
Abstract
New taxa of the order Reculida (Insecta: Gryllones): imagines of Chauliodites babiy sp. nov. (family Chaulioditidae), Stegopterum kabalum sp. nov., S. pashka sp. nov., Permoshurabia tshasha sp. nov., Permuliercula chimaera gen. et sp. nov. (family Geinitziidae), Khosara ultima sp. nov. (family Liomopteridae), and Permotoma aba gen. et sp. nov. (family Skaliciidae) and a nymph of Tarakanikha satura gen. et sp. nov. (Gryllones incertae ordinis) have been described from the Upper Permian Babii Kamen’ locality (the Vyatkian Stage of Russia). Description of Shurabia annosa Aristov, 2004 has been modified and the species has been assigned to the genus Geinitzia.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
The present article is a continuation of the work on the description of Gryllones insects from the Babii Kamen’ locality (Tarakanikha and Kedrovo beds of the Maltsevo Formation). This locality is situated in the Kemerovo Region of Russia and belongs to the Vyatkian layer of the Late Permian (Sadovnikov, 2016). According to other authors (Shcherbakov, 2008), Babii Kamen’ is dated to the Lower Triassic. In the first part of this work (Aristov, 2020), eoblattid insects were considered. Representatives of the order Reculida that belong to the superorder Blattidea of the infraclass Gryllones, and a nymph of an obscure taxonomic position that belongs to the same infraclass are considered below. In addition, the fauna of all Gryllones insects of the Babii Kamen’ locality is considered and compared with other Upper Permian and Lower and Middle Triassic faunas.
The infraclass Gryllones is represented by orthoptera-like and cockroach-like Gryllones insects in Babii Kamen’. The former include Mesoedischiidae orthopterans (Gorochov, 2005). Cockroach-like insects (superorder composition as in: Aristov, 2015a) are represented by eoblattids (Aristov, 2020) and their descendants: cockroaches (Blattida: Subioblattidae: Rasnitsyn et al., 2013), reculids (see below) and embiids (Embiida: Alexarasniidae: Shcherbakov, 2015).
Two genera and four species of the family Chaulioditidae (Aristov, 2003) and one genus with one species of the family Geinitziidae (Aristov, 2004) were identified in the reculid insect (Insecta: Reculida) fauna of Kedrovo Beds of Babii Kamen’ previously. Another species of Chaulioditidae, four species belonging to three genera of the family Geinitziidae, and single representatives of both Liomopteridae and Skaliciidae are described below. Of the families listed, Chaulioditidae and Geinitziidae are known from both Permian and Triassic, whereas Skaliciidae and Liomopteridae are known from the Permian only (Rasnitsyn et al., 2013). In addition, a nymph of an obscure position, Tarakanikha satura gen. et sp. nov., was described and attributed to the Gryllones insects; this insect was found in the Tarakanikha beds of the Maltsevo Formation. These beds are older than the Kedrovo beds, but also belong to the Babii Kamen’ locality. In addition to this nymph, the front wing of a Chauliodites insect not different from Ch. cancellatus (Aristov, 2003) described from the Kedrovo beds (Aristov, 2003) was found in the Tarakanikha beds. Chaulioditidae dominate with regard to the number of specimens, as they account for 68% of the collection (five species of two genera), Geinitziidae are less abundant at 27% (six species of four genera), and Skaliciidae and Liomopteridae are represented by single specimens and account for 3 and 2%, respectively (one species of one genus for each).
The composition of the Babii Kamen’ fauna is similar to that of the same-age reculid faunas of the Tunguska basin (Vyatkian Stage of the Kemerovo Region of Russia: Sadovnikov, 2016). Chaulioditidae and Geinitziidae were found in the Nirungdakan-1, Nizhnyaya Lyulyukta-1, Anakit-1, and Anakit-3 localities, with each taxon represented by a single genus with two species (Aristov, 2011). The absence of Skaliciidae and Liomopteridae can be explained by a smaller collection size (62 specimens in Babii Kamen’ and 11 specimens in the Tunguska localities). Chaulioditidae dominate in the Tunguska localities, where they account for 82% of the specimens.
Babii Kamen’ is also similar to the Late Permian (Severodvinian) Isady locality in the Vologda Region of Russia with regard to reculid composition (Aristov, 2013b). Chaulioditidae predominated at this locality, where they accounted for 52% of the of the reculida remains (four genera with seven species), whereas Geinitziidae accounted for 36% (three genera with three species) and Liomopteridae, for 12% (four genera with four species). It should be noted that the proportion of eoblattids and reculids among all insects in Babii Kamen’ was significantly higher than in Isady.
