
Abstract
Water oxidation in photosystem II (PSII) is performed by the oxygen-evolving complex Mn4CaO5 which can be extracted from PSII and then reconstructed using exogenous cations Mn(II) and Ca2+. The binding efficiency of other cations to the Mn-binding sites in Mn-depleted PSII was investigated without any positive results. At the same time, a study of the Fe cations interaction with Mn-binding sites showed that it binds at a level comparable with the binding of Mn cations. Binding of Fe(II) cations first requires its light-dependent oxidation. In general, the interaction of Fe(II) with Mn-depleted PSII has a number of features similar to the two-quantum model of photoactivation of the complex with the release of oxygen. Interestingly, incubation of Ca-depleted PSII with Fe(II) cations under certain conditions is accompanied by the formation of a chimeric cluster Mn/Fe in the oxygen-evolving complex. PSII with the cluster 2Mn2Fe was found to be capable of water oxidation, but only to the H2O2 intermediate. However, the cluster 3Mn1Fe can oxidize water to O2 with an efficiency about 25% of the original in the absence of extrinsic proteins PsbQ and PsbP. In the presence of these proteins, the efficiency of O2 evolution can reach 80% of the original when adding exogenous Ca2+. In this review, we summarized information on the formation of chimeric Mn–Fe clusters in the oxygen-evolving complex. The data cited may be useful for detailing the mechanism of water oxidation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Oxygen-producing organisms (which include the higher plants, algae, and cyanobacteria) carry out photosynthesis, the most important light-dependent biological process that is the basis of life on Earth. The main components of the photosynthetic biochemical apparatus associated with the light-dependent part of photosynthesis are photosystem II (PSII) and photosystem I (PSI). PSII contains a unique catalytic center facilitating the oxidation of water molecules which are the donors of protons and electrons required for NADP+ reduction and ATP synthesis with the resulting molecular oxygen released into the atmosphere as a by-product. This reaction is essentially the only source of O2 on our planet. The catalytic center of water oxidation consists of four manganese cations and one calcium cation interconnected by oxygen bridges—Mn4CaO5. The structural organization of the manganese/calcium cluster has been solved by X-ray diffraction at 1.9 Å resolution (Umena et al. 2011). The atomic structure of the oxygen-evolving complex (OEC) within PSII is shown in Fig. 1. It is an irregular cube formed by three manganese cations, a calcium cation and four oxygen atoms. The fourth manganese cation (Mn4 according to the numbering scheme adopted by Umena et al. (2011), also called the “dangler” cation) is located at some distance from the cube but is connected to it. This “dangler” manganese cation is connected to the Mn3Ca cube by two oxygen bridges O4 and O5. The Mn4 cation and the calcium cation each have two water molecules as ligands. Whether these water molecules are substrate water remains to be seen. The catalytic center is connected to the protein structure PSII by amino acid residues. The cation Mn4 is bound to the high-affinity Mn-binding site (HAS). A negative criticism of the work of Umena et al. (2011) was raised in regard to the possibility of X-ray-induced reduction of manganese cations during the experiment, which could affect the accuracy of distance determination between cluster elements. This possible artifact was later eliminated using serial femtosecond X-ray free-electron laser crystallography (Suga et al. 2015) for investigation of the Mn/Ca cluster structure in the S1 state. In subsequent works, this method was used to study the structural changes occurring in the cluster during the transition from S1 state to S2 and S3 states (Suga et al. 2017, 2019; Kern et al. 2018). Another method of structural investigation, cryo-electron microscopy, was recently used to study monomeric PSII from Synechocystis sp. PCC 6803 lacking the OEC (Gisriel et al. 2020). The authors found significant differences in the structure of the Mn-binding HAS obtained by the cryo-electron microscopy and X-ray diffraction methods earlier (Zhang et al. 2017). Quite recently, a number of advances have been made in the study of the OEC structure, with regard to changes in the catalytic cycle process and understanding the details of the water oxidation mechanism and molecular oxygen synthesis (Shen 2015; Lubitz et al. 2019; Oliver et al. 2022; Shevela et al. 2023). The most discussed hypotheses of which oxygen atoms are involved in the synthesis of O2, the oxygen bridge O5 is the most likely candidate. However, the nature of the second oxygen atom is less clear. It could be a newly inserted water molecule Wx (Shen 2015) or Wf (Lubitz et al. 2019) or water molecule W3 bound to Ca2+ ion (Shen 2015).

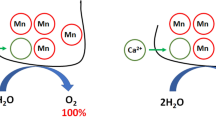
A Photosynthetic apparatus and electron transport chain in plants. Four protein macrocomplexes participate in the production of ATP and NADPH: photosystems PSII and PSI, the cytochrome b6/f complex, and the ATP synthase. PSII functions as a water/PQ oxidoreductase, which has a manganese complex [Mn4O5Ca], located on D1 protein on the PSII donor side, as well as pheophytin, plastoquinones QA and QB, and a non-heme iron binding a bicarbonate ion on the PSII acceptor side. The structural data on membrane-protein complexes and soluble proteins were obtained from Protein Data Bank (PDB): PSII–6JLJ, PSI–6JBO, cyt b6/f complex–7QRM, ATP synthase–6VOF, plastocyanin–2CJ3, ferredoxin–4ZHP, ferredoxin-NADP+ reductase–1FNB. B Structure of PSII and catalytic center of oxygen-evolving complex: Ca, Mn(1–4), and O (red circles) are ions of calcium and manganese and oxygen atoms; W(1–4) are oxygen atoms of water molecules acting as ligands of manganese and calcium. PDB ID: 6jlj. Mn1 ligands: D1-Glu189, His332 and Asp342, D1-Asp342; Mn2 ligands: Ala344 and CP43-Glu354; Mn3 ligands: D1-Glu333 and CP43-Glu354; Mn4 (HAS) ligands: D1-Asp170 and D1-Glu333; Ca2+ ligands: D1-Asp170 and D1-Ala344 (Umena et al. 2011). The right panel shows the structures of chimeric Mn-Fe clusters in the oxygen-evolving complex, obtained after removing the calcium cation
When studying the role and mechanism of metal cations in enzymes, the method of replacing a native cation with other metal cations is often used in order to clarify the question—what features of a native metal cation are important for its special function? In the case of PSII, the possibilities of binding and replacing Ca2+ and Mn cations in Mn4CaO5 catalytic center of the OEC with different cations have been extensively previously investigated. The Ca2+ cation can be substituted while retaining oxygen-evolving function only by Sr2+ (Ghanotakis et al. 1984a). Other cations can effectively bind to the Ca site of the OEC but without recovery of OEC activity, with these including Cd2+ cations (Waggoner and Yocum 1990) and various lanthanides (Ghanotakis et al. 1985; Ono 2000; Lovyagina et al. 2021). The efficiency of binding of the metal cation to the Ca site and its ability to restore the functional activity of OEC depends on the ion radius and the value of the aqua ion pKa (Vrettos et al. 2001). The possibility of binding other metal cations to the Mn-binding sites of the PSII has also been investigated, but less extensively. Ghirardi et al. (1996), using the method of flash-probe fluorescence, showed that the cations Zn2+ and Co2+ bind to the Mn-binding site in PSII(-Mn) membranes less efficiently than Mn cations. However, at the same time, it was found that the Mn-binding site(s) bind(s) Fe(II) cations with an efficiency and specificity comparable to the Mn cations (Semin et al. 1995). In subsequent works, this effect was studied in detail and a method was developed for replacing 1 or 2 manganese cations in a catalytic cluster with an Fe cation. In this paper, we provide a brief review of published studies concerned with the Mn cation(s) substitution process in the OEC with iron cations and the effects that have been observed from doing so.
Binding of Fe(II) cations to Mn-binding sites in PSII lacking oxygen-evolving complex
Manganese cations can be extracted from the OEC of PSII by using hydroxylamine or Tris at alkaline pH. Treatment with these compounds results in the reduction of Mn cations and these reduced cations then exit from the binding site. The difference in the effect of these compounds is that Tris extracts all extrinsic proteins, whereas hydroxylamine does not extract PsbO extrinsic protein (Tamura and Cheniae 1987). PSII(-Mn) membranes are capable of oxidizing exogenous electron donors under light and have two oxidation sites located on the donor side. One of these sites is the high-affinity Mn-binding site (HAS); the second site is the low-affinity site (LAS) (Hsu et al. 1987; Kurashov et al. 2009). HAS is the oxidation site for Mn(II) cations, donor pair (Mn(II) + H2O2), and diphenylcarbazide (DPC) (Hsu et al. 1987; Semin et al. 2004; Kurashov et al. 2009), while Mn(II) and DPC are oxidized also at the LAS (Hsu et al. 1987; Kurashov et al. 2009). At the HAS, the oxidant is YZ· (Hoganson et al. 1989), while at the LAS, the His190 residue of the integral protein of the PSII reaction center D1 may participate in oxidation (Kurashov et al. 2009).
Hsu et al. (1987) showed that binding of the Mn(II) cation to the HAS resulted in blocking of the oxidation of the DPC at that site. This is reflected in a 50% decrease in the rate of oxidation of DPC (DPC is oxidized only through the LAS in this case). The use of this test in the study of Fe(II) binding parameters showed that Fe(II) cations bind to a high-affinity Mn-binding site with a high efficiency (Semin et al. 1995) comparable to that of binding manganese cations (Hsu et al. 1987). This is not unique in biological important molecules. There exist a number of enzymes where the catalytic center can contain iron or manganese although the function is the same. To such proteins belong the family of superoxide dismutases (SOD) utilizing either Fe or Mn to catalyze the dismutation of O2·–. In these enzymes, the coordination of ligands at the Fe site of Fe-SOD closely corresponds to the Mn coordination found in Mn-SOD (Lah et al. 1995). Another example would be binuclear iron-containing enzymes such as ribonucleotide reductase in which the iron cofactor can be replaced with manganese (Atta et al. 1992). Additional evidence of highly effective binding of Fe(II) cations to the Mn-binding site are experiments of Allen with coworkers (Kálmán et al. 2010; Tang et al. 2009). Using the EPR method, they showed the binding of Fe(II) cations to an artificially created Mn-binding site in bacterial centers and its oxidation by primary electron donor. Binding of Fe(II) cations requires light, as no effect of Fe(II) cations was observed in the dark. This indicates that only oxidized Fe cations bind to the Mn-binding HAS. However, it should be noted that incubation of the PSII(-Mn) membranes with the Fe(III) is not accompanied by binding of the oxidized iron cation to the HAS and blocking it. The reason for this effect may be the following. Salts of Fe(III) do not dissolve in the buffer with a neutral pH due to the fast formation of insoluble Fe(OH)3. The use of Fe(III) solution stabilized with sucrose (Charley et al. 1963) also did not result in HAS binding and blocking (Semin et al. 1995). However, it has been found relatively recently that such a preparation is a suspension of nanoparticles (Kudasheva et al. 2004), which may be the reason for the lack of binding of iron cations.
These results were confirmed using the method of recording the kinetics of fluorescence decay in response to a flash of light (Semin et al. 2002). In the same work, it was shown that oxidized iron cations, by binding to a HAS with great efficiency, prevent the binding of manganese cations to this site—i.e., this site is blocked by bound Fe(III) cation. Blocking iron cations are bound to the HAS very firmly and cannot be removed by centrifugation. However, they can be extracted with citrate buffer at pH 3.0 (citrate is an efficient chelator of ferric cations having first stability constant (logK1) 11.8 (Sillen and Marten 1964)) (Semin and Seibert 2006a).
Kinetic studies have shown that not one iron cation is involved in blocking, but two or more (Semin et al. 1995, 2002). When measuring the concentration of iron cations in the buffer during blocking, it was found that oxidation of five iron cations was necessary to provide blocking effect (Semin and Seibert 2004). Carboxyl amino acid group, possibly amino acid D1-Asp170, is involved in binding the iron cations blocking the HAS (Semin and Seibert 2006a).
The need for light-dependent charge separation in PSII(-Mn) membranes, accompanied by the generation of oxidant YZ·, and, accordingly, electron transport for oxidation of Fe(II) cations is quite clear from the above facts. However, in the experiment, blocking takes place during the short incubation of the sample (several min) under room light and without the addition of an exogenous electron acceptor. The exogenous acceptor increases the rate of Fe(II) oxidation, and the diuron inhibits this process (Lovyagina et al. 2005). These data suggest that the PSII(-Mn) membrane preparation contains an electron acceptor that ensures the transport of electrons from Fe(II) and that molecular oxygen can be such an acceptor. Indeed, under anaerobic conditions, the rate of Fe(II) oxidation is significantly slowed down (Lovyagina et al. 2005).
The catalytic center of OEC contains in addition to 4 redox-active Mn cations one redox-inactive Ca2+ cation which is essential for the oxygen-evolving activity of the catalytic Mn4CaO5 complex. Without calcium cation, molecular oxygen is not synthesized, indicating its important role in the water oxidation process. The mechanism of Ca2+ participation in this process is unclear. Possibly, it binds the substrate water molecule (W3 or W4 (Kawakami et al. 2011)) or participate in controlling the redox potential of Mn cluster (Tsui et al. 2013; Saito et al. 2021). Em (MnIII/IV) depends on the Lewis acidity of redox-inactive metals (Ca2+ in the OEC) (i.e., the pKa of aqua complexes of redox-inactive metals) (Tsui et al. 2013). Quantum chemical calculations showed that the valence of metals predominantly affects Em (MnIII/IV), whereas the ionic radius of metals affects Em (MnIII/IV) only slightly (Saito et al. 2021). Ca2+ is a necessary component for the reconstruction of Mn/Ca catalytic center during photoactivation (self-assembly of Mn cluster). In this regard, the effect of calcium cations on the binding of iron cations to the Mn-binding sites, in particular, on the reduction of YZ· by the Fe(II) cation in response to a flash of light and on the blocking of HAS by the Fe cation, was studied (Semin et al. 2021). It was shown that the efficiency of Fe(II) oxidation and the efficiency of HAS blocking, depending on the Ca2+ concentration, do not coincide (Semin et al. 2021). The maximum level of inhibition of the Fe(II) oxidation process was 20%, while the inhibition of the blocking process was twice as high at the same Ca2+ concentration. These results indicate that the blocking process is a more complex process than the oxidation of Fe(II) cations at HAS.
The process of iron cations binding to HAS in PSII(-Mn) membranes is also of significant interest because it reduces a number of electron transport chain properties altered by Mn removal. Extraction of Mn from OEC leads to a significant change in the characteristics of the electron transporter YZ such as kinetics and temperature characteristics of YZ oxidation and reduction (see for a review Semin et al. 2005). This may be a consequence of the hydrogen bond break between YZ and D1-His190. The use of the incorporation of iron cations into PSII(-Mn) membranes made it possible to find that the hydrogen bond between these two amino acid residues is not regular, but is a low barrier hydrogen bond (Semin et al. 2005). This bond is broken by extraction of Mn and restored by binding of Fe cation. The data obtained with X-ray diffraction method confirmed our results and conclusions (Saito et al. 2011).
The process of self-assembly of the manganese cluster in apo-PSII is described by a two-quantum model according to the following sequence of events: A (Mn(II)) ⇒ B (Mn(III), unstable) ⇒ dark rearrangement ⇒ C (Mn(III) + Mn(II), unstable) ⇒ D (Mn(III)-Mn(III), stable). At the first stage (A), Mn(II) cation binds to HAS, then it is oxidized by tyrosine YZ (B), followed by a mandatory dark period (the 100 ms half-time). Dark period according to one hypothesis represents the relocation of a Mn(III) ion from the HAS to its final location in the assembled cluster which is a thermodynamically more stable site (Avramov et al. 2020). A free HAS binds the second Mn(II) cation (C). Mn(II) cation is oxidized by tyrosine and a stable dimer (D) is formed. Thus, the most important step in the process of photoactivation (in fact, the reaction of Mn(II) oxidation and binding on the donor side of PSII(-Mn)) is the presence of a mandatory dark period. Taking into account the high similarity between the oxidation processes of Mn(II) and Fe(II) in the PSII(-Mn) membranes, the oxidation of Fe(II) cations in the pulsed illumination mode was studied by adjusting the flash duration and the dark interval between flashes (Semin and Seibert 2006b). The flash-probe fluorescence method was used to estimate the blocking efficiency. It has been shown that the yield of blocking increases with flash number and reaches 50% after seven flashes. When the dark interval between the flashes (Δt) was varied, it was found that the percentage of blocking decreases at Δt < 100 ms (t1/2, 4–10 ms). This result shows the necessity of a dark rearrangement during the blocking process and indicates the formation of a binuclear iron center. Schematically, the process of interaction of Fe(II) cations with Mn-depleted PSII membranes is shown in Fig. 2. An important circumstance should be noted. Concentration binding curves of Mn(II) and Fe(II) cations to the HAS are practically the same (concentration of 50% inhibition of DPC oxidation are equal 0.6 and 0.9 µM (Semin et al. 1995)), which suggests the identity of kinetic parameters. For Mn(II), such parameters (dissociation constant KD and second-order rate constant for YZ reduction k2) are respectively 0.15 µM (Hsu et al. 1987) and 6·106 M−1 s−1 (Hoganson et al. 1989). These parameters represent the binding process of cations Mn(II) or Fe(II) to HAS and their oxidation by tyrosine. The process of HAS blocking with an iron cation requires binding of at least two Fe cations (Semin et al. 1995; Semin and Seibert 2004) and oxidation of 4–5 Fe cations (Semin and Seibert 2004) and 50% blocking takes about 5 s (Semin and Seibert 2004).

Hypothetical mechanism of Fe(II) cations binding to the Mn-binding sites of OEC in the Mn-depleted PSII membranes and blocking of the high-affinity Mn-binding site. 1st Fe(II) cation binding to the HAS: [Fe(II) (solution) → Fe(II) (HAS)]; 1st Fe2+ cation photooxidation: [Fe(II) (HAS) + hν → Fe(III) + e−]; relocation of Fe(III) cation to another Mn-binding site: [Fe(III) (HAS) → Fe(III) (Mn1 site)]; 2nd Fe(II) cation binding to the HAS: [Fe(II) (solution) → Fe(II) (HAS)]; 2nd Fe(II) cation photooxidation: [Fe(II) (HAS) + hν → Fe(III) + e−]
PSII(-Mn) membranes with iron cations bound to the Mn-binding sites do not have any activity: they do not evolve oxygen under illumination (Semin et al. 2002), and do not oxidize water to hydrogen peroxide (Semin et al. 2013); i.e., there is no electronic transport in such membranes. Thus, the iron cluster replacing the entire catalytic center Mn4CaO5 in the OEC is inactive. In this regard, the question arises—will the tetranuclear manganese cluster in which 1 or 2 Mn cations are replaced by an iron cation show activity? Investigation of this problem has yielded very interesting results.
Interaction of Fe(II) cations with PSII containing manganese cluster: substitution of manganese cations in OEC with iron cations
PSII(-Ca) with 2Mn2Fe chimeric cluster
In 1984 year, Ghanotakis with colleagues investigated the protective function of extrinsic proteins (EP: PsbO, PsbQ, PsbP) (Ghanotakis et al. 1984b). The authors found that EP cover the Mn cluster, providing protection against exogenic reductants reducing Mn cations, thereby destroying the Mn cluster. EPs provide protection mainly against the action of bulky reductants such as hydroquinone (not hydroxylamine). The authors also mentioned a protector function of EP against charged reductants such as iron cations (Ghanotakis et al. 1984b). In PSII preparations without EP, for example, in the Ca-depleted PSII, the Mn cluster is available for all reducing agents. This feature made it possible to study the effect of Fe(II) cations on the Mn cluster in PSII(-Ca) membranes. The experiments were carried out on the so-called Ca-depleted PSII membranes that do not contain the Ca cation in OEC and EP PsbQ and PsbP. Such membranes are obtained by washing with a buffer with high ionic strength (Ono and Inoue 1990). Experiments showed that long-term (more than 2 h) incubation of PSII(-Ca) membranes with FeSO4 in the dark was accompanied by extraction of two Mn cations (Semin and Seibert 2016). Two Mn cations per reaction center (RC) is the maximum level of extraction. Other reductans such as H2O2 and hydroxylamine are more effective by extracting 3 Mn/RC (Table 1). Taking into account the results obtained (see above), the authors assumed that 2 Mn cations are replaced by Fe cations.
The substitution process may proceed as follows. Bivalent iron cations are quite strong reductants in a medium with a neutral pH. The mid-point reduction potential of Fe(II)/Fe(OH)3 is about 0 V at neutral pH (Widdel et al. 1993), which is significantly less than the redox potential of bound Mn cations (close to 0.9 V (Pace et al. 2012)). Fe(II) cations reduce 1 or 2 Mn cations, which are in the trivalent state (valence of at least 2 Mn cations in S1 state according to the high-valent and low-valent schemes is ≥ 3. (Najafpour et al. 2017)). This enables the monovalent reduction reaction of the Mn(III) cations by the Fe(II). Mn(II) cations leave the binding sites, and Fe(III) cations bind to free Mn-binding sites with high affinity and specificity (see above) (Fig. 1B, right panel). The following evidence indicate that iron cation is bound to at least one Mn-binding site. Selective extraction of Mn or Fe cations from the 2Mn/2Fe Fe heteronuclear cluster demonstrates that the high-affinity Mn-binding site is occupied by Fe cation (Semin and Seibert 2016). In this work, a citrate was used to extract Fe from the chimeric cluster, which has previously been shown to extract Fe cations but not Mn cations (Semin and Seibert 2006a). In favor of the possibility of the HAS in binding of the Fe(III) cation after substitution is the fact that a carboxyl-containing amino acid (possibly D1-Asp170) is involved in binding of the Mn cation in the native PSII membranes and the Fe(III) cation in the apo-PSII membranes (Semin and Seibert 2006a). Another evidence in favor of Fe binding to Mn-binding sites is a significant increase in resistance of Mn cations in the chimeric cluster to the action of hydroquinone (H2Q) (Semin et al. 2018). In PSII(-Ca) membranes, H2Q extracts 3 Mn cations from four, in PSII membranes containing 2 Mn/RC extracts 1, but does not extract any Mn cations from chimeric cluster in PSII(2Mn,2Fe) (Semin et al. 2018). The resulting chimeric cluster 2Mn2Fe is unable to synthesize molecular oxygen, but capable of oxidizing water, but at a slower rate (53%) than native preparations, and not to molecular oxygen, but to hydrogen peroxide (Semin et al. 2018).
Effect of pH on the functional activity of PSII
The rate of O2 generation by the oxygen-evolving complex in photosynthetic organisms is significantly dependent on the pH of the environment. The dependence of the oxygen-evolving activity of PSII preparations has a bell-shaped form with a maximum in the pH region of 6.0–7.0 and falling areas with 50% inhibition in the pH region of 4.8–5.5 and 7.4–8.0 (Damoder and Dismukes 1984; Schlodder and Meyer 1987; Vass and Styring 1991; Haddy et al. 1999; Schiller and Dau 2000; Semin et al. 2004). In the acidic pH region, the inactivation of OEC is largely determined by the loss of extrinsic proteins PsbP, PsbQ, and PsbO. Extrinsic proteins PsbP, PsbQ, and PsbO have corresponding pK 5.0, 4.1, and 3.6 (Shen and Inoue 1991). An increase in the concentration of protons in the medium also affects the manganese cluster of OEC, including the S transitions of the catalytic cycle. In Bernát et al., and Suzuki et al., almost identical results were obtained in PSII preparations isolated from spinach (Bernát et al. 2002) and thermophilic cyanobacteria Thermosynechoccocus elongatus (Suzuki et al. 2005). In the experiments performed, it was found that in the OEC of spinach S1 → S2, the transition is independent of pH in the region 4.1–8.4, while in S2 → S3, S3 → [S4] → S0, and S0 → S1 the transitions have pK of 4.0, 4.5, and 4.7, respectively (Bernát et al. 2002).
Resistance of Mn cations in the oxygen-evolving complex of PSII to the action of reductants at different pH
Investigating the pH influence on the effect of reductants on the manganese cluster found that in the pH region 5.7, there is an increase in the resistance of one of the Mn cations to its reduction by reductant. If three out of four manganese cations were extracted at pH 6.5 by H2Q and H2O2, only two Mn cations were extracted at pH 5.7 (Table 1). The same is the case with Fe(II). Fe(II) cations extract two Mn cations at pH 6.5 and only one at pH 5.7 (Table 1). Thus, pH-dependent resistance to exogenous reducing agents exists only for one manganese cation that is part of the manganese OEC cluster. Since the extraction of manganese cations occurs as a result of their reduction, the results obtained indicate the important role of the ratio of redox potentials of the reducing agent and manganese cations in this process (Kuntzleman et al. 2004). In this regard, it can be assumed that an increase in resistance to the action of reducing agents of one of the manganese cations in the cluster is associated with a decrease in its redox potential with a decrease in pH.
Ca2+ effect on redox property of Mn cluster
The catalytic center of OEC contains Ca2+ cation, the function of which, despite intensive research, remains largely unclear. Recently, a new hypothesis relative to Ca2+ function has appeared based on model experiments. Tsui and Agapie (2013) found a linear relationship between the redox potential of the heterometallic metal-oxide cluster and the Lewis acidity of the redox inactive metal cation. The authors hypothesized that this correlation is evidence of the involvement of calcium cation in modulating the redox potential of the manganese cluster. Thus, the calcium cation in OEC can affect the redox potential of one or more manganese cations and regulate the efficiency of reducing manganese cations with exogenous redox agents. In this regard, the effect of Ca2+ cations on the efficiency of Mn extraction by hydroquinone and Fe(II) cations was investigated. The results are presented in Table 1 and show the effect of Ca2+ cation on the reduction of manganese cations in OEC.
Incubation of PSII(-Ca) membranes in the presence of Ca2+ with hydroquinone prevents the extraction of one manganese cation at pH 6.5 but does not affect the extraction process at pH 5.7 (Table 1). The inhibitory effect of Ca2+ on the extraction of manganese cations from OEC by hydroquinone and iron cations confirms the possibility of the influence of the Ca2+ cation on the redox potential of one or more manganese cations in OEC.
The above results suggest that at pH 5.7, either the oxygen bridge/bridges or one or more amino acid groups of the manganese cluster or its immediate environment is protonated. This process is accompanied by a decrease of the redox potential of one the manganese cations, making it inaccessible to the reducing agent. It should be noted that the pH dependence of the oxygen-evolving activity of the membrane preparations, measured in sufficient detail in 0.1 pH increments, has a small arm at pH 5.7 (Schiller and Dau 2000). A more prominent arm in pH dependence of PSII functional activity at pH 5.3 appears in D1-S169A and D2-K317A mutants (Ghosh et al. 2019).
PSII(-Ca) with 3Mn1Fe chimeric cluster without extrinsic proteins PsbP and PsbQ
Taking into account that the extraction of Mn cations from OEC by iron cations is accompanied in fact by the substitution of Mn cations by Fe cations (see above) (Fig. 1B, right panel), the process of interaction of Fe(II) cations with PSII(-Ca) membranes was investigated in more detail. It was found only 1Mn/RC extraction, that is, the formation of a chimeric cluster in OEC (Semin et al. 2018). It was also shown that the presence of Fe cations in the 3Mn1Fe heteronuclear cluster increases the resistance of the remaining Mn cations to H2Q action, since this reducing agent is able to extract Mn cations from homonuclear Mn clusters of PSII(-Ca,4Mn) and PSII(-Ca,2Mn) membranes but not from the heteronuclear cluster in PSII(-Ca,3Mn,1Fe) membranes. This fact indicates that the cluster contains an iron cation stabilizing the Mn cations, as in the case of the chimeric cluster 2Mn2Fe. Interestingly, extraction of the Fe cation from the 3Mn1Fe cluster is accompanied by a loss of 1 Mn cation, which confirms the stabilizing role of Fe in the chimeric cluster. Functional activity of PSII with a 3Mn1Fe cluster was investigated. PSII preparations with a 3Mn1Fe cluster in the OEC are able to photoreduce the exogenous electron acceptor 2,6-dichlorophenolindophenol, possibly due to incomplete oxidation of water molecules as is the case with PSII(-Ca,2Mn,2Fe) samples. However, in the contrast to PSII(-Ca,2Mn,2Fe) samples, PSII(-Ca,3Mn,1Fe) membranes can evolve O2 at a low rate in the presence of exogenous Ca2+ (at about 27% of the rate of O2 evolution in native PSII membranes). The possibility of water oxidation by a chimeric cluster with oxygen release is of interest for studying the mechanism of photosynthetic oxidation of water (identification of manganese cations that play a general role in water photolysis, etc.), as well as for the development of artificial water photolysis systems as generators of molecular oxygen and hydrogen.
PSII(-Ca) with 3Mn1Fe chimeric cluster with extrinsic proteins
To replace Mn cations in OEC, it is necessary to ensure their availability for Fe cations. To this end, in previous studies (Semin and Seibert 2016; Semin et al. 2018), PSII membranes were treated with a high-molar solution of NaCl, which provided extraction of extrinsic proteins PsbP and PsbQ and Ca2+ cation from membranes. To preserve all extrinsic proteins in the PSII preparation, another method of Ca2+ extracting from OEC was used—the method developed by Ono and Inoue (1988). This method (treatment of the PSII membranes with citrate buffer with acidic pH) allows to obtain the PSII membranes without Ca2+ cation in OEC but with EP. But, crucially, unlike the native PSII membranes in citrate-treated PSII (-Ca) membranes, the Mn cluster is available for small-sized reducing agents (Vander Meulen et al. 2002, 2004). Indeed, one Mn cation in OEC found to be replaced by an Fe cation after incubation of Ca-depleted PSII membranes with Fe(II) cations after removal of citrate; i.e., PSII membranes with a chimeric cluster 3Mn1Fe in OEC were obtained in the presence of all extrinsic proteins. It is important to note that there are 2 significant differences between the characteristics of PSII preparations with a chimeric cluster 3Mn1Fe with (Semin and Davletshina 2023) and without EP (Semin et al. 2018). In preparations prepared by replacing Mn cation in the presence of EP (Semin and Davletshina 2023), only 1 Mn cation is substituted at pH 6.5, while without EP at the same pH 2 Mn cations are replaced (substitution of one cation occurs only at pH 5.7) (Semin et al. 2018). But the most significant difference in preparations is their evolving activity in the presence of exogenous Ca2+. PSII preparations with a chimeric cluster without EP have an activity of about 25% (Semin et al. 2018), while in the presence of EP their activity increases to 80% (Semin and Davletshina 2023). What could be the reason for such differences? Taking into account the fact that EP increases the redox potential of one of OEC Mn cations (Fe(II) extracts 2Mn at pH 6.5 in the absence of EP, but 1Mn in the presence of EP), we can assume that the influence of EP is carried out in the step before the substitution of Mn cation with Fe(II) cation, and not after.
Conclusion
This review summarized the recent progress and understanding associated with the replacement of manganese cations in OEC with Fe cations. Semin with colleagues discovered highly efficient and highly specific binding of iron cations to Mn-binding sites in Mn-depleted PSII (Semin et al. 1995; Semin et al. 2002, 2005; Semin and Seibert 2004, 2006a,b). Using this feature, Semin and co-authors developed a method for producing chimeric clusters in OEC consisting of iron and manganese cations in different ratios (Semin and Seibert 2016; Semin et al. 2018). It should be noted that recently there has been increasing interest in the possible use of metal cations located in the row with Mn, either in the replacement of Mn cations in the OEC, or in the creation of artificial water oxidation catalysts (Saito et al. 2023; Gates et al. 2022; Kondo et al. 2021). Saito et al. (2023) investigated the Fe4CaO5 cluster in the PSII protein environment using a quantum mechanical/molecular mechanical approach as the first step in resolving the issue why Mn and not Fe is employed by nature in the water oxidation catalyst. The results obtained showed significant differences between the protonation states and oxidation states of the Fe4CaO5 cluster and Mn4CaO5 cluster. Gates with coworkers obtained photoactivation of OEC using Co2+ ions instead of Mn2+ in combination with Ca2+ and water (Gates et al. 2022), although the activity of the reconstituted preparations was significantly less (25%) than in the case of Mn(II). Mn, Fe, Co, Ni, and Cu cations are being intensively investigated in the works on the creation of artificial water-splitting catalysts (see review Kondo et al. 2021).
Studies of the possibility of replacing Mn cations in OEC with cations of other metals may be of interest to clarify the mechanism of water oxidation and molecular oxygen synthesis. In this regard, we want to draw attention to the following interesting fact obtained during the study of the chimeric cluster 3Mn1Fe (Semin and Davletshina 2023). Currently, there are a number of hypotheses for the mechanism of water oxidation (Shen 2015; Lubitz et al. 2019; Oliver et al. 2022; Shevela et al. 2023). All of them assume participation in the oxidation of water and the synthesis of O2 of all 4 Mn cations, including the cation (Mn4) associated with HAS, which implies its oxidation during the Kok cycle. In this regard, it should be noted that according to Semin and Davletshina (2023), the chimeric cluster containing not 4, but 3 Mn has a very high oxygen-evolving activity (about 80%) in the presence of exogenous Ca2+ and EP. This fact suggests that the oxidation of the H2O occurs with the participation of three Mn cations, since the Fe cation in the cluster is in the 3-valence state, and further oxidation of it requires very strong oxidizing agents. This data suggests that the Mn cation bound to HAS and HAS itself are not the main participants of O2 synthesis. More likely, HAS is mostly involved in resynthesis of Mn cluster, destroyed by photoinhibition process. In fact, this scheme is discussed now in literature (see review Oliver et al. 2022).
It should be noted that iron cations embedded in the chimeric cluster increase the resistance of Mn cations to the action of exogenic reductants (Semin et al. 2018). Combined with the high catalytic activity of the chimeric cluster 3Mn1Fe in the presence of extrinsic proteins (Semin and Davletshina 2023), this object is of interest for more detailed studies in order to create artificial energy systems for water decomposition. In this regard, studies aimed at increasing the oxygen-evolving activity of PSII preparations with a chimeric cluster are of interest, namely the development of new methods for the reconstruction of extrinsic proteins and the preparation of chimeric clusters in the presence of Ca2+, which, as it was found not so long ago, is very important for synthesis of Mn cluster during the photoactivation process (Avramov et al. 2020). In addition, a study of the possibility of synthesis on the donor side of PSII(-Mn) an iron-sulfur cluster is of interest, taking into account the high affinity of Fe cations binding to Mn-binding sites. Such preparations would be interesting in terms of the possibility of catalytic activity of such clusters and evolutionary origin PSII. Computational approaches can be used in parallel with experimental studies. They are able to provide a number of interesting conclusions regarding the structure and functioning of OEC with chimeric clusters.
Data availability
All data supporting the findings of this study are available within the paper including references.
Abbreviations
- Chl:
-
Chlorophyll
- D1:
-
Integral protein of the photosystem II reaction center
- DPC:
-
Diphenylcarbazide
- EP:
-
Extrinsic proteins
- HAS:
-
High-affinity Mn-binding site
- H2Q:
-
Hydroquinone
- LAS:
-
Low-affinity Mn-binding site
- OEC:
-
Oxygen-evolving complex
- PSII:
-
Photosystem II
- PSII(-Ca) :
-
Ca2+-depleted PSII membranes
- PSII(-Mn) :
-
Mn-depleted PSII membranes
- RC:
-
Reaction center
- Tris:
-
Tris(hydroxymethyl)amino methane
- SOD:
-
Superoxide dismutase
- YZ :
-
Redox-active tyrosine D1-Tyr161, the first electron donor to P680+ in PSII
References
Atta M, Nordlund P, Aberg A, Eklund H, Fontecave M (1992) Substitution of manganese for iron in ribonucleotide reductase from Escherichia coli. Spectroscopic and crystallographic characterization. J Biol Chem 267(29):20682–20688. https://doi.org/10.1016/S0021-9258(19)36739-0
Avramov AP, Hwang HJ, Burnap RL (2020) The role of Ca2+ and protein scaffolding in the formation of nature’s water oxidizing complex. Proc Natl Acad Sci USA 117:28036–28045. https://doi.org/10.1073/pnas.2011315117
Bernát G, Morvaridi F, Feyziyev Y, Styring S (2002) pH dependence of the four individual transitions in the catalytic S-cycle during photosynthetic oxygen evolution. Biochemistry 41:5830–5843. https://doi.org/10.1021/bi011691u
Charley PJ, Sarkar B, Stitt CF, Saltman P (1963) Chelation of iron by sugars. Biochim Biophys Acta 69:313–321. https://doi.org/10.1016/0006-3002(63)91264-2
Damoder R, Dismukes GC (1984) pH dependence of the multiline, manganese EPR signal for the ’S2’ state in PS II particles. Absence of proton release during the S1 → S2 electron transfer step of the oxygen evolving system. FEBS Lett 174:157–161. https://doi.org/10.1016/0014-5793(84)81096-0
Gates C, Ananyev G, Roy-Chowdhury S, Cullinane B, Miller M, Fromme P, Dismukes GC (2022) Why did nature choose manganese over cobalt to make oxygen photosynthetically on the earth? J Phys Chem B 126(17):3257–3268. https://doi.org/10.1021/acs.jpcb.2c00749
Ghanotakis DF, Babcock GT, Yocum CF (1984a) Calcium reconstitutes high rates of oxygen evolution in polypeptide depleted photosystem II preparations. FEBS Lett 167:127–130. https://doi.org/10.1016/0014-5793(84)80846-7
Ghanotakis DF, Topper JN, Yocum CF (1984b) Exogenous reductants reduce and destroy the Mn-complex in photosystem II membranes depleted of the 17 and 23 kDa polypeptides. Biochim Biophys Acta 767:524–531. https://doi.org/10.1016/0005-2728(84)90051-3
Ghanotakis DF, Babcock GT, Yocum CF (1985) Structure of the oxygen-evolving complex of Photosystem II: calcium and lanthanum compete for sites on the oxidizing side of Photosystem II which control the binding of water-soluble polypeptides and regulate the activity of the manganese complex. Biochim Biophys Acta 809:173–180. https://doi.org/10.1016/0005-2728(85)90060-X
Ghirardi ML, Lutton TW, Seibert M (1996) Interactions between diphenylcarbazide, zinc, cobalt, and manganese on the oxidizing side of photosystem II. Biochemistry 35:1820–1828. https://doi.org/10.1021/bi951657d
Ghosh I, Khan S, Banerjee G, Dziarski A, Vinyard DJ, Debus RJ, Brudvig GW (2019) Insights into proton-transfer pathways during water oxidation in photosystem II. J Phys Chem B 123:8195–8202. https://doi.org/10.1021/acs.jpcb.9b06244
Gisriel CJ, Zhou K, Huang H-L, Debus RJ, Xiong Y, Brudvig GW (2020) Cryo-EM structure of monomeric photosystem II from Synechocystis sp. PCC 6803 lacking the water-oxidation complex. Joule 4(10):2131–2148. https://doi.org/10.1016/j.joule.2020.07.016
Haddy A, Hatchell JA, Kimel RA, Thomas R (1999) Azide as a competitor of chloride in oxygen evolution by photosystem II. Biochemistry 38:6104–6110. https://doi.org/10.1021/bi983075c
Hoganson CW, Ghanotakis DF, Babcock GT, Yocum CF (1989) Mn2+ reduces YZ+ in manganese-depleted Photosystem II preparations. Photosynth Res 22:285–293. https://doi.org/10.1007/BF00048306
Hsu BD, Lee J-Y, Pan R-L (1987) The high-affinity binding site for manganese on the oxidizing side of photosystem II. Biochim Biophys Acta 890(1):89–96. https://doi.org/10.1016/0005-2728(87)90072-7
Kálmán L, Flores M, Williams JC, Allen JP (2010) Electronic structure of Fe3+ at a metal-binding site introduced in modified bacterial reaction centers. Appl Magn Reson 37:27–37. https://doi.org/10.1007/s00723-009-0037-z
Kawakami K, Umena Y, Kamiya N, Shen J-R (2011) Structure of the catalytic, inorganic core of oxygen-evolving photosystem II at 1.9 Å resolution. J Photochem Photobiol B 104(1–2):9–18. https://doi.org/10.1016/j.jphotobiol.2011.03.017
Kern J, Chatterjee R, Young ID, Fuller FD, Lassalle L, Ibrahim M, Gul S, Fransson T, Brewster AS, Alonso-Mori R et al (2018) Structures of the intermediates of Kok’s photosynthetic water oxidation clock. Nature 563:421–425. https://doi.org/10.1038/s41586-018-0681-2
Kondo M, Tatewaki H, Masaoka S (2021) Design of molecular water oxidation catalysts with earth-abundant metal ions. Chem Soc Rev 50(12):6790–6831. https://doi.org/10.1039/D0CS01442G
Kudasheva DS, Lai J, Ulman A, Cowman MK (2004) Structure of carbohydrate-bound polynuclear iron oxyhydroxide nanoparticles in parenteral formulations. J Inorg Biochem 98:1757–1769. https://doi.org/10.1016/j.jinorgbio.2004.06.010
Kuntzleman T, McCarrick R, Penner-Hahn J, Yocum C (2004) Probing reactive sites within the photosystem II manganese cluster: evidence for separate populations of manganese that differ in redox potential. Phys Chem Chem Phys 6:4897–4904. https://doi.org/10.1039/B406601D
Kurashov VN, Lovyagina ER, Shkolnikov DYu, Solntsev MK, Mamedov MD, Semin BK (2009) Investigation of the low-affinity oxidation site for exogenous electron donors in the Mn-depleted photosystem II complexes. Biochim Biophys Acta 1787:1492–1498. https://doi.org/10.1016/j.bbabio.2009.07.002
