
Abstract
New grylloblattidan insect specimens from the early Permian (Asselian to early Sakmarian) Meisenheim Formation of the Saar-Nahe Basin in southwest Germany are described. The most abundant specimens are assigned to Pictoborella germanica (Prokop et al. in Geodiversitas 34: 271–281, 2012) and to Oborella brauckmanni Prokop et al., 2012. Morphological variation in these taxa is documented and discussed. Two new species, Oborella moschelensis sp. nov., and Artinska glanensis sp. nov., represented by one and three specimens, respectively, are proposed. A possible forewing of undetermined Liomopteridae is also recorded. With respect to grylloblattidans, the insects of the Meisenheim Formation reveal a decidedly Permian character.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Extant grylloblattid insects are a relict group comprising about 30 wingless species in a single family, the Grylloblattidae (Vršanský et al. 2001; Cui et al. 2019). In contrast, many Palaeozoic and Mesozoic winged taxa have been regarded as belonging to this group based on symplesiomorphies. Thus their attribution to the extant clade is debatable and the Grylloblattodea comprising all fossil taxa may be paraphyletic or even polyphyletic (Vršanský et al. 2001; Aristov et al. 2009). However, these insects are known since the late Carboniferous (Gzhelian) and were widespread, common and diverse in Permian insect assemblages (e.g., Beckemeyer and Hall 2007; Aristov 2010; Myskowiak et al. 2015; Rasnitsyn et al. 2015; Cui et al. 2019), especially from Gondwana, where they may account for up to 35% of the insect remains (Pretorius et al. 2021). After the Permian the order declined and the winged grylloblattodeans eventually vanished in the Late Cretaceous (Rasnitsyn et al. 2004; Zhang et al. 2022). The early Permian Niedermoschel assemblage from the Saar-Nahe Basin in southwest Germany comprises a large number of grylloblattodean insects (Hörnschemeyer 1999). It is of considerable significance, as one of the earliest such assemblages bridging the time span from the earliest Permian (Gzehlian–Asselian) Carrizo Arroyo assemblage (e.g., Rasnitsyn et al. 2004; Prokop and Kukalová-Peck 2017) to the Sakmarian or Artinskian Obora assemblage (e.g., Kukalová 1964).
Here we describe a number of grylloblattodean fossils in the State Collection of Natural History of Rhineland-Palatinate at the Natural History Museum Mainz originating mostly from the Niedermoschel black shale, and some additional fossils from other exposures of the Meisenheim Formation. This significantly improves our knowledge of the diversity and intraspecific variability of grylloblattodean insects from the early Permian of the Saar-Nahe Basin.
Materials and methods
The fossils described herein originate from the early Permian Meisenheim Formation of the Saar-Nahe Basin (Fig. 1), which has an Asselian age possibly ranging into the Sakmarian (Menning et al. 2022) and is well-known for the rich occurrences of vertebrates exceptionally preserved in profundal facies of larger freshwater lakes (e.g., Schindler and Heidtke 2007 and references therein). In contrast, the Permian palaeoentomofauna of the Saar-Nahe Basin is comparatively poorly known and, following the works of Guthörl in the last century (see Guthörl 1964), has only recently re-gained large-scale scientific interest. This is mostly due to evaluation of the rich and diverse insect finds from the so-called Niedermoschel black shale (Poschmann et al. 2021).

Simplified geology of the Saar-Nahe Basin, with mentioned localities and stratigraphical column (with approximate ages, mya) and stratigraphical positions of Humberg, Morsbacher Hof, and Niedermoschel black shales approximately indicated (modified from Poschmann and Schindler 2023)
The fossils were examined using a Leica MZ 7.5 stereomicroscope both dry and immersed in isopropanol. Photographs were taken with specimens under isopropanol using a Canon EOS 600D SLR camera equipped with a Canon MP-E 65 mm macro lens. Original photographs were processed using the image-editing software Adobe Photoshop. Drawings were made from enlarged photographs using Inkscape. Most specimens are deposited in the Landessammlung für Naturkunde Rheinland-Pfalz at the Naturhistorisches Museum Mainz (NHMMZ), three specimens (formerly Raisch collection) are deposited in the Urweltmuseum GEOSKOP, Burg Lichtenberg (UGKU).
The wing venation nomenclature generally follows the concept of Kukalová-Peck (1991). Note that the first anal vein may alternatively be regarded as a postcubital vein sensu Schubnel et al. (2020).
Wing venation abbreviations: AA, Analis anterior; C, costal vein; CuA/CuP, cubitus anterior/posterior; M, median vein; RA/RP, radius anterior/posterior; R, radius; ScP, subcosta posterior.
Systematic palaeontology
Order Grylloblattida Walker, 1914
Clade Lemmatophorina Storozhenko, 1997
Family Lemmatophoridae Sellards, 1909
Remarks. We follow the proposal of Cui et al. (2022) and attribute the specimens herein assigned to Pictoborella to Lemmatophoridae rather than to Euryptilonidae, because MA is generally simple within Lemmatophoridae. M and CuA are often shortly fused just distad their bases in the new specimens, but this character seems to be somewhat unstable.
Genus Pictoborella Cui et al. 2022
Type species. Pictoborella clara Cui et al. 2022, from Cabarz and Sperbersbach, Germany; early Permian.
Other species. Pictoborella germanica (Prokop et al., 2012), from the Saar-Nahe Basin, Germany; early Permian.
Diagnosis (forewing only). See Cui et al. (2022).
Pictoborella germanica (Prokop et al., 2012)

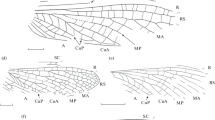
Pictoborella germanica (Prokop et al., 2012), Jeckenbach Subformation; photographs and explanatory drawings, respectively. a Forewing NHMMZ PE 1995/7778-LS (flipped horizontally), Niedermoschel; b forewing NHMMZ PE 1995/7748-LSa (flipped horizontally), Niedermoschel; c forewing PE 2021/5001-LS, Niedermoschel; d forewing UGKU 3167, Sitters; e forewing NHMMZ PE 1995/7735-LS (flipped horizontally), Niedermoschel. (Scale bars = 5 mm)

Pictoborella germanica (Prokop et al., 2012), Humberg black shale of Freisen-Grügelborn; photographs and explanatory drawings, respectively. a Forewing UGKU 3166; b forewing UGKU 3165 (flipped horizontally). (Scale bars = 5 mm)
Diagnosis. 10–14 crossveins in forewing costal area, in one or two rows; RP forked; M in forewing forked close to separation from common base or anastomosis with CuA1; MA simple; MP forked; CuA with two main branches, CuA1 with two or three branches, CuA2 simple; forewing with striking colouration pattern covering a large area especially in the apical part of wing (modified from Prokop et al. 2012).