The reculid fauna of the Vyatkian localities Sokovka and Balymotikha in the Vladimir Region and the Nedubrovo Vyatkian locality in the Vologda Region was represented by Chaulioditidae only (Aristov, 2013a). The same composition is characteristic of the Lower Triassic faunas of the Olenekian Petropavlovka in the Orenburg Region of Russia (D.S. Aristov, unpublished data) and the Middle Triassic (Anisian) locations of Germany (Bashkuev et al., 2012) and France (Aristov et al., 2011). Babii Kamen’ stands out against the background of the Vyatkian–Anisian reculid faunas due to the relatively high diversity (the presence of Skaliciidae and Liomopteridae). With this exception, there are no significant differences between the reculid faunas of the Upper Permian and Lower Triassic.
Babalidae are the endemic eoblattids found in Babii Kamen’ (Aristov, 2020), whereas Megakhosaridae and Blattogryllidae are quite widespread both in the Upper Permian and the Middle Triassic (Storozhenko and Aristov, 2014). Early Triassic eoblattids are not known at present. The Babii Kamen’ Eoblattid assemblage is significantly different from the Isady one with regard to the absence of Paleozoic elements. The last finds of Permotermopsidae, Atactophlebiidae, and Cacurgidae were recorded in Isady (Aristov et al., 2013). The composition of eoblattids from Babii Kamen, with the exception of endemic Babalidae, is characteristic of both the Vyatkian Nedubrovo and Tunguska basin localities and the Anisian Vosges. Megakhosaridae (Storozhenko and Aristov, 2014) were found in Nedubrovo, Megakhosaridae and Blattogryllidae were found in Anakit-3 (Tunguska Basin: Aristov, 2011) and the Vosges, and Mesorthopteridae were additionally found in the Vosges (Aristov et al., 2011).
Thus, due to recent findings of the Permian Liomopteridae and Skaliciidae, the Reculida fauna of Babii Kamen’ appears more Permian. The Eoblattida fauna is characteristic of both the Vyatkian within the Upper Permian and the Middle Triassic; the presence of endemic Babalidae and the absence of Mesorthopteridae distinguish it from other faunas of this time. The remaining Gryllones insects of Babii Kamen–the Mesoedischiidae orthopterans, the Subioblattidae cockroaches and the Alexarasniidae embiids–are known from the Mesozoic (Rasnitsyn et al., 2013).
The composition of the Gryllones species described earlier (Aristov, 2019) and in the present work does not contradict the assignment of the Maltsevo Formation in the Kuznetsk Coal Basin to the terminal Permian (Yan et al., 2019, and others). These deposits were previously assigned to the Lower Triassic (Shcherbakov, 2008). The assemblage was considered crisis, largely due to the dominance of Chaulioditidae, previously regarded as a “disaster taxon”, that is, a taxon common for situations of abrupt biotic changes. The wide distribution of this family in the epochs close to the temporal boundary of the Permian and Triassic border was subsequently demonstrated, and Chaulioditidae were no longer considered a disaster taxon (Ponomarenko, 2016).
The Gryllones insect fauna of Babii Kamen’ contains both Paleozoic and post-Paleozoic (appearing in the Paleozoic and passing into the Mesozoic) elements with a predominance of the latter represented by seven families. There are only three Paleozoic families, including the endemic Babalidae (see above). Given that the four Gryllones families are the first finds of the Mesozoic groups, the fauna seems to be more Mesozoic in composition. Only Reculida appear a more or less Paleozoic group. However, this does not demonstrate the Triassic age of the locality, since a high proportion of post-Paleozoic families is also characteristic of the Vyatkian localities (Rasnitsyn et al., 2013).
Despite the fact that the Babii Kamen’ deposits were formed during volcanic activity in Siberia (mudstones of this location are tufogenic in nature: Shcherbakov, 2015), the eoblattid and reculid fauna does not appear a crisis fauna. The 47 eoblattid specimens from the largest Upper Permian locality of Isady represent six families, whereas the 50 reculid specimens represent three families. The 14 eoblattid specimens from Babii Kamen’ represent three families, and the 62 reculid specimens represent four families. That is, the diversity of these two localities is quite comparable. Another cockroach order, the embiids, first appears in Babii Kamen’ (Alexarasniidae Shcherbakov family, 2015). The share of other cockroaches, the Blattida proper, in the Babii Kamen’ fauna is strongly reduced relatively to those in the older and younger localities (the share of cockroaches is 3%). However, cockroaches account for approximately half of insect remains in most of the Vyatkian and Olenekian localities of European Russia. The observed diversity also decreases, as the few cockroaches found belong to the Subioblattidae family and represent the first find of this family (Rasnitsyn et al., 2013). The identification of Blattulidae cockroaches from the Babii Kamen’ in the abovementioned study was erroneous (P. Vršanský, personal communication). No Paleozoic-Mesozoic Phylloblattidae, Argentinoblattidae, and Caloblattinidae families were found in Babii Kamen’ (Rasnitsyn et al., 2013), this possibly being due a temporary sharp (by an order of magnitude) decrease in the number of cockroaches. The proportion of cockroaches is similarly low in the localities of Anakit, Nedubrovo and Zalazna (Kemerovo, Vologda, and Kirov regions of Russia, respectively), which are of approximately the same age as the Babii Kamen’.