Lah MS, Dixon MM, Pattridge KA, Stallings WC, Fee JA, Ludwig ML (1995) Structure-function in Escherichia coli iron superoxide dismutase: comparisons with the manganese enzyme from Thermus thermophilus. Biochemistry 34(5):1646–1660. https://doi.org/10.1021/bi00005a021
Lovyagina ER, Davletshina LN, Kultysheva MYu, Timofeev KN, Ivanov II, Semin BK (2005) Characteristic features of the interaction between Fe(II) cations and the donor side of the manganese-depleted photosystem II. Russ J Plant Physiol 52(1):7–14. https://doi.org/10.1007/s11183-005-0002-0
Lovyagina ER, Loktyushkin AV, Semin BK (2021) Effective binding of Tb3+ and La3+ cations on the donor side of Mn-depleted photosystem II. J Biol Inorg Chem 26:1–11. https://doi.org/10.1007/s00775-020-01832-w
Lubitz W, Chrysina M, Cox N (2019) Water oxidation in photosystem II. Photosynth Res 142(1):105–125. https://doi.org/10.1007/s11120-019-00648-3
Najafpour M, Heidari S, Balaghi SE, Hołyńska M, Sadr MH, Soltani B, Khatamian M, Larkum AW, Allakhverdiev SI (2017) Proposed mechanisms for water oxidation by Photosystem II and nanosized manganese oxides. Biochim Biophys Acta 2:156–174. https://doi.org/10.1016/j.bbabio.2016.11.007
Oliver N, Avramov AP, Nürnberg DJ, Dau H, Burnap RL (2022) From manganese oxidation to water oxidation: assembly and evolution of the water-splitting complex in photosystem II. Photosynth Res 152(2):107–133. https://doi.org/10.1007/s11120-022-00912-z
Ono T (2000) Effects of lanthanide substitution at Ca2+-site on the properties of the oxygen evolving center of photosystem II. J Inorg Biochem 82:85–91. https://doi.org/10.1016/S0162-0134(00)00144-6
Ono T, Inoue Y (1988) Discrete extraction of the Ca atom functional for O2 evolution in higher plant photosystem II by a simple low pH treatment. FEBS Lett 227:147–152. https://doi.org/10.1016/0014-5793(88)80886-X
Ono T, Inoue Y (1990) Abnormal redox reactions in photosynthetic O2-evolving centers in NaCl/EDTA-washed PS II. A dark-stable EPR multiline signal and an unknown positive charge accumulator. Biochim Biophys Acta 1020:269–277. https://doi.org/10.1016/0005-2728(90)90157-Y
Pace RJ, Stranger R, Petrie S (2012) Why nature chose Mn for the water oxidase in photosystem II. Dalton Trans 41(24):7179–7189. https://doi.org/10.1039/C2DT30185G
Saito K, Shen J-R, Ishida T, Ishikita H (2011) Short hydrogen bond between redox-active tyrosine YZ and D1-His190 in the photosystem II crystal structure. Biochemistry 50:9836–9844. https://doi.org/10.1021/bi201366j
Saito K, Nakagawa M, Mandal M, Ishikita H (2021) Role of redox-inactive metals in controlling the redox potential of heterometallic manganese–oxido clusters. Photosynth Res 148:153–159. https://doi.org/10.1007/s11120-021-00846-y
Saito M, Saito K, Ishikita H (2023) Structural and energetic insights into Mn-to-Fe substitution in the oxygen-evolving complex. iScience 26(8):107352. https://doi.org/10.1016/j.isci.2023.107352
Schiller H, Dau H (2000) Preparation protocols for high-activity photosystem II membrane particles of green algae and higher plants, pH dependence of oxygen evolution and comparison of the S2-state multiline signal by X-band EPR spectroscopy. J Photochem Photobiol B 55:138–144. https://doi.org/10.1016/S1011-1344(00)00036-1
Schlodder E, Meyer B (1987) pH dependence of oxygen evolution and reduction kinetics of photooxidized chlorophyll aII (P-680) in Photosystem II particles from Synechococcus sp. Biochim Biophys Acta 890:23–31. https://doi.org/10.1016/0005-2728(87)90064-8
Semin BK, Davletshina LN (2023) High-efficiency oxygen evolution by photosystem II oxygen-evolving complex containing 3Mn per reaction center. J Biol Inorg Chem 28:393–401. https://doi.org/10.1007/s00775-023-01987-2
Semin BK, Davletshina LN, Rubin AB (2015) Correlation between pH dependence of O2 evolution and sensitivity of Mn cations in the oxygen-evolving complex to exogenous reductants. Photosynth Res 125:95−103. https://doi.org/10.1007/s11120-015-0155-4
Semin BK, Seibert M (2004) Iron bound to the high-affinity Mn-binding site of the oxygen-evolving complex shifts the pK of a component controlling electron transport via Y(Z). Biochemistry 43:6772–6782. https://doi.org/10.1021/bi036047p
Semin BK, Seibert M (2006a) A carboxylic residue at the high-affinity, Mn-binding site participates in the binding of iron cations that block the site. Biochim Biophys Acta 1757(3):189–197. https://doi.org/10.1016/j.bbabio.2006.02.001
Semin BK, Seibert M (2006b) Flash-induced blocking of the high-affinity manganese-binding site in photosystem II by iron cations: dependence on the dark interval between flashes and binary oscillations of fluorescence yield. J Phys Chem B 110:25532–25542. https://doi.org/10.1021/jp0652796
Semin BK, Seibert M (2016) Substituting Fe for two of the four Mn ions in photosystem II–effects on water-oxidation. J Bioenerg Biomembr 48:227–240. https://doi.org/10.1007/s10863-016-9651-2
Semin BK, Ivanov II, Rubin AB, Parak F (1995) High-specific binding of Fe(II) at the Mn-binding site in Mn-depleted PSII membranes from spinach. FEBS Lett 375:223–226. https://doi.org/10.1016/0014-5793(95)01215-Z
Semin BK, Ghirardi ML, Seibert M (2002) Blocking of electron donation by Mn(II) to YZ• following incubation of Mn-depleted photosystem II membranes with Fe(II) in the light. Biochemistry 41:5854–5864. https://doi.org/10.1021/bi0200054
Semin BK, Davletshina LN, Aleksandrov AYu, Lanchinskaya VYu, Novakova AA, Ivanov II (2004) pH dependence of iron binding to the donor side of photosystem II. Biochem Mosc 69:410–419. https://doi.org/10.1023/B:BIRY.0000022066.38297.8a
Semin BK, Lovyagina ER, Timofeev KN, Ivanov II, Rubin AB, Seibert M (2005) Iron-blocking the high-affinity Mn-binding site in photosystem II facilitates identification of the type of hydrogen bond participating in proton-coupled electron transport via YZ. Biochemistry 44:9746–9757. https://doi.org/10.1021/bi047618w
Semin BK, Davletshina LN, Timofeev KN, Ivanov II, Rubin AB, Seibert M (2013) Production of reactive oxygen species in decoupled, Ca2+-depleted PSII and their use in assigning a function to chloride on both sides of PSII. Photosynth Res 117(1):385–399. https://doi.org/10.1007/s11120-013-9870-x
Semin BK, Davletshina LN, Seibert M, Rubin AB (2018) Creation of a 3Mn/1Fe cluster in the oxygen-evolving complex of photosystem II and investigation of its functional activity. J Photochem Photobiol B 178:192–200. https://doi.org/10.1016/j.jphotobiol.2017.11.016
Semin BK, Davletshina LN, Goryachev SN, Seibert M (2021) Ca2+ effects on Fe(II) interactions with Mn-binding sites in Mn-depleted oxygen-evolving complexes of photosystem II and on Fe replacement of Mn in Mn-containing. Ca-Depleted Complexes Photosynth Res 147(2):229–237. https://doi.org/10.1007/s11120-020-00813-z
Shen JR (2015) The structure of photosystem II and the mechanism of water oxidation in photosynthesis. Annu Rev Plant Biol 66:23–48. https://doi.org/10.1146/annurev-arplant-050312-120129
Shen J-R, Inoue Y (1991) Low pH-induced dissociation of three extrinsic proteins from O2-evolving photosystem II. Plant Cell Physiol 32(3):453–457. https://doi.org/10.1093/oxfordjournals.pcp.a078101
Shevela D, Kern JF, Govindjee G, Messinger J (2023) Solar energy conversion by photosystem II: principles and structures. Photosynth Res 156:279–307. https://doi.org/10.1007/s11120-022-00991-y
Sillen LG, Marten AE (1964) Stability constants of metal–ion complexes, Special Publ 17. The Chemical Society, Burlington House, London, p W1
Suga M, Akita F, Hirata K, Ueno G, Murakami H, Nakajima Y, Shimizu T, Yamashita K, Yamamoto M, Ago H, Shen J-R (2015) Native structure of photosystem II at 1.95 Å resolution viewed by femtosecond X-ray pulses. Nature 517:99–103. https://doi.org/10.1038/nature13991
Suga M, Akita F, Sugahara M, Kubo M, Nakajima Y, Nakane T, Yamashita K, Umena Y, Nakabayashi M, Yamane T et al (2017) Light-induced structural changes and the site of O=O bond formation in PSII caught by XFEL. Nature 543:131–135. https://doi.org/10.1038/nature21400
Suga M, Akita F, Yamashita K, Nakajima Y, Ueno G, Li H, Yamane T, Hirata K, Umena Y, Yonekura S et al (2019) An oxyl/oxo mechanism for oxygen-oxygen coupling in PSII revealed by an X-ray free-electron laser. Science 366(6463):334–338. https://doi.org/10.1126/science.aax6998
Suzuki H, Sugiura M, Noguchi T (2005) pH dependence of the flash-Induced S-state transitions in the oxygen-evolving center of photosystem II from Thermosynechoccocus elongatus as revealed by Fourier transform infrared spectroscopy. Biochemistry 44:1708–1718. https://doi.org/10.1021/bi0483312
Tamura N, Cheniae GM (1987) Photoactivation of the water-oxidizing complex in Photosystem II membranes depleted of Mn and extrinsic proteins. I. Biochemical and kinetic characterization. Biochim Biophys Acta 890:179–194. https://doi.org/10.1016/0005-2728(87)90019-3
Tang K, Williams JC, Allen JP, Kálmán L (2009) Effect of anions on the binding and oxidation of divalent manganese and iron in modified bacterial reaction centers. Biophys J 96(8):3295–3304. https://doi.org/10.1016/j.bpj.2009.01.027
Tsui EY, Agapie T (2013) Reduction potentials of heterometallic manganese–oxido cubane complexes modulated by redox-inactive metals. Proc Natl Acad Sci USA 110:10084–10088. https://doi.org/10.1073/pnas.1302677110
Tsui EY, Tran R, Yano J, Agapie T (2013) Redox-inactive metals modulate the reduction potential in heterometallic manganese-oxido clusters. Nat Chem 5(4):293–299. https://doi.org/10.1038/nchem.1578
Umena Y, Kawakami K, Shen J-R, Kamiya N (2011) Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 473:55–60. https://doi.org/10.1038/nature09913
Vander Meulen KA, Hobson A, Yocum CF (2002) Calcium depletion modifies the structure of the photosystem II O2-evolving complex. Biochemistry 41:958–966. https://doi.org/10.1021/bi0109414
Vander Meulen KA, Hobson A, Yocum CF (2004) Reconstitution of the photosystem II Ca2+ binding site. Biochim Biophys Acta 1655:179–183. https://doi.org/10.1016/j.bbabio.2003.08.012
Vass I, Styring S (1991) pH-Dependent charge equilibria between tyrosine-D and the S states in photosystem II. Estimation of relative midpoint redox potentials. Biochemistry 30:830–839. https://doi.org/10.1021/bi00217a037
Vrettos JS, Stone DA, Brudvig GW (2001) Quantifying the ion selectivity of the Ca2+ site in photosystem II. Evidence for direct involvement of Ca2+ in O2 formation. Biochemistry 40:7937–7945. https://doi.org/10.1021/bi010679z
Waggoner CM, Yocum CF (1990) Calcium activated oxygen evolution. In: Baltscheffsky M (ed) Current research in photosynthesis, Springer, Dordrecht, pp 733–736. https://doi.org/10.1007/978-94-009-0511-5_167
Widdel F, Schnell S, Heising S, Ehrenreich A, Assmus B, Schink B (1993) Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 362(6423):834–836. https://doi.org/10.1038/362834a0
Zhang M, Bommer M, Chatterjee R, Hussein R, Yano J, Dau H, Kern J, Dobbek H, Zouni A (2017) Structural insights into the light-driven auto-assembly process of the water-oxidizing Mn4CaO5-cluster in photosystem II. eLife 6:e26933. https://doi.org/10.7554/eLife.26933
Funding
The research was carried out as part of the Scientific Project of the State Order of the Government of Russian Federation to Lomonosov Moscow State University No. 121032500058–7.
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Semin, B., Loktyushkin, A. & Lovyagina, E. Current analysis of cations substitution in the oxygen-evolving complex of photosystem II. Biophys Rev 16, 237–247 (2024). https://doi.org/10.1007/s12551-024-01186-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12551-024-01186-6