Type locality and type stratum. Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation, Jeckenbach Subformation.
Remarks. Prokop et al. (2012) gave the locality data for their material as “Odernheim town, Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation, Odernheim Subformation, Odernheim lake horizon (L-O8 or M8 respectively), lacustrine black shales”. Personal communication with one of the original authors and collector of the type material revealed that “Odernheim town site” is actually referring to a number of different localities or outcrops within the Meisenheim Formation subsuming different stratigraphic levels under that “locality” name (Nel and Poschmann 2020). Thus, the exact stratigraphic level of this material is equivocal. Judging from the figured specimens, their preservation and lithology, as well as the fact that there are several specimens (type material of Oborella germanica and O. brauckmanni of the original publication), we strongly suspect the Niedermoschel black shale and therefore the Jeckenbach Subformation as probably representing the type stratum.
Material examined. Specimens NHMMZ PE 1995/7735-LS, NHMMZ PE 1995/7748-LS, NHMMZ PE 1995/7778-LS, NHMMZ PE 2011/5205-LS, NHMMZ PE 2018/5002-LS_2, NHMMZ PE 2018/5002-LS_3, NHMMZ PE 2021/5001-LS, and NHMMZ PE 2022/5002-LS from the Niedermoschel black shale east of Niedermoschel at the road to Alsenz, and NHMMZ PE2021/5002-LS from the Niedermoschel black shale at Mannweiler-Cölln (see Poschmann et al. 2021 for locality data); UGKU 3167 (formerly Raisch collection SRK SF_54) from a slightly older lake deposit within the Jeckenbach Subformation at Sitters (see Poschmann and Schindler 2004 for locality data); UGKU 3166 (formerly Raisch collection SRK Gb_687) and UGKU 3165 (formerly Raisch collection SRK Gb_1) from the Humberg black shale (Odernheim Subformation) near Freisen-Grügelborn/Saarland (see Poschmann and Schindler 2004, 2023 for locality data).
Description. Forewing with slightly convex costal margin and rounded apex, usually with striking colouration pattern preserved; colouration pattern basically consisting of widened coloured areas surrounding crossveins, sometimes fused into larger spots or irregular areas approaching transverse stripes, apical fourth to third of forewing usually more uniformly coloured; hind wing more uniformally coloured, with uncoloured/colour pattern mainly restricted to anterior margin; forewing with crossveins often preserved, up to about 10 simple crossveins in costal field, which may branch and form a simple network; up to eight crossveins were observed between ScP and RA, up to four between RA and RP, up to five between RP and MA, up to five between MA and MP, up to five between MP and CuA1, up to four between CuA1 and CuA2, and up to six between CuA2 and CuP. Lengths of wings 10.1–11.6 mm, widths 3.7–4.0 mm, length/width ratios 2.73–3.10. ScP straight to slightly sigmoidal, apically deflected, terminating in costal margin 2.2–2.8 mm from wing apex; area between costal margin and ScP only slightly broader than area between ScP and R at about one quarter of wing length; R slightly sigmoidal, RA and RP separating at about one third of wing length; RA simple, almost straight, showing in some specimens a shallow hook-like termination, ending on anterior margin 0.8–1.5 mm from wing apex; width of area between RA and RP irregular, widest at apical third to fourth of wing; RP bifurcating at about apical third of wing, ending with two branches at or above wing apex; M divided into MA and MP well basal of separation of RA and RP near anastomosis (if present) or fused basal part of M with CuA; MA simple, almost straight, ending below wing apex; MP with basal part weakly sclerotized, bifurcated at or slightly apical to bifurcation of CuA1, with the two branches ending on posterior wing margin; MP area rather narrow; CuA with two basal branches, CuA1 branched; CuA2 simple and almost straight; M and CuA1 basally fused, anastomosed, or at least very close; CuP and anal area poorly preserved.
Remarks. Specimen UGKU 3166 is morphologically close to the specimens from the Jeckenbach Subformation, but differs in a more strongly curved anterior forewing margin, a shorter distance between the origin of the RP fork and the origin of the MP fork, in the posterior branch of CuA1 showing a short fork, in having the bases of M and CuA not fused or anastomosed, and in its small size. UGKU 3165 from the same locality is larger and has CuA1 with two branches, but agrees with UGKU 3166 in having a short distance between the origin of the RP fork and the origin of the MP fork, and in having the bases of M and CuA not fused or anastomosed. The coloration pattern in these specimens significantly differs, but is interpreted as intraspecific variability as it is different among all specimens, unlike in extant Panorpa spp. that show similar patterns of coloration, but which are stable within the different species (Tillier 2008). CuA1 with three branches occasionally occurs also in Pictoborella clara (Cui et al. 2022: Fig. 4). For the time being, the material base to distinguish the Grügelborn Pictoborella specimens from those previously described from the Jeckenbach Subformation is too weak to justify the erection of a distinct taxon, i.e. subspecies.

Pictoborella germanica (Prokop et al., 2012), Niedermoschel black shale; photographs and explanatory drawings, respectively. a Forewing NHMMZ PE 2021/5002-LSa, Mannweiler-Cölln; b hind wing NHMMZ PE 2018/5002-LS (specimen 1), Niedermoschel; c hind wing NHMMZ PE 2018/5002-LS (specimen 2), Niedermoschel; d supposed hind wing NHMMZ PE 2011/5205-LSa (flipped horizontally), Niedermoschel. (Scale bars = 5 mm)
Genus Oborella Kukalová, 1964
Type species. Oborella matura Kukalová, 1964; early Permian of Moravia (Czech Republic).
Diagnosis. See Kukalová (1964)
Oborella brauckmanni Prokop et al., 2012
Figure 5

Oborella brauckmanni Prokop et al., 2012, Niedermoschel black shale at Niedermoschel; photographs and explanatory drawings, respectively. a Forewing NHMMZ PE 1995/9647-LS (flipped horizontally); b forewing NHMMZ PE 1995/9656-LS; c forewing NHMMZ PE 2021/5014-LS; d forewing NHMMZ PE 2011/5201-LS; e ?hind wing NHMMZ PE 1995/9642-LS (flipped horizontally). (Scale bars = 5 mm)
Diagnosis. Forewings of medium size, slightly broadened in distal half. Anterior margin almost straight; crossveins in costal area simple, short and in one row; ScP terminating in C at 2/3 wing length; subcostal area about as wide as costal area or slightly wider; RP simply forked, originating slightly before or at middle of wing; stems of M and CuA fused for a rather long distance; MA simple; MP originating at about 1/4 wing length, simply forked; CuA with two main branches, both simple; hind wings essentially the same but slightly broader (modified from Prokop et al. 2012).
Type locality and type stratum. Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation.
Remark. Concerning type locality and type stratum the same applies as above stated for P. germanica.
Material examined. Specimens NHMMZ PE 1995/9642-LS, NHMMZ PE 1995/9647-LS, NHMMZ PE 1995/9656-LS, NHMMZ PE 2011/5201-LS, NHMMZ PE 2021/5014-LS. All from the Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Description. Forewing with slightly convex costal margin and rounded apex lacking any colouration pattern, i.e. uniformly coloured, except for the possible presence of small uncoloured spots in the costo-apical margin of some specimens; crossveins may or may not be preserved, judging from wings studied here, there can be up to 10 simple crossveins in costal area, up to three observed between ScP and RA, up to two between RA and RP, and also between RP and MA, up to three between MA and MP, two or three between MP and CuA1, up to three between CuA2 and CuP. Lengths of wings 8.3–9.2 mm, widths 2.6–3.0 mm, length/width ratios 2.77–3.31. ScP straight to slightly sigmoidal, apically deflected but reaching costal margin 2.7–3.2 mm from wing apex; area between costal margin and ScP moderately broad in midwing; R nearly straight, RA and RP separating slightly basal to or at middle of wing; RA mostly simple, in one case with terminal fork, nearly straight, and ending with hook-like termination on anterior margin 1.2–1.5 mm from wing apex; RA and RP area irregular widest at about midlength; RP bifurcated in distal third of wing, ending with two branches just above wing apex; M divided into MA and MP about 1,4–1.8 mm basal of separation of RA and RP; MA simple, almost straight, ending slightly below wing apex; area between RP and MA with few cross-veins; MP with basal part weakly sclerotized, bifurcated with the two branches ending on posterior wing margin; MP area rather narrow; CuA with two basal branches, both simple and almost straight; CuP and anal area poorly preserved.
Remarks. This taxon is similar in venation pattern to Oborella Kukalová, 1964 and Blania Kukalová, 1964, the latter having been synonymized with Oborella by Prokop et al. (2012). It differs from other species in these genera, and especially from the type species O. matura Kukalová, 1964 by consistently more slender forewings with RP simply forked (versus ending in three or more branches, except in Blania rotunda Kukalová, 1964) and simple CuA1 (versus forked).
Oborella moschelensis sp. nov.
Figure 6a

Various grylloblattids assigned to genus Oborella, Niedermoschel black shale at Niedermoschel; photographs and explanatory drawings, respectively. a Oborella moschelensis sp. nov., holotype, forewing NHMMZ PE 2022/5001-LS (flipped horizontally); b Oborella sp., probably forewing NHMMZ PE 2021/5000-LS (flipped horizontally); c Oborella monsjovisensis Poschmann and Nel 2020, holotype, forewing NHMMZ PE 2020/5003-LS (flipped horizontally); d ?Oborella sp., complete insect NHMMZ PE 2021/5012-LS from Rockenhausen, Hopbachtal (flipped horizontally). (Scale bars = 5 mm)
Zoobank LSID urn:lsid:zoobank.org:act:D7C50118-1D2D-4BA3-AB08-A7EED22C0B5B.
Etymology. Named after the Moschel, a small creek near the type locality.
Holotype. Forewing NHMMZ PE 2022/5001-LS, deposited in the Landessammlung für Naturkunde Rheinland-Pfalz at the Naturhistorisches Museum Mainz (NHMMZ).
Type locality and type stratum. Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation, Jeckenbach Subformation, Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Diagnosis. Crossveins in forewing costal area simple, short and in one row; RP diverging from RA in the basal 1/4 of wing length, just distad point of separation between CuA and M; MA simple; MP forked; CuA with two main branches, CuA1 with four branches, CuA2 simple; forewing uniformly coloured except along crossveins.
Description. Well-preserved forewing lacking basal part, with almost straight costal margin and rounded apex, preserved length 6.9 mm (estimated complete length about 8 mm), width 3.0 mm; ScP not well-preserved, probably ending in C about 2/3 of wing length; R almost straight; RP diverging from RA well before 1/4 of wing length; simple RA ending at 3/4 of wing length; RP forked two times, ending in three branches on posterior wing margin, posterior-most branch ending close to wing apex; stems of M and CuA fused, M diverging from CuA slightly basad of divergence of RP and RA; MA simple, ending below wing apex; MP branched slightly before midwing, ending in two branches; CuA1 branched, ending in two short forks (four branches) on posterior wing margin; CuA2 and probably CuP both simple; anal area not preserved; probably more than six simple crossveins in coastal area, about five crossveins preserved between ScP and RA, up to seven between RA and RP, forming an irregular network, three between RP2 and RP3, six between RP and MA, seven between MA and MP, at least one in the MP fork, five between MP and CuA1, at least five between CuA1 and CuA2, and at least two between CuA2 and CuP; forewing uniformly coloured, except along crossveins.
Discussion. The venation pattern in this species is again close to Pictoborella and other Oborella species. It differs from these taxa in the base of RP just distad the point of separation of CuA1 from M. It also differs from other Oborella (and Blania) species in a straighter RA and resulting space between RA and RP showing a constant widening. Furthermore, it differs in its simple MA, except from O. rusticana Kukalová, 1964 and O. matura Kukalová, 1964, which share this character. Oborella moschelensis sp. nov. differs from the latter species in CuA1 ending in four branches (versus two or three). It also differs from Pictoborella in the colouration pattern.
Oborella sp.
Figure 6b
Material. NHMMZ PE 2021/5000-LS, from the Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Remarks. NHMMZ PE 2021/5000-LS is closely approaching the size and venation characters described for P. germanica, but differs in ScP apparently ending in RA, termination of RA being very close to termination of anterior branch of RP, and in colouration, restricted to immediate vicinity of veins.
Oborella monsjovisensis Poschmann and Nel, 2020
Figure 6c
Holotype. Forewing NHMMZ PE 2020/5003-LS, deposited in the Naturhistorisches Museum Mainz/Landessammlung für Naturkunde Rheinland-Pfalz.
Diagnosis. Forewing characters only. 14 crossveins in costal area; RA simple, terminating close to wing apex; RP, and MP simply forked; MA simple or forked; CuA with two main branches, anterior one CuA1 forked and posterior one CuA2 simple; M and CuA very close or anastomosed near wing base (emended from Poschmann and Nel 2020).
Type horizon and locality. Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation, Jeckenbach Subformation, Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Remarks. The species mainly stands apart from the other lemmatophorid taxa described herein in the large forewing size approaching a length of 15 mm, a large distance between the terminations of ScP and RA, and MA being branched (see below).
The supposed Liomopteridae Liomopterites germanicus Brauckmann and Willmann, 1990 (regarded as possibly belonging to Liomopterum by Cui et al. 2022) is rather similar to Oborella monsjovisensis but lacks the basal part of the wing. L. germanicus has more crossveins and its anterior branch of RP has a distal fork in contrast to the simple one in O. monsjovisensis. However, these two taxa are very similar and probably belong to the same genus. It is suggested here that L. germanicus is possibly a lemmatophorid close to Oborella. Nevertheless, the lack of information on the potential basal fusion of CuA with M forbids us to exclude affinities with the Liomopteridae.
?Oborella sp.
Figure 6d
Material. NHMMZ PE 2021/5012-LS from the Morsbacher Hof black shale (informal stratigraphic unit M5 after Boy et al. 2012), from the Hopbach Valley north of Rockenhausen.
Description. An almost complete insect, preserved length about 13.5 mm, but with structures difficult to tell apart. ?Forewing length about 9.8 mm; ScP long, ending in C; origin of RP slightly more apical than MA/MP fork; RA and RP simply forked; MA simple, MP simply forked; base of M probably fused with CuA; CuA1 with three branches; CuA2 simple. Head probably prognathous; legs strong; hind leg tarsus probably with five segments.
Remarks. Wing venation largely agrees with Pictoborella and Oborella monsjovisensis. The shorter wing, longer ScP, forked RA, and simple MA more readily agree with Pictoborella. But prognathous head and strongly built tarsus differ from P. clara (cf., Cui et al. 2022: Fig. 2) and from P. germanica (specimen NHMMZ PE 2022/5002-LS; Fig. 7). However, this specimen is left here in open nomenclature as ?Oborella sp.

Pictoborella germanica (Prokop et al., 2012), Niedermoschel black shale of Niedermoschel. Almost complete, but poorly preserved specimen, NHMMZ PE 2022/5002-LS, photograph and explanatory drawing, respectively. (Scale bars = 5 mm)
Genus Artinska Sellards, 1909
Type species. Artinska clara Sellards, 1909; early Permian (Artinskian) of Kansas/USA.
Diagnosis. Forewing characters only. Forewing broad, costal margin arched and apex rounded; ScP terminating well beyond middle of wing; R and RA straight as far as pterostigmal region, where there is a slight curvature; RP arising proximad middle of wing, with at least one deep fork and occasionally a small fork on one of the branches; M free from R at base; MA usually arising slightly proximad origin of RP, either free from RP or coalesced with it, and being either simple, forked, or three-branched; MP either simple, two-branched, or three-branched; CuA with characteristic form in family, with from two to four branches; CuP unbranched and weakly formed; anal veins present, both very variable in form (modified from Carpenter 1935).
Artinska glanensis sp. nov.

Artinska glanensis sp. nov., Niedermoschel black shale at Niedermoschel, Holotype specimen NHMMZ PE 2018/5001-LS; photographs and explanatory drawings, respectively. a Left forewing (flipped horizontally); b almost complete insect fossil; c poorly preserved head and anterior thoracal region including basal part of right antenna (An); d explanatory drawing showing left and right forewing. (Scale bars = 5 mm (a, b, and d), 1 mm (c))

Artinska glanensis sp. nov., Niedermoschel black shale at Niedermoschel; photographs and explanatory drawings, respectively. a Paratype, forewing part NHMMZ PE 2021/5011-LSa (flipped horizontally); b paratype, forewing counterpart NHMMZ PE 2021/5011-LSb; c paratype, part of forewing NHMMZ PE 2022/5000-LS. (Scale bars = 5 mm)
Zoobank LSID urn:lsid:zoobank.org:act:4D0E0E7F-864A-46B1-86D6-F6F147E4409F.
1999 Artinska sp.–Hörnschemeyer: 58, fig. 4
2007 Artinska sp.–Brauckmann: 182, fig. 8
Etymology. Named after the Glan, a creek west of the type region.
Holotype. NHMMZ PE 2018/5001-LS, deposited in the Landessammlung für Naturkunde Rheinland-Pfalz at the Naturhistorisches Museum Mainz (NHMMZ).
Paratypes. NHMMZ PE 2021/5011-LS, and NHMMZ PE 2022/5000-LS, topo- and stratotypic.
Type locality and type stratum. Saar-Nahe Basin, Rheinland-Pfalz, Germany. Early Permian, Rotliegend, Meisenheim Formation, Jeckenbach Subformation, Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Diagnosis. Forewing rather small, length between 7.7 and 8.1 mm; fork of R into RA and RP distinctly distad first fork of M; RA simple, very weakly S-shaped; RP branched, with apex anteriad wing apex; RP and MA well separated; ScP elongate, reaching distal third of wing.
Description. Forewing with slightly convex coastal margin and rounded apex, no obvious colouration pattern preserved; crossveins in coastal area poorly preserved or almost lacking, up to about seven simple crossveins were observed between ScP and RA, six between RA and RP, eight between RP and MA, four between MA and MP, six between MP and CuA1, four between CuA1 and CuA2, and seven between CuA2 and CuP. Lengths of wings 7.7–8.1 mm, widths 2.8–3.0 mm, length/width ratios 2.70–2.75. ScP almost straight, terminating in costal margin 2.1–2.4 mm from wing apex; area between costal margin and ScP approximately as broad as area between ScP and R; R slightly sigmoidal, RA and RP separating just about one third of wing length; RA simple, slightly sigmoidal, termination slightly bent towards RP, ending on anterior margin 0.7–1.0 mm from wing apex; RA and RP area irregular, widest at about apical third of wing; RP bifurcating at about apical third of wing, ending with two branches above wing apex; M divided into MA and MP well basal of separation of RA and RP 1.8–2.1 mm from wing base; MA simple or forked apical to branching of RP, almost straight, ending near wing apex; MP with basal part weakly sclerotized, bifurcated clearly apical to bifurcation of CuA1, with two branches ending on posterior wing margin; MP area rather narrow; CuA with two basal branches, CuA1 branched one or two times; CuA2 simple and almost straight; bases of M and CuA1 separated, connected by a crossvein, perhaps with very short anastomosis in NHMMZ PE 2018/5001-LS; CuP straight, simple or possibly forked in NHMMZ PE 2018/5001-LS; PCu simple; anal area poorly preserved.
Discussion. This new fossil falls in the Lemmatophoridae (sensu Storozhenko 1998) because it has the diagnostic characters of the family: forewing small or medium in size, narrow, covered with hairs (unknown for this fossil), membranous, with a rounded apex. ScP ending into C at apex or in apical third of wing; costal area narrow, crossed by simple straight branches of ScP; R simple; RP branching off R in basal third or in middle of wing, simple or with two branches; radial area moderately widened; M divided into MA and MR just proximal or distal to base of RP; MA simple or with a fork, often forming an anastomosis with RP; MP with 2–3 branches, desclerotized in middle; CuA divided into branching CuA1 and simple CuA2; anastomosis between M and CuA bases quite often expressed; CuP simple. Anal area narrow; A1 and A2 simple; all crossveins simple, straight, sparse (after Storozhenko 1998, translated from Russian). The moderately elongate wing supports an attribution to the Lemmatophorinae. The base of RP situated in basal third of the wing, the narrow costal area, and the forked RP make it fall in the genus Artinska (Storozhenko 1998).
Three species of Artinska were described from the Artinskian Wellington Formation of North America. A. sellardsi Tillyard, 1928 is larger (forewing length 10.5–12 mm) than Artinska glanensis sp. nov. and has the first fork of R at approximately the same position as the first fork of M in relation to wing length. This also applies to A. ovata Sellards. 1909, which additionally has RA much more strongly S-shaped than A. glanensis sp. nov. (cf. Tillyard 1928; Carpenter 1935; Beckemeyer 2009). A. clara Sellards, 1909 is larger (forewing length 8.5–11.5 mm) than the new species and its RP terminates in a more posterior position near the wing apex (Tillyard 1928; Carpenter 1935). A. larisae Novokshonov, 1999 from the Kungurian of Tshekarda again has RA much more strongly S-shaped than A. glanensis sp. nov., and a conspicuous ‘junction’ of RP and MA, but the latter may be an individual aberration (Novokshonov 1999). Artinska infigurabilis Aristov, 2004 from the same locality has a relatively short ScP just reaching the second half of the forewing and CuA1 is simple or bifurcating. Artinska sojanensis Aristov, 2009 from the Kazanian of Sojana/Russia is larger (forewing length 8.5–11.5 mm) and has RA branched (Aristov 2009). A. vorkutensis Aristov, 2005 in Rasnitsyn et al. (2005) from the mid- to late Permian Inta Formation of the Pechora Basin has RA much more strongly S-shaped and RP unbranched (Rasnitsyn et al. 2005).
Family Liomopteridae Sellards, 1909
Genus and species inc. sed.
Figure 10

Undetermined Liomopteridae, Niedermoschel, incomplete forewing NHMMZ PE 2021/5013-LS; photograph and explanatory drawing, respectively. (Scale bars = 5 mm)
Material. Forewing NHMMZ PE 2021/5013-LS from the Niedermoschel black shale, east of Niedermoschel at the road to Alsenz (see Poschmann et al. 2021 for locality data).
Description. Forewing with slightly convex costal margin. Preserved part of wing about 17.6 mm long and 5.3 mm wide. Costal area narrow, with about seven crossveins visible, many more certainly present distally but not clearly preserved. ScP long, ending on C in apical fourth of wing. Area between ScP and R about as wide as costal area, with undetermined number of crossveins. RA ending near wing apex. RP diverging from RA at about one fourth of wing; RA-RP area widest at about apical fourth of wing, near apical fork of RP. The unusual venation pattern in cubital area may be an aberrant feature due to secondary fusions of veins.
Remarks. Despite its incompleteness, a tentative attribution of this forewing to the Liomopteridae is supported by the following diagnostic characters (Storozhenko 1998, translated from Russian): forewing rather small, medium size, hairless, membranous, without pattern or light, with large dark spots and stripes; with a broadly or narrowly rounded apex; ScP ending into C at apex or in apical half of wing; costal area narrow, crossed by simple straight crossveins; RA simple; RP branching off R in basal third of wing, with two branches, directed towards wing apex; radial area moderately widened; M divides into MA and MP, slightly proximal to base of RP; MA with 2–3 branches; no anastomosis between M and CuA. Unfortunately, this wing lacks too many characters to be attributed to a precise genus.
Conclusion
The Lower Permian Meisenheim Formation (Asselian to possibly early Sakmarian) hitherto yielded about 40 described insect species from 11 different orders (Table 1). These originate from sediments of intramontane lakes, and especially from lake phases showing a deltaic influence. A slight contact metamorphism of the host sediment strongly facilitates the detection of insect fossils and is an additional factor contributing to the rich insect record of some localities, such as Niedermoschel, Mannweiler-Cölln, and Freisen-Grügelborn. The relative abundance of higher insect taxa of these sites strongly differs from those seen in Pennsylvanian (‘Westfalian’ and Gzhelian) deposits of the Saar-Nahe Basin (and elsewhere). In the latter, Palaeodictyoptera and especially Dictyoptera strongly dominate in terms of both taxonomic richness and abundance (see Brauckmann et al. 2007). In contrast, in insect deposits of the Meisenheim Formation Paoliida (Blattinopsidae), Archaeorthoptera, and Grylloblattida are remarkably diverse and/or abundant. Acercaria sensu lato are an important component with Miomoptera (Palaeomanteidae) being more common than Dictyoptera, as exemplified by the Niedermoschel assemblage (Hörnschemeyer 1999; Poschmann et al. 2021). Thus, the composition of the insect assemblages of the Meisenheim Formation reveals a decidedly Permian character and stands in marked contrast to insect assemblages hitherto known from Pennsylvanian sites of the Saar-Nahe Basin. This agrees well with general patterns of insect diversity changes from the Pennsylvanian to early Permian (cf., e.g., Rasnitsyn et al. 2004, 2015).
Data availability
There are no additional data.
References
Aristov, D.S. 2004. The fauna of grylloblattid insects (Grylloblattida) of the Lower Permian locality of Tshekarda. Paleontological Journal 38 (suppl. 2): S80–S145.
Aristov, D.S. 2009. New grylloblattids of the family Lemmatophoridae (Insecta: Grylloblattida) from the Permian of Russia. Paleontological Journal 43: 272–276.
Aristov, D.S. 2010. Review of grylloblattid insects (Insecta: Grylloblattida) from the Solikamsk deposits of the Perm region. Paleontological Journal 44: 505–514.
Aristov, D.S., R. Prevec, and M.B. Mostovski. 2009. New and poorly known grylloblattids (Insecta: Grylloblattida) from the Lopingian of the Lebombo Basin, South Africa. African Invertebrates 50: 279–286.
Beckemeyer, R.J. 2009. Artinska ovata (Sellards) 1909 and Paraprisca fragilis (Sellards) 1909 (Insecta: Polyneoptera: Lemmatophoridae) newly reported from the Lower Permian of Noble County, Oklahoma, with notes on Wellington Formation Lemmatophoridae. Transactions of the Kansas Academy of Science 112: 45–56.
Beckemeyer, R.J., and J.D. Hall. 2007. The entomofauna of the Lower Permian fossil insect beds of Kansas and Oklahoma USA. African Invertebrates 48: 23–39.
Béthoux, O., and M. Poschmann. 2009. A new lobeattid insect from the Permo-Carboniferous of Niedermoschel, southwestern Germany (Archaeorthoptera). Journal of Orthoptera Research 18: 139–143. https://doi.org/10.1665/034.018.0201.
Boy, J.A., J. Haneke, G. Kowalczyk, V. Lorenz, T. Schindler, H. Stollhofen, and H. Thum, 2012. Rotliegend im Saar-Nahe-Becken, am Taunus-Südrand und im nördlichen Oberrheingraben. In Stratigraphie von Deutschland X. Rotliegend. Teil I: Innervariscische Becken, Deutsche Stratigraphische Kommission, eds. Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften 61: 254–377.
Brauckmann, C. 2007. Die Insekten im Permokarbon des Saar-Nahe-Beckens. In Kohlesümpfe, Seen und Halbwüsten. Dokumente einer rund 300 Millionen Jahre alten Lebewelt zwischen Saarbrücken und Mainz, Schindler, T., and U.H.J. Heidtke, eds. POLLICHIA Sonderveröffentlichung 10: 170–196. Bad Dürkheim.
Brauckmann, C., and R. Willmann. 1990. Insekten aus dem Permo-Silesium der Bohrung Weiterstadt 1 (Blattodea, „Protorthoptera”; Oberrheinische Tiefebene, SW-Deutschland). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 1990 (8): 470–478.
Brauckmann, C., M. Poschmann, and T. Schindler. 2007. Verzeichnis der Arten. In Kohlesümpfe, Seen und Halbwüsten. Dokumente einer rund 300 Millionen Jahre alten Lebewelt zwischen Saarbrücken und Mainz, Schindler, T., and U.H.J. Heidtke, eds. POLLICHIA Sonderveröffentlichung 10: 11 p. on supplemental CD. Bad Dürkheim.
Burckhardt, D., A. Nel, M. Raisch, and M.J. Poschmann. 2023. A new putative moss bug (Insecta: Hemiptera) from the lower Permian of the Saar-Nahe Basin, SW Germany, and the age of Coleorrhyncha. Historical Biology 35 (6): 832–837. https://doi.org/10.1080/08912963.2022.2067759.
Carpenter, F.M. 1935. The Lower Permian insects of Kansas. Part 7. The order Protoperlaria. Proceedings of the American Academy of Arts and Sciences 70: 103–146. https://doi.org/10.2307/20023126.
Cui, Y., C. Shih, and D. Ren. 2019. Notoptera – Rock Crawlers and Ice Crawlers. In Rhythms of Insect Evolution, ed. D. Ren, C. Shih, T. Gao, Y. Wang, and Y. Yao, 137–147. Chichester, UK: Wiley Blackwell.
Cui, Y., S. Brauner, J.W. Schneider, and O. Béthoux. 2022. Grylloblattidan insects from Sperbersbach and Cabarz (Germany), two new early Permian and insect-rich localities. Journal of Paleontology 96: 355–374. https://doi.org/10.1017/jpa.2021.101. (first published online 2021).
Guthörl, P. 1934. Die Arthropoden aus dem Carbon und Perm des Saar-Nahe-Pfalz-Gebietes. Abhandlungen der Preußischen Geologischen Landesanstalt, Neue Folge 164: 1–219.
Guthörl, P. 1964. Obituary, with a compilation of his publications. Annales Universitatis Saraviensis 4: 1–9.
Hörnschemeyer, T. 1999. Fossil insects from the Lower Permian of Nierdermoschel (Germany). AMBA projects AM/PFICM98/1.99: Proceedings of the First Palaeoentomological Conference Moscow 1998: 57–59.
Hörnschemeyer, T., and H. Stapf. 2001. Review of Blattinopsidae (Protorthoptera) with description of new species from the Lower Permian of Niedermoschel (Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 221: 81–109. https://doi.org/10.1127/njgpa/221/2001/81.
Kinzelbach, R., and H. Lutz. 1984. Eine neue Eintagsfliege Misthodotes stapfi n. sp. aus dem Rotliegenden des Nahe-Gebietes (Ephemeroptera: Permoplecoptera: Misthodotidae). Paläontologische Zeitschrift 58: 247–253. https://doi.org/10.1007/BF02986063.
Kirejtshuk, A.G., M. Poschmann, J. Prokop, R. Garrouste, and A. Nel. 2014. Evolution of the elytral venation and structural adaptations in the oldest Palaeozoic beetles (Insecta: Coleoptera: Tshekardocoleidae). Journal of Systematic Palaeontology 12 (5): 575–600. https://doi.org/10.1080/14772019.2013.821530.
Kukalová, J. 1964. Permian insects of Moravia. Part 2. Liomopteridea. Sborník Geologických Věd, Paleontologie 3: 39–118.
Kukalová-Peck, J. 1991. Fossil history and the evolution of hexapod structures, 141–179. In The insects of Australia. A textbook for students and research workers, 2nd edition, ed. CSIRO. Carlton: Melbourne University Press
Menning, M., J. Glodny, J. Boy, R. Gast, G. Kowalczyk, T. Martens, R. Rößler, T. Schindler, V. von Seckendorf, and S. Voigt. 2022. The Rotliegend in the Stratigraphic Table of Germany 2016 (STG 2016). Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 173: 3–139. https://doi.org/10.1127/zdgg/2022/0311.
Myskowiak, J., J.M. Pouillon, and A. Nel. 2015. The oldest record of the grylloblattodean family Euryptilonidae discovered in the Late Carboniferous of France. Comptes Rendus Palevol 14: 1–4. https://doi.org/10.1016/j.crpv.2014.08.002.
Nel, A., and M.J. Poschmann. 2020. A new stem Orthoptera (Archaeorthoptera: Oedischioidea) from the Early Permian of the Saar-Nahe Basin, southwest Germany. Geobios 63: 47–52. https://doi.org/10.1016/j.geobios.2020.10.001.
Nel, A., and M.J. Poschmann. 2021a. Comparison of the recently described early Permian paoliid genus Permomertovia with the eoblattid genera of the families Permulidae and Permotermopsidae. Palaeoentomology. 4: 39–43. https://doi.org/10.11646/palaeoentomology.4.1.8.
Nel, A., and M.J. Poschmann. 2021b. Early Permian acercarian insects (Miomoptera, Palaeomanteidae) from the Saar-Nahe Basin (Rhineland-Palatinate, SW Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 300: 267–277. https://doi.org/10.1127/njgpa/2021/0989.
Nel, A., and M.J. Poschmann. 2021c. A new archaeorthopteran insect from the Early Permian of the Saar-Nahe Basin, SW Germany. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 301: 79–84. https://doi.org/10.1127/njgpa/2021/0998.
Nel, A., and M.J. Poschmann. 2021d. A new representative of the ‘orthopteroid’ family Cnemidolestidae (Archaeorthoptera) from the Early Permian of Germany. Acta Palaeontologica Polonica 66: 641–646. https://doi.org/10.4202/app.00879.2021.
Novokshonov, V. 1999. Eine neue Notoptere aus dem unteren Perm des Urals (Insecta: Grylloblattida, Lemmatophoridae). Neues Jahrbuch für Geologie und Paläontologie Monatshefte 1999: 641–646.
Perez-de la Fuente, R., A. Nel, and M.J. Poschmann. 2022. Shedding light on the enigmatic extinct insect order Glosselytrodea: new diversity and key morphological insights on non-tegminized Permian forms. Papers in Palaeontology. https://doi.org/10.1002/spp2.1452.
Poschmann, M.J., and A. Nel. 2020. New Early Permian insects (Insecta: Paoliida and Grylloblattida) from the Niedermoschel black shale of the Saar-Nahe Basin, SW Germany. Palaeoentomology 3: 623–631. https://doi.org/10.11646/palaeoentomology.3.6.14.
Poschmann, M.J., and A. Nel. 2021. The first Permian scorpionfly from Germany (Insecta, Panorpida: Protomeropidae). Palaeoentomology 4: 231–236. https://doi.org/10.11646/palaeoentomology.4.3.10.
Poschmann, M., and T. Schindler. 2004. Sitters and Grügelborn, two important Fossil-Lagerstaetten in the Rotliegend (?Late Carboniferous – Early Permian) of the Saar-Nahe Basin (SW-Germany), with the description of a new palaeoniscoid (Osteichthyes, Actinopterygii). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 232 (2–3): 283–314. https://doi.org/10.1127/njgpa/232/2004/283.
Poschmann, M.J., and T. Schindler. 2023. Egg deposition on carapaces of the giant clam shrimp Palaeolimnadiopsis (Crustacea: Spinicaudata) from the Early Permian of the Saar-Nahe Basin, SW-Germany. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 307 (1): 81–89. https://doi.org/10.1127/njgpa/2023/1114.
Poschmann, M., A. Nel, and T. Schindler. 2019. Erster Nachweis von Riesen-Urlibellen (Odonatoptera: Meganisoptera: Meganeuridae) im Permokarbon des Saar-Nahe-Beckens (Rheinland-Pfalz, SW-Deutschland). Mainzer Naturwissenschaftliches Archiv 56: 71–86.
Poschmann, M.J., A. Nel, T. Schindler, and D. Uhl. 2021. Die Niedermoschel-Bank, ein bedeutender Fundhorizont mit einer hoch-diversen Insekten- und Pflanzenassoziation im Rotliegend (Unterperm) des Saar-Nahe-Beckens (Rheinland-Pfalz, SW-Deutschland). Mainzer Naturwissenschaftliches Archiv 58: 17–40.
Poschmann, M.J., A. Nel, and M. Raisch. 2023. A new archaeorthopteran insect from the Early Permian of the Saar-Nahe Basin, SW Germany, and the earliest record of Elcanoidea (Orthoptera). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 308 (2): 181–187.
Pretorius, A.I., C.C. Labandeira, A. Nel, and R. Prevec. 2021. Latest Permian insects from Wapadsberg Pass, southern Karoo Basin, South Africa. Austral Entomology 60: 560–570. https://doi.org/10.1111/aen.12540.
Prokop, J., and J. Kukalová-Peck. 2017. New insects from the earliest Permian of Carrizo Arroyo (New Mexico, USA) bridging the gap between the Carboniferous and Permian entomofaunas. Insect Systematics & Evolution 48: 493–511. https://doi.org/10.1163/1876312X-48022160.
Prokop, J., K.-D. Weiß, R.-P. Dechambre, and A. Nel. 2012. Early Permian insects from Saar-Nahe Basin of Odernheim town site, Rheinland-Pfalz in Germany (Insecta: Grylloblattida, Blattinopseida). Geodiversitas 34: 271–281. https://doi.org/10.5252/g2012n2a2.
Rasnitsyn, A.P., D.S. Aristov, A.V. Gorochov, J.M. Rowland, and N.D. Sinitshenkova. 2004. Important new insect fossils from Carrizo Arroyo and the Permo-Carboniferous faunal boundary. New Mexico Museum of Natural History and Science Bulletin 25: 215–246.
Rasnitsyn, A.P., I.D. Sukacheva, and D.S. Aristov. 2005. Permian insects of the Vorkuta Group in the Pechora Basin, and their stratigraphic implications. Paleontological Journal 39: 404–416.
Rasnitsyn, A.P., D.S. Aristov, and D.A. Rasnitsyn. 2015. Dynamics of insect diversity during the Early and Middle Permian. Paleontological Journal 49: 1282–1309.
Schindler, T., and U.H.J. Heidtke, eds. 2007. Kohlesümpfe, Seen und Halbwüsten. Bad Dürkheim: POLLICHIA Sonderveröffentlichung 10.
Schneider, J. 1978. Zur Taxonomie und Biostratigraphie der Blattodea (Insecta) des Karbon und Perm der DDR. Freiberger Forschungsheft C340: 1–152.
Schneider, J.W. 1982. Entwurf einer biostratigraphischen Zonengliederung mittels der Spiloblattinidae (Blattodea, Insecta) für das kontinentale, euramerische Permokarbon. Freiberger Forschungsheft C375: 27–47.
Schneider, J. 1983. Die Blattodea (Insecta) des Paläozoikums. Teil 1: Systematik, Ökologie und Biostratigraphie. Freiberger Forschungsheft C382: 106–145.
Schubnel, T., L. Desutter-Grandcolas, F. Legendre, J. Prokop, A. Mazurier, R. Garrouste, P. Grandcolas, and A. Nel. 2020. To be or not to be: Postcubital vein in insects revealed by microtomography. Systematic Entomology 45: 327–336. https://doi.org/10.1111/syen.12399.
Sellards, E.H. 1909. Types of Permian insects. Part III. Megasecoptera, Oryctoblattinidae and Protorthoptera. American Journal of Science (series 5) 27: 151–173.
Storozhenko, S.Yu. 1997. Classification of order Grylloblattida, with description of new taxa. Far Eastern Entomologist 42: 1–20.
Storozhenko, S.Yu. 1998. Systematics, phylogeny and evolution of the grylloblattids (Insecta: Grylloblattida), 1–207. Vladivostok: Dal'nauka. [in Russian]
Tillier, P. 2008. Contribution à l’étude des Mécoptères de France. Deuxième partie: Clé de détermination des Panorpa de France (Mecoptera Panorpidae). L’entomologiste 64: 21–30.
Tillyard, R.J. 1928. Kansas Permian Insects. Part 11. Order Protoperlaria: Family Lemmatophoridae (continued). American Journal of Science (ser. 5) 16: 313–348. https://doi.org/10.2475/ajs.s5-16.94.313.
Vršanský, P., S.Y. Storozhenko, C.C. Labandeira, and P. Ihringova. 2001. Galloisiana olgae sp. nov. (Grylloblattodea: Grylloblattidae) and the paleobiology of a relict order of insects. Annals of the Entomological Society of America 94: 179–184. https://doi.org/10.1603/0013-8746(2001)094[0179:GOSNGG]2.0.CO;2.
Walker, E.M. 1914. A new species of Orthoptera, forming a new genus and family. The Canadian Entomologist 46: 93–99. https://doi.org/10.4039/Ent4693-3.
Zhang, Q., E.A. Jarzembowski, and B. Wang. 2022. Widespread grylloblattid insects after the end-Permian mass extinction. Frontiers in Earth Sciences 10: 853833. https://doi.org/10.3389/feart.2022.853833.
Acknowledgements
We thank Petra Schäfers, Kai Nungesser, Thomas Schindler, Sabine Glienke, and Tom Bizer (all GDKE RLP, Mainz) for joint fieldwork, curating material in their care and/or for providing specimens. Sebastian Voigt (GEOSKOP Thallichtenberg) provided repository data for material stored at the GEOSKOP. We are greatly indebted to two anonymous reviewers whose insightful comments helped to improve this contribution.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Joachim T. Haug.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Poschmann, M.J., Nel, A. & Raisch, M. Diversity and variability of grylloblattidan insects (Grylloblattida) from the early Permian Meisenheim Formation of the Saar-Nahe Basin (SW-Germany). PalZ 98, 67–84 (2024). https://doi.org/10.1007/s12542-023-00672-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-023-00672-8