The Olenekian Petropavlovka in the Orenburg Region of Russia is the only large Lower Triassic fauna of insects at present. The post-crisis fauna of most Gryllones insects of this locality is poor and is represented only by the undefined orthopterans, cockroaches, and Chaulioditidae reculids (Shcherbakov et al., 2019)
The outpouring of Siberian traps as such apparently did not have catastrophic consequences for Gryllones insects. On the contrary, three of the nine Gryllones families found in Babii Kamen’ (Babalidae, Alexarasniidae, and Mesoedischiidae) were identified for the first time. The Babii Kamen’ finds are the last for the two families Liomopteridae and Skaliciidae, whereas four families (Subioblattidae, Megakhosaridae, Blattogryllidae and Chaulioditidae) pass from the Paleozoic to the Mesozoic.
Thus, changes in the Gryllones insect fauna indeed occurred at the boundary of the Paleozoic and Mesozoic, but they consisted in an increase, rather than a decrease, of diversity. Paradoxically, the reason for this could be precisely volcanic activity. The Gryllones listed above, except for Mesoedischiidae, were more or less closely associated with plant litter and, accordingly, with plants. The biological activity of volcanic soils is higher than that of non-volcanic zonal soils. This creates favorable conditions for plants and, indirectly, for stratobionts.
Our work additionally confirms the rather high, not typical for a crisis, diversity of the Babii Kamen’ fauna, which does not contradict the Permian age of this location. The same applies to the Tunguska fauna of the same age as the Babii Kamen’ and the Nedubrovo locality (assigned to the Triassic in some works: Shcherbakov, 2008). The Gryllones fauna of these localities look like depleted versions of the Babii Kamen’ fauna.
The normalized number of families (the number of families from a locality divided by the natural logarithm of the number of specimens from this locality) is known to increase at the end of the Permian (Babii Kamen’ and Tunguska: Rasnitsyn et al., 2013). Our new data confirm this increase.
The “crisis” in the localities listed above is manifested primarily as the low proportion of cockroaches (as a result of ecological depression), but not as a noticeable decline in diversity. Given the proportion of new families, this is not the end of the Permian extinction, but rather the beginning of the Mesozoic diversification, which was demonstrated earlier (Rasnitsyn et al., 2013) and is confirmed by our new data.
-
Superorder Blattidea
-
Order Reculida
-
Family Chaulioditidae Handlirsch, 1906
-
Genus Chauliodites Heer, 1864
-
Chauliodites babiy Aristov, sp. nov.
-
Plate 6, fig. 1

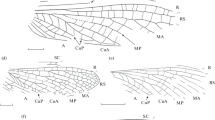
Representatives of the order Reculida from Babii Kamen’ (Kemerovo Region; Upper Permian, Vyatkian Stage), forewings: (a) Chauliodites babiy sp. nov., holotype PIN, no. 4887/212 (family Chaulioditidae); (b) Geinitzia annosa (Aristov, 2004) comb. nov., holotype PIN, no. 4887/28; (c) Stegopterum kabalum sp. nov., holotype PIN, no. 4887/520; (d) S. pashka sp. nov., holotype PIN, no. 4887/533; (e–g) Permoshurabia tshasha sp. nov.: holotype PIN, no. 4887/529; (f) paratype PIN, no. 4887/555; (g) paratype PIN, no. 4887/519; (h) Permuliercula chimaera gen. et sp. nov., holotype PIN, no. 4887/223 (family Geinitziidae). Scale bar 1 mm in Figs. 1 and 2.
Etymology. From the Babii Kamen’ locality.
Holotype. PIN, no. 4887/212, positive and negative forewing imprints; Kemerovo Region, Novokuznetsk District, right bank of the Tom’ River 10 km downstream of the Ust’-Naryk settlement, Babii Kamen’ locality; Upper Permian, Vyatkian Stage, Upper Vyatkian, Mal’tsevo formation, Kedrovo beds.

Explanation of Plate 6
Fig. 1.Chauliodites babiy sp. nov., holotype PIN, no. 4887/212, forewing (Chaulioditidae).
Fig. 2.Geinitzia annosa (Aristov, 2004) comb. nov., holotype PIN, no. 4887/28, forewing (Geinitziidae).
Fig. 3 .Stegopterum kabalum sp. nov. holotype PIN, no. 4887/520, forewing (Geinitziidae).
Fig. 4 .Stegopterum pashka sp. nov. holotype PIN, no. 4887/533, forewing (Geinitziidae).
Figs. 5 and 6 .Permoshurabia tshasha sp. nov. (Geinitziidae), forewings: (5) holotype PIN, no. 4887/529; (6) paratype PIN, no. 4887/519.
Fig. 7 .Permuliercula chimaera sp. nov. holotype PIN, no. 4887/222, forewing (Geinitziidae).
Fig. 8 .Khosara ultima sp. nov. holotype PIN, no. 4887/568, forewing (Liomopteridae).
Fig. 9 .Permotoma aba sp. nov., holotype PIN, no. 4887/193, forewing (Skaliciidae).
Fig. 10 .Tarakanikha satura sp. nov., holotype PIN, no. 4887/351, overall appearance (Gryllones incertae ordinis).
Description (Fig. 1a). Anterior margin of the forewing convex. The costal field at the RS base 3.5 times wider than the subcostal. SC with simple and Y-shaped anterior branches, ends in the distal third of the wing. R with sparse anterior branches, curved towards the anterior edge of the wing in front of the RS base and behind it. The interradial field widens sharply at the base. RS base located at the end of the basal quarter of the wing, RS starts branching anterior to the distal third of the wing and forms two or more branches. M starts branching at a certain distance from the RS base. MA starts branching close to the base and forms two branches, MP simple. CuA divided into simple CuA1 and CuA2 immediately anterior to the RS base. CuA1 and CuA2 apices not fused, CuA1 apex ends in the distal third of the wing. The intercubital field is not widened at the base. A1 simple, A2 with two branches that do not form anastomoses. The simple cross-veins form a double series of cells in the interradial field. Wing membrane dark with small light spots.
Dimensions, mm: forewing length 10.
Comparison. The M structure in the new species is most similar to that in Ch. issadensis Aristov, 2009 from the Upper Permian Isady locality in Russia, but is distinguished from it by the non-fused apices of CuA branches. The apices of CuA1 and CuA2 are fused in Ch. issadensis (Aristov, 2013b).
Material. Holotype.
-
Family Geinitziidae Handlirsch, 1906
Identification Table for the Permian Geinitziidae genera
1(2) CuA with four branches Geinitzia........................ ...................................................=Handlirsch, 1906
2(1) CuA with three branches.
3(4) CuA2 apical part curved parallel to the wing’s posterior margin.............Shurabia Martynov, 1937
4(3) CuA2 apical part not curved parallel to the wing’s posterior margin.
5(6) RS plicate backwards................................................................................Stegopterum Sharov, 1961
6(5) RS dichotomic or irregularly branched.................................................Permoshurabia Aristov, 2009
-
Genus Geinitzia Handlirsch, 1906
-
Geinitzia annosa (Aristov, 2004) comb. nov.
-
Plate 6, fig. 2

Representatives of the order Reculida and Gryllones incertae ordinis from Babii Kamen’ (Kemerovo Region; Upper Permian, Vyatkian Stage): (a) Khosara ultima sp. nov., holotype PIN, no. 4887/568 (family Liomopteridae), forewing; (b) Permotoma aba gen. et sp. nov., holotype PIN, no. 4887/193 (family Skaliciidae), forewing; (c) Tarakanikha satura gen. et sp. nov., holotype PIN, no. 4887/351 (Gryllones incertae ordinis), overall appearance.
Shurabia annosa: Aristov, 2004, p. 51, Figs. 2b and 3b.
Holotype. PIN, no. 4887/28, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 1b). Anterior margin of the forewing weakly convex. The costal field at the RS base 1.6 times wider than the subcostal. SC with dense simple anterior branches, ends by the wing’s middle. Anterior R branches simple and dense. The four-branch RS starts branching in the basal third of the wing, and the interradial field is expanded at the middle. MA with a short fork at the wing’s apex, MP starts branching in the distal third of the wing, with two endings. CuA1 starts branching in its basal third, posterior CuA1 branch with a short fork. CuA2 apical part curved parallel to the wing’s posterior margin. CuP with arc-like curvature directed towards the wing’s anterior edge. Spot- and band-like coloring along the cross-veins in the wing’s posterior half.
Dimensions, mm: forewing length approximately 11.
Comparison. A short fork at the posterior CuA1 branch distinguishes G. annosa from other species of the genus. The posterior CuA1 branch in other Geinitzia spp. is branched at the base (Storozhenko, 1998).
Remarks. