional scaling (NMS) (PRIMER, version 6.1; Clarke and Gorley 2006). The Bray-Curtis method was used to determine community similarity and ordinate vegetation plot assemblages in two dimensions. One-way analysis of similarity (ANOSIM) was used with 9999 permutations to test for differences between assemblages. The ANOSIM test statistic, global R, was calculated following bootstrapping where the observed R value was compared against a random distribution of R to determine its significance (Clarke 1993). The global R values range from −1 to 1, with values closer to 1 indicating dissimilarity between groups, values near 0 indicating no differences between groups and values closer to −1 indicating similarity between groups (Clarke and Warwick 2001).
Northern Pike Response to Excavation
Emigrating northern pike were sampled during late spring and early summer of 2011, 2012, and 2013 with outmigration traps set at the openings of enhancement and reference channels. In 2011, a minnow trap with a 2.54 cm opening, wrapped and sewn with 0.8 mm nylon mesh to the top and center of a 0.8 mm nylon mesh panel 366 cm in length, 122 cm in height, was buoyed with floats and weighted with a lead core line. The trap was deployed with three reinforcing rods; two across the channel opening and one to the minnow trap stretched downstream in the channel center. For 2012 and 2013, emigration traps were modified to allow greater water flow and space in the holding pen to minimize fish mortality. The new trap consisted of three connected 60 cm diameter hoops with 2.54 cm throats, a mesh size of 1.6 mm and the same attachment design as the previous trap. Both traps spanned the entire channel opening to prevent immigration and capture all emigrating fishes. Traps were emptied every 24–48 h and all fish caught were identified to species, counted and released. Traps were deployed to coincide with the timing of northern pike juvenile outmigration and occurred from 2-June to 1-July in 2011, 23-May to 20-June in 2012 and 4-June to 1-July in 2013.
Comparisons of emigrating young-of-the-year (YOY) northern pike were made between channel types after catches were standardized by catch-per-unit-effort (CPUE; number of YOY northern pike caught per 24 h per net or net-night). For each year, daily CPUE was nested within channel type and treated as a random factor in a LMM with lmerTest (R package lme4) where channel type and year were fixed factors. Main effects and treatment-by-year interaction were assessed using the Satterthwaite procedure at α = 0.05.
Environmental variables were evaluated for differences between channel types for 2011, 2012 and 2013. Water temperature (± 0.1 °C) was recorded with Onset HOBO ProV2 data loggers at each net every two-hours from mid-depth. Recorded temperatures were then converted to a Mean Daily Temperature (MDT) and averaged by treatment per day. Dissolved oxygen concentration ([DO]) was measured with either the YSI 55 Dissolved Oxygen (polarographic sensor) or an YSI ProOdo (Optical sensor) meter at each daily emigration trap check. Dissolved oxygen was measured during daylight (0830 – 1600 h) from the surface, mid-depth and bottom of the water column. Dissolved oxygen concentrations were averaged from the three measurements and treatment specific daily means were used to compare channel types among years. Both MDT and [DO] were compared between channel type and among years in the same manner as CPUE of emigration YOY northern pike.
Finally, daily water levels (mIGLD85) representing the northern pike spawning and emigration period (1-April to 1-July) were downloaded from the NOAA monitoring station located in Alexandria Bay, NY. (ABAN6–8311062; https://tidesandcurrents.noaa.gov/waterlevels.html?id=8311062). Data was used to evaluate potential relationships between habitat connectivity and outmigration CPUE for 2011 and 2013, but a larger dataset from excavated channels (n = 3), reference sites (n = 11), and excavated spawning pool complexes (n = 5; Massa and Farrell, accepted pending revision) was compared for 2011 to 2017 that followed the same fish survey protocol. Spawning pool complexes were excavated within dense Typha along French Creek as a series of pools and connecting channels in enhancements led by Ducks Unlimited (Fig. 1) and were designed to improve habitat, create new northern pike spawning habitat, and increase connectivity to upslope wet meadows. Total area of habitat created by pools was 1.36 Ha, where individual pools averaged (±SE) 0.04 ± 0.005 Ha.
Hierarchal modeling was used to determine the influence of water levels on northern pike outmigration patterns with the “pcount” function of the R-package “unmarked” (version 0.12–2). Because pcount is based on a Poisson distribution, each site was considered a replicate (n = 19) and total abundance of juvenile northern pike caught during the entire outmigration period was the response. Due to differences in set duration of nets among years and sites, total net-nights per site per year was used in modeling to account for varying effort, as well as whether habitat type (i.e., excavated or reference) influenced outmigration. Thus, water levels and effort were considered observation-level covariates, while habitat type was a site-level covariate in modelling routines. Main-effect and additive models were compared using AICc with the R-package “AICmodavg” (Mazerolle 2017) to rank models, while Chi-square goodness-of-fit tests were performed using the “Nmix.gof.test” function in “AICmodavg” to evaluate the fit of the data and estimate c-hat.
Results
Vegetation Response
Over five-years, 50 plant species were identified within enhancement channels (n = 10 plots), while 35 species were found in reference channels (n = 9 plots). Both channel types shared 48% of the species pool, including those with the highest importance values. In enhancement channels, Spirodela polyrhiza (36%), Ceratophyllum demersum (31%) and Hydrocharis morsus-ranae (19%) had highest importance values (Table 1). Similarly, Spirodela polyrhiza (48%) and Hydrocharis morsus-ranae (34%) were top ranked by importance values at reference channels, while Wolffia sp. (22%) ranked third (Table 1). For channel vegetation, 38% of the total species pool was found exclusively in enhancement channels and included Elodea canadensis (16.2%) and Zizania aquatica (4.9%). In contrast, 14% of the species pool was found exclusively in reference channels highlighted by non-native Typha x glauca (8.9%; Table 1).
Structural categories revealed prominence of floating-leaved vegetation at reference channels from 2010 onward, while fine- and broad-leaved SAV were favored at enhancement channels (Fig. 2). With SAV categories combined, we found a significant interaction between channel types and year (LMM: F4, 59 = 3.854, p = 0.007), because percent cover of SAV increased at enhancement channels, but remained relatively stable at reference sites (Fig. 3). Consequently, SAV percent cover was significantly higher in enhancement than reference channels in 2011 (p = 0.01), 2012 (p = 0.005) and 2013 (p = 0.02; Fig. 3), and when pooled among 5-years percent cover of SAV (mean ± SE) was significantly higher at enhancement channels (52.2 ± 7.6%) than reference channels (19.8 ± 8.0%; p = 0.009; Fig. 3).

Relative percent cover of aquatic vegetation for seven structural categories: emergent, floating-leaved, broad-leaved submersed, fine-leaved submersed, algae, and other in enhanced (E) channel plots (n = 10) and reference (R) channel plots (n = 9) 2009 to 2013

Mean percent cover of submerged aquatic vegetation (SAV) at mid channel between enhancement (white bars) and reference sites (grey bars) with box and whisker plots. Mean (x) and median (line) within the boxes, each box represents upper 75th and lower 25th percentile. Bars represent minimum and maximum values, and circles denote outliers. Statistically significant differences between channel type each year denoted with asterisks based on mixed-effect nested ANOVAs, (df = 1,17, α = 0.05)
We found 134 species of plants growing in riparian areas (edge and floodplain) of enhancement channels (n = 32 plots) compared with 60 species at reference channels (n = 36 plots). Importance values indicated that T. x glauca was the top ranked taxon at the enhancement (33%) and reference channels (70%), while rank order of other taxa differed between channel types (Table 1). Carex lacustris (11.1%) and invasive species, Acorus calamus (5.5%) and Phragmites australis (2.8%) were only found in riparian enhancement channel plots.
In response to excavation, enhancement channels experienced increased FQI scores and percent cover of short-emergent vegetation and appeared to coincide with declines of robust-emergent vegetation at edge and floodplain habitats (Fig. 4). Significant within-year differences occurred between channel types (p ≤ 0.05) after 2011 for FQI scores and percent cover of short-emergent vegetation (Fig. 4a, b), while differences in cover of robust-emergent vegetation was pronounced in 2009 (Fig. 4c). Although treatment-by-year interactions were weak or not apparent when edge and floodplain habitats were examined independently, within year differences between channel types were similar (Fig. 4). Thus, when data were pooled among years, FQI scores were significantly higher at enhancement than reference sites for both floodplain (LMM: F1, 17 = 11.28, p = 0.004) and edge habitats (F1, 17 = 4.34, p = 0.05; Fig. 4a). Percent cover of short-emergent vegetation was also significantly higher at enhancement than reference channels for floodplain habitat (F1, 17 = 5.85, p = 0.03) but marginal for edge habitat (F1, 17 = 3.60, p = 0.08; Fig. 4b). In contrast, percent cover of robust emergent vegetation was lower at enhancement channels at floodplain (F1, 17 = 8.17, p = 0.01) and edge habitats (F1, 17 = 8.32, p = 0.01; Fig. 4c).

Box-plots of floodplain riparian (left panels) and edge (right panels) habitats between enhanced (grey) and reference channels (white) among years for (a) Floristic Quality Index, (b) percent cover of short-emergent (SE) vegetation, and (c) percent cover of robust-emergent (RE) vegetation. Mean (x) and median (line) indicated, each box represents upper 75th and lower 25th percentile. Bars represent minimum and maximum values, and circles denote outliers. Statistically significant differences between channel type each year denoted with asterisks based on mixed-effect nested ANOVAs (df = 1,17, α = 0.05)
With pooled edge and floodplain plots that represented the riparian zone, differences between the excavated and reference channels were pronounced, as treatment effects were found for each habitat metric pooled among years (F1, 17 > 5.90, p ≤ 0.03; Fig. 5). Treatment-by-year interactions for FQI scores and percent cover of robust emergent vegetation were also more robust (LMM: F4, 68 > 2.90, p < 0.03), indicating increased ecological condition (i.e., increased diversity and community composition of native species) through time (Fig. 5a and c respectively). When annual data were examined, both FQI scores and cover of short-emergent vegetation tended to be higher at enhancement sites than reference channels for most years (p ≤ 0.10; Fig. 5a, b), while percent cover of robust-emergent vegetation was significantly lower at enhancement channels in 2009 (p = 0.01) and 2013 (p = 0.02; Fig. 5c).

Box and whisker plots of (a) Floristic Quality Index scores, (b) percent cover of short-emergent vegetation, and (c) percent cover of robust-emergent vegetation at the reference (white) and enhancement (grey) channels among years that were pooled between the edge and riparian habitat zones. Mean (x) and median (line) within the boxes, each box represents upper 75th and lower 25th percentile. Bars represent minimum and maximum values, and circles denote outliers. Differences between channel type each year were evaluated with mixed-effect nested ANOVAs; asterisks denote statistically significant differences (α ≤ 0.05), while † indicates differences at α ≤ 0.10 and α > 0.05
When data were pooled among years, maximum height of Typha was lower at enhancement (2.19 m ± 0.04) relative to reference sites (2.54 m ± 0.03) (LMM: F1, 17 = 18.42, p < 0.001). Generally, Typha height at enhancement channels tended to increase with time since excavation, but with the exception of 2012, Typha maximum height was significantly lower at enhancement sites each year (p < 0.02). Mean Typha stem density, however, did not differ statistically between enhancement (21.6 ± 1.0 stems/m2) and reference sites (25.1 ± 0.9 stems/m2) when data were pooled among years (LMM: F1, 17 = 2.20, p = 0.16).
In support of observed patterns, the NMS model revealed two distinct vegetation assemblages for the reference and enhancement channels (Fig. 6). Assemblages of riparian vegetation differed significantly between channel types in 2009 (ANOSIM Global R = 0.304), 2010 (ANOSIM Global R = 0.164), 2011 (ANOSIM Global R = 0.21), and 2013 (ANOSIM Global R = 0.25; Fig. 6).

Two-dimensional NMS model for plant assemblages from riparian plots at enhancement channels (open circles; n = 22) and reference channels (closed circles, n = 24) from 2009 through 2013. All ordination plots had a 2-D stress value ≤0.2. ANOSIM R values are displayed
Northern Pike Response to Restored Connectivity
Between 2011 and 2013, a total of 304 YOY northern pike were captured emigrating from enhancement channels, while 74 were caught at reference channels. Daily CPUE of YOY northern pike, pooled among years, was significantly higher at enhancement (mean ± SD: 0.63 ± 0.07) than reference channels (0.37 ± 0.08) (LMM: F 1, 379 = 4.197, p = 0.041; Fig. 7). Catches of YOY northern pike also had a significant year effect (LMM: F 2, 388 = 8.754, p < 0.001) where CPUE was significantly lower in 2012 than in 2011 and 2013, such that catches of northern pike from excavated channels in 2011 and 2013 were significantly higher than both channel types in 2012 (Fig. 7). Total catch of northern pike was comparably high in 2011 (n = 180) and 2013 (n = 183), but approximately 12 times lower in 2012 (n = 15). Moreover, only one YOY northern pike was caught from reference channels in 2012. The low numbers of emigrating YOY northern pike in 2012 did not appear influenced by yearly variation in overall fish abundances caught by nets. Total fish abundance was lowest in 2011 (n = 2546) and comparably high catches were observed in 2012 (n = 4145) and 2013 (n = 4479), suggesting an alternative cause for the low catches of YOY northern pike in 2012.

Mean Daily CPUE of YOY northern pike (NP) between channel type, among years. CPUE was significantly higher at enhancement than reference channels with pooled data among years (LMM: F 1, 379 = 4.197, p = 0.041). A significant year effect was found where abundance of YOY NP was significantly lower in 2012 than 2011 and 2013 (LMM: F 2, 388 = 8.754, p < 0.001). Bars not sharing the same letter denote significant channel type-by-year differences (Tukey; α ≤ 0.05)
Environmental Comparison between Channel Types
Mean daily temperatures were significantly higher at enhancement than reference sites each year (p < 0.001; Fig. 8). As a result, MDT were warmer at enhancement (mean ± SE: 20.3 ± 0.26 °C) than reference channels (18.4 ± 0.28 °C) when pooled among years (LMM: F1, 245 = 179.00, p < 0.001). By channel type, we also found a year effect (LMM: F2, 69 = 3.23, p = 0.04) where MDT in 2012 (mean ± SE: 20.1 ± 0.49 °C) was higher than in 2013 (18.5 ± 0.43 °C; p = 0.04), while MDT in 2011 (19.5 ± 0.42 °C) did not differ from the other years. Mean [DO] was also higher at enhancement than reference channels each year (p < 0.01; Fig. 8) and higher at enhancement (mean ± SE: 4.13 ± 0.16 mg/L) relative to reference channels (2.31 ± 0.20 mg/L) when pooled among years (LMM: F1, 285 = 81.23, p < 0.001). We also found a year effect (LMM: F2, 73 = 14.97, p < 0.001), where [DO] was comparably low in 2011 (2.34 ± 0.26 mg/L) and 2013 (2.91 ± 0.25 mg/L), but significantly higher in 2012 (4.43 ± 0.29 mg/L; p < 0.001).

Mean daily temperatures (MDT) from enhanced and reference channels each survey period (a) and (b) mean daily dissolved oxygen concentrations. Asterisks indicate significant differences between channel type within year (p < 0.05)
Water Level Effect on YOY Northern Pike Emigration
Qualitatively, catches of YOY northern pike appeared related to hydrologic conditions during spring spawning and emigration periods (1- April to 1-July). Between 2011 and 2013, mean water levels were highest in 2011 (mean ± SD: 74.97 ± 0.21 mIGLD85), followed by 2013 (74.77 ± 0.16 mIGLD85) and lowest in 2012 (74.76 ± 0.03 mIGLD85) (Fig. 9). When the timing of peak water level was examined, those from 2011 and 2013 occurred in June (75.26 and 75.02 mIGLD85, respectively) and corresponded with northern pike emigration. In contrast, peak water level in 2012 occurred in mid-April (74.85 mIGLD85), when spawning northern pike would access upslope habitats, but levels declined by ~ 0.10 m when YOY were expected to emigrate (Fig. 9).

Mean daily water level (mIGLD85) during northern pike spawning and emigration (1-April to 1-July) from 2011 to 2017. Data downloaded from Alexandria Bay, NY (NOAA buoy station ID: 8311062, https://tidesandcurrents.noaa.gov/waterlevels.html?id=8311062)
Between 2011 and 2016, mean water levels had a range of 0.29 m (Fig. 9). Mean water levels (±SD) in 2017 (75.46 ± 0.21 mIGLD85), however, exceeded the second highest water year by 0.45 m (i.e., 2011) and was 0.63 m higher than the average water level of the preceding 6 years, indicative of an extreme high water event (Fig. 9). Corresponding with high water levels in 2017, when site specific CPUE of out-migrating YOY northern pike was ranked among years (n = 63), nine of the 10 highest CPUEs recorded occurred in 2017, resulting in substantially higher catches that year (Fig. 10).

Yearly comparison of mean northern pike (NP) CPUE (± SE) between excavated spawning habitats and reference channels. Data from 2014 was excluded because it consisted of a single data point
Hierarchal modelling further suggested that water levels were a primary factor in the high catches of northern pike in 2017. Although none of the models had a strong fit to the data based on goodness-of-fit testing and c-hat comparisons (Table 2), the top ranked model from AICc model selection was the additive model of water levels and effort (observational-level covariates) followed by the additive model with the site-level covariate (Table 2). Both observational-level covariates in the top ranked model had positive coefficients indicating that higher water levels and greater effort improved the detectability of out-migrating northern pike; however, AICc model-average estimates (±95% CI) suggested that water levels (1.52 ± 0.06) had a stronger influence on detection probability than effort (0.91 ± 0.05).
Discussion
In a Typha-dominated coastal wetland, channel excavation restored connectivity to isolated wet meadow zones, increased channel riparian vegetation and created functioning northern pike reproductive habitat. After the dominant Typha was reduced and the spoil side-casted, the excavated channels were rapidly colonized by native vegetation. This outcome is most likely a result of a newly exposed and viable seedbank (van der Valk 1986; Hulme and Bremner 2006; Vosse et al. 2008; O’Donnell et al. 2014) that was released from competition by Typha (Farrar and Goldberg 2009). This mechanism is supported by Farrell et al. (2010) in a study of transplanted elevation-composited sediments from nearby Typha dominated St. Lawrence River wetlands into a greenhouse under two sets of moisture conditions (mesic and saturated). A diversity of seed bank germinated, with the mesic treatment producing the greatest plant species diversity.
In addition to physical disturbance caused by excavation (e.g. spoil), renewed hydrologic connectivity to sedge meadow habitat likely provided a dispersal pathway that may have promoted colonization of native vegetation (Nilsson et al. 1991; Seabloom and van der Valk 2003; Nilsson et al. 2010). In two years, a unique and more diverse community of vegetation (as assessed by FQI) was found at excavated channels relative to reference sites that appeared representative of conditions expected under natural hydrologic conditions (Wilcox and Meeker 1991; Farrell et al. 2010). The timeframe that vegetation responded to channel excavation was similar to that reported by others (i.e., 2–4 years; Keddy and Reznicek 1986; Wilcox and Nichols 2008) who examined the response of wetland vegetation under different hydrologic disturbance regimes. Therefore, the disturbance caused by channel excavation (both exposed sediments and reduced competitive dominance of Typha) promoted a diversity of native plants to germinate (i.e., as assessed by the FQI) despite the lack of complete natural hydrologic disturbance due to water level regulation under Plan 1958D (Keddy and Reznicek 1986; Wilcox 2004). It was also apparent that improvements in habitat quality at enhancement channels, such as declines in robust-emergent vegetation, and corresponding increases in cover of SAV, short-emergent vegetation and FQI scores were sustained until the end of the study. It is still likely that over time invasive Typha will re-encroach upon the newly exposed riparian channel habitats due to its dominance in adjacent areas unaffected by excavation (Wilcox et al. 2018) and known mechanisms of spread (Larkin et al. 2012).
A primary goal of channel excavation was to remove fish passage barriers and create hydrologic links to enhance connectivity to isolated wet meadow habitats (i.e., preferred spawning habitat of northern pike; Casselman and Lewis 1996; Farrell 2001; Cooper et al. 2008) to promote reproduction. Not only did northern pike find and exploit newly accessible spawning habitat, but habitat and environmental conditions in the excavated channels appeared more favorable for early-life survival than unaltered reference sites. Although spawning and larval fish habitat relationships were indirectly assessed, the higher percent cover of broad- and fine-leaved SAV in the enhancement channels relative to reference sites suggested higher quality fish habitat (Casselman and Lewis 1996; Randall et al. 1996; Nilsson et al. 2014). Water temperatures and [DO] were also significantly higher at the enhancement channels and these two variables are often associated with increased survivorship and growth for northern pike (Inskip 1982; Casselman and Lewis 1996; Farrell 2001; Farrell et al. 2006). Massa and Farrell (accepted pending revision) similarly report higher temperature and [DO] for excavated sites in 2016. The lower temperatures and [DO] at the reference channels are suspected to be influenced by ground water seepage that maintained these remnant channels in the face of Typha expansion. Although it is possible that thermal conditions at the reference channels were influenced by shading effects of taller Typha and higher cover of floating leaved vegetation than excavated channels, we feel that this shading effect was not sufficient to account for the observed differences in temperatures between the treatments.
Consistently higher catches of YOY northern pike at enhancement channels appeared related to accessibility to preferred spawning habitat, and higher habitat and environmental quality (Casselman and Lewis 1996; Nilsson et al. 2014). Excavated channels appeared to function just as well or better as northern pike early-life habitats than reference sites as evident by significantly higher YOY CPUE at enhancement channels between 2011 and 2013 (Holland and Huston 1984; Farrell et al. 2006; Nilsson et al. 2014; Oele et al. 2015). Massa and Farrell (accepted pending revision), used marked and released pike larvae to evaluate relative survival between excavation and reference sites in French Creek in 2016 and evaluated forage allocation through a study of zooplanktivory. Consistent with our findings on northern pike CPUE, channel and pool excavations demonstrated greater survival rates to emigration relative to reference locations. Differences in prey density and selection relative to larval pike diets among channel types showed some interesting dynamics, but did not suggest major prey limitation or reflect any explanatory effect on survival rates (Massa and Farrell accepted pending revision).
Johnson (1957) purported a strong relationship between water levels and northern pike production, while Nilsson et al. (2014) found strong northern pike reproduction when shallow vegetated areas were re-flooded. Our data, between 2011 and 2013, also indicates hydrologic conditions during the spawning and emigration period influenced northern pike reproduction. Despite consistently catching more YOY northern pike at enhancement channels each year, total abundance was more than twelve-fold lower in 2012 than 2011 and 2013. Water levels in 2012 were among the highest recorded at the beginning of the spawning period (i.e., peaking by mid-April), but unlike other years, water levels declined throughout the remainder of the season. We suspect in 2012 northern pike gained access to inundated spawning habitats in early to mid-April, but the subsequent drawdown reduced hydrologic connectivity between the spawning habitat and emigration routes to French Creek and increased early-life mortality (Johnson 1957; Inskip 1982; Mingelbier et al. 2008; Nilsson et al. 2014).
Water temperature and [DO] measured in 2012 were similar or higher than in 2011 and 2013. At both the reference and excavated channels in 2012, temperature was within the range considered optimal for the growth of juvenile northern pike (i.e., 19–21 °C; Casselman 1978), while [DO] was the highest measured among years. If temperature and [DO] were major contributors to catches of northern pike, then CPUE in 2012 would expectantly had been higher, but was significantly lower than catches at excavated channels in 2011 and 2013.
The relationships between pike, water level and connectivity within habitat serve to highlight the importance of flow regime as a master variable in river restoration (Poff et al. 1997; Richter et al. 2003). The influence of hydrologic conditions on natural reproduction of northern pike was most apparent when combined site data were examined over seven years. Although, hierarchal modelling indicated that water levels had a stronger influence of detecting juvenile northern pike than effort, both variables showed strong positive relationships. The high water levels in early spring of 2017 provided greater access to preferred northern pike spawning habitat, while the rising levels during outmigration would have also maintained connectivity of emigration routes that would expectantly promote survival from both habitat types (Farrell 2001; Smith et al. 2007; Mingelbier et al. 2008; Foubert et al. 2019). Caution should also be applied, however, when extrapolating patterns from modeling results as the modelling fit was weak and included an extreme hydrologic event. Additional years of data that encompass a wider range in water levels, and inclusion of variables related to natural reproduction and early-life survivorship of northern pike (e.g., temperature, [DO]; Smith et al. 2007) may improve predictive value.
We anticipated that higher early spring levels would provide greater accessibility for northern pike to critical early-life habitats (Farrell 2001; Cooper et al. 2008), while prolonged low levels are necessary to alleviate the spread and reduce current densities of T. x glauca (Keddy and Reznicek 1986; Wilcox et al. 2008). The new regulation plan 2014 is designed to allow greater variability in an attempt to benefit wetland processes (IJC 2014). T. x glauca is competitively superior to Carex spp. when water levels fluctuations are suppressed and stable (Wilcox et al. 2008). Moderate high water levels can also facilitate T. x glauca (Farrell et al. 2010) and extreme high water levels (>1 m over plants) in the growing season can create a die-off (Wilcox 2004; Wei and Chow-Fraser 2006) but those extremes are beyond plan constraints. Low water levels and droughts favor graminoids and sedge species (Farrell et al. 2010; Wilcox 2004) and would allow species suppressed by Typha (e.g., Carex spp.) an opportunity to recolonize disturbed habitat following a flood (Keddy and Reznicek 1986; Wilcox 2004; Farrell et al. 2010) but changes to seedbanks may prevent such resilience in more modified wetland complexes (Frieswyk and Zedler 2006). Concern remains that connectivity enhancements and natural linkages will be blocked in the future due to ongoing Typha encroachment and through the effect of dislodged portions of floating mats clogging channels (Farrell et al. 2010; Wilcox and Bateman 2018). Monitoring these processes in the context of regulation and climatic variation will be important in determining the long-term viability of connectivity restoration projects.
References
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67:1–48
Boers AM, Zedler JB (2008) Stabilized water levels and Typha invasiveness. Wetlands 28:676–685
Boers AM, Veltman RLD, Zedler JB (2007) Typha x glauca dominance and extended hydroperiod constrain restoration of wetland diversity. Ecological Engineering 29:232–244
Bourdaghs M, Johnston CA, Regal RR (2006) Properties and performance of the floristic quality index in Great Lakes coastal wetlands. Wetlands 26:718–735
Bried JT, Strout KL, Portante T (2012) Coefficients of conservatism for the vascular flora of New York and New England: inter-state comparisons and expert opinion bias. Northeastern Naturalist 19:101–114
Bry C (1996) Role of vegetation in the life cycle of pike. In: Craig JF (ed) Pike: biology and exploitation. Chapman and Hall, London, pp 45–67
Casselman JM (1978) Effects of environmental factors on growth, survival, activity and exploitation of northern pike. In: Kendall RL (ed), selected coolwater fishes of North America. American Fisheries Society,Special Publication 11, p 114–128
Casselman JM, Lewis CA (1996) Habitat requirements of northern pike (Esox lucius). Canadian Journal of Fisheries and Aquatic Sciences 53:161–174
Clarke KR (1993) Non-parametric multivariate analyses of changes in community structure. Australian Journal of Ecology 18:117–143
Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial. PRIMER-E ltd. Plymouth (190 pp.)
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation, second edn. Natural Environmental Research Council, Plymouth Marine Laboratory, Plymouth, United Kingdom
Cooper JE, Mead JV, Farrell JM, Werner RG (2008) Potential effects of spawning habitat changes on the segregation of northern pike (Esox lucius) and muskellunge (E. masquinongy) in the upper St. Lawrence River. Hydrobiologia 601:41–53
Dionne M, Short F, Burdick D (1999) Fish utilization of enhanced, created, and reference salt-marsh habitat in the Gulf of Maine. American Fisheries Society Symposium 22:384–404
Farrell JM (2001) Reproductive success of sympatric northern pike and muskellunge in an upper St. Lawrence River Bay. Transactions of the American Fisheries Society 130:796–808
Farrell JM, Mead JV, Murry BA (2006) Protracted spawning of St Lawrence river northern pike (Esox lucius): simulated effects on survival, growth, and production. Ecology of Freshwater Fish 15:169–179
Farrell JM, Murry BA, Leopold DJ, Rippke MB, Rippke MB, Godwin KS, Hafner SD (2010) Water-level regulation and coastal wetland vegetation in the upper St. Lawrence River: inferences from historical aerial imagery, seed banks, and Typha dynamics. Hydrobiologia 647:127–144
Farrer EC, Goldberg DE (2009) Litter drives ecosystem and plant community changes in cattail invasion. Ecological Applications 19:398–412
Freeman MC, Bowen ZH, Bovee KD, Irwin ER (2001) Flow and habitat effects on juvenile fish abundance in natural and altered flow regimes. Ecological Applications 11:179–190
Foubert A, Le Pichon C, Mingelbier M, Farrell JM, Morin J, Lecomte F (2019) Modeling the effective spawning and nursery habitats of northern pike within a large spatio-temporally variable river landscape (St. Lawrence River, Canada). Limnology and Oceanography 64:803–819
Frieswyk CB, Zedler JB (2006) Do seed banks confer resilience to coastal wetlands invaded by Typha×glauca? Canadian Journal of Botany 84:1882–1893
Galatowitsch SM, Anderson NO, Ascher PD (1999) Invasiveness in wetland plants in temperate North America. Wetlands 19:733–755
Havens KJ, Varnell LM, Watts BD (2002) Maturation of a constructed tidal marsh relative to two natural reference tidal marshes over 12 years. Ecological Engineering 18:305–315
Herman KD, Masters LA, Penskar MR, Reznicek AA, Wilhelm GS, Brodovic WW, Gardiner KP (2001) Floristic quality assessment with wetland categories and examples of computer applications for the state of Michigan. Michigan Dept. of natural resources, wildlife division, natural heritage program, [Lansing, MI]
Holland LE, Huston ML (1984) Relationship of young-of-the-year northern pike to aquatic vegetation types in backwaters of the upper Mississippi River. North American Journal of Fisheries Management 4:514–522
Hulme PE, Bremner ET (2006) Assessing the impact of Impatiens glandulifera on riparian habitats: partitioning diversity components following species removal. Journal of Applied Ecology 43:43–50
IJC (2014) International Joint Commission. Lake Ontario St. Lawrence River Plan 2014: Protecting against extreme water levels, restoring wetlands and preparing for climate change
Inskip PD (1982) Habitat suitability index models: northern pike. No. FSW/OBS-82/10.17. Fish and wildlife service Fort Collins co. Western energy and land use team
Johnson FH (1957) Northern pike year-class strength and spring water levels. Transactions of the American Fisheries Society 86:285–293
Jude DJ, Pappas J (1992) Fish utilization of Great Lakes coastal wetlands. Journal of Great Lakes Research 18:651–672
Keddy PA, Reznicek AA (1986) Great Lakes vegetation dynamics: the role of fluctuating water levels and buried seed. Journal of Great Lakes Research 12(1):25–36
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package: tests in linear mixed effects models. Journal of Statistical Software 82:1–26
Landress CM (2016) Fish assemblage associations with floodplain connectivity following restoration to benefit an endangered catostomid. Transactions of the American Fisheries Society 145:83–93
Larkin DJ, Madon SP, West JM, Zedler JB (2008) Topographic heterogeneity influences fish use of an experimentally-restored tidal marsh. Ecological Applications 18:483–496
Larkin DJ, Freyman MJ, Lishawa SC, Geddes P, Tuchman NC (2012) Mechanisms of dominance by the invasive hybrid cattail Typha x glauca. Biological Invasions 14:65–77
Lishawa SC, Lawrence BA, Albert DA, Tuchman NC (2015) Biomass harvest of invasive Typha promotes plant diversity in a Great Lakes coastal wetland. Restoration Ecology 23:228–237
Massa EA, Farrell JM (accepted pending revision) Improving habitat connectivity in a Typha dominated wetland shows increased larval northern pike survival. Wetlands
Mazerolle MJ (2017) AICcmodavg: model selection and multimodel inference based on (Q)AIC(c). R package version 2.1–1. https://cran.r-project.org/package=AICcmodavg. Viewed August 12, 2018
Minello T, Webb JJ (1997) Use of natural and created Spartina alterniflora salt marshes by fishery species and other aquatic fauna in Galveston Bay, Texas, USA. Marine Ecology Progress Series 151:165–179
Mingelbier M, Brodeur P, Morin J (2008) Spatially explicit model predicting the spawning habitat and early stage mortality of northern pike (Esox lucius) in a large system: the St. Lawrence River between 1960 and 2000. Hydrobiologia 601:55–69
Nilsson C, Garfjell M, Grelsson G (1991) Importance of hydrochory in structuring plant communities along rivers. Canadian Journal of Botany 69:2631–2633
Nilsson C, Brown RL, Jansson R, Merritt DM (2010) The role of hydrochory in structuring riparian and wetland vegetation. Biological Reviews 85:837–858
Nilsson J, Engstedt O, Larsson P (2014) Wetlands for northern pike (Esox lucius L.) recruitment in the Baltic Sea. Hydrobiologia 721:145–154
O’Donnell J, Fryirs K, Leishman MR (2014) Digging deep for diversity: riparian seed bank abundance and species richness in relation to burial depth. Freshwater Biology 59:100–113
Oele DL, Hogan JD, McIntyre PB (2015) Chemical tracking of northern pike migrations: if we restore access to breeding habitat, will they come? Journal of Great Lakes Research 41:853–861
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BR, Sparks RE, Stromberg JC (1997) The natural flow regime. BioScience 47:769–784
Randall RG, Minns CK, Cairns VW, Moore JE (1996) The relationship between an index of fish production and submerged macrophytes and other habitat features at three littoral areas in the Great Lakes. Canadian Journal of Fisheries and Aquatic Sciences 53:35–44
Richter BD, Mathews R, Harrison DL, Wigington R (2003) Ecologically sustainable water management: managing river flows for ecological integrity. Ecological Applications 13:206–224
Rippke MB, Distler MT, Farrell JM (2010) Holocene vegetation dynamics of an upper St. Lawrence River wetland: paleoecological evidence for a recent increase in cattail (Typha). Wetlands 30:805–816
Schummer ML, Palframan J, McNaughton E, Barney T, Petrie SA (2012) Comparisons of bird, aquatic macroinvertebrate, and plant communities among dredged ponds and natural wetland habitats at long point, Lake Erie, Ontario. Wetlands 32:945–953
Seabloom EW, Van der Valk AG (2003) Plant diversity, composition, and invasion of restored and natural prairie pothole wetlands: implications for restoration. Wetlands 23:1–12
Smith SG (1967) Experimental and natural hybrids in north American Typha (Typhaceae). The American Midland Naturalist 78:257–287
Smith BM, Farrell JM, Underwood HB, Smith SJ (2007) Year-class formation of upper St. Lawrence River northern pike. North American Journal of Fisheries Management 27:481–491
Swink F, Wilhelm G (1994) Plants of the Chicago region. Indiana Academy of Science Indianapolis
Taft JB, Wilhelm GS, Ladd DM, Masters LA (1997) Floristic quality assessment for vegetation in Illinois, a method for assessing vegetation integrity. Illinois Native Plant Society
Vaccaro LE, Bedford BL, Johnston CA (2009) Litter accumulation promotes dominance of invasive species of cattails (Typha spp.) in Lake Ontario wetlands. Wetlands 29:1036–1048
van der Valk AG (1986) The impact of litter and annual plants on recruitment from the seed bank of a lacustrine wetland. Aquatic Botany 24:13–26
Vosse S, Esler KJ, Richardson DM, Holmes PM (2008) Can riparian seed banks initiate restoration after alien plant invasion? Evidence from the Western cape, South Africa. South African Journal of Botany 74:432–444
Wei A, Chow-Fraser P (2006) Synergistic impact of water level fluctuation and invasion of Glyceria on Typha in a freshwater marsh of Lake Ontario. Aquatic Botany 84:63–69
Weinstein MP, Litvin SY (2016) Macro-restoration of tidal wetlands: a whole estuary approach. Ecological Restoration 34:27–38
Wilcox DA (2004) Implications of hydrologic variability on the success of plants in Great Lakes wetlands. Aquatic Ecosystem Health and Management 7:223–231
Wilcox DA, Bateman JA (2018) Photointerpretation analysis of plant communities in Lake Ontario wetlands following 65 years of lake-level regulation. Journal of Great Lakes Research 44:1306–1313
Wilcox DA, Meeker JE (1991) Disturbance effects on aquatic vegetation in regulated and unregulated lakes in northern Minnesota. Canadian Journal of Botany 69:1542–1551
Wilcox DA, Nichols SJ (2008) The effects of water-level fluctuations on vegetation in a Lake Huron wetland. Wetlands 28:487–501
Wilcox DA, Kowalski KP, Hoare HL, Carlson ML, Morgan HN (2008) Cattail invasion of sedge/grass meadows in Lake Ontario: photointerpretation analysis of sixteen wetlands over five decades. Journal of Great Lakes Research 34:301–323
Wilcox DA, Buckler K, Czayka A (2018) Controlling cattail invasion in sedge / grass meadows. Wetlands 38:337–347
Wilhelm G (1977) Ecological assessment of open land areas in Kane County, Illinois: a checklist of the Kane County flora with numerical evaluations--its basis, rationale and application. Kane County Urban Development
Williams GD, Zedler JB (1999) Fish assemblage composition in constructed and natural tidal marshes of San Diego Bay: relative influence of channel morphology and restoration history. Estuaries and Coasts 22:702–716
Acknowledgements
This project was funded through the Fish, Enhancement, Mitigation, and Research Fund administered by the US Fish and Wildlife Service, and the NOAA Great Lakes Restoration Initiative through Ducks Unlimited. Restoration projects were completed by Partners for Fish and Wildlife at the Cortland, NY Field Office and Ducks Unlimited. We thank Scott Schlueter of the US Fish and Wildlife Service and Sarah Fleming, NOAA PI, Ducks Unlimited and the NYS Department of Environmental Conservation, especially Irene Mazzocchi, Rodger Klindt and Steve LaPan for logistical and collaborative support. We also thank Thousand Islands Biological Station staff, students, and technicians, with special mention to J. Runner, E. Polzer, M. Distler, C. Barry, B. Henning, and M. Regan. This research article is a contribution of the Thousand Islands Biological Station.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Neveldine, B.L., Leblanc, J.P. & Farrell, J.M. Vegetation Response and Juvenile Northern Pike (Esox lucius) Outmigration Following Connectivity Enhancement of a Typha Dominated Coastal Wetland. Wetlands 39, 921–934 (2019). https://doi.org/10.1007/s13157-019-01145-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-019-01145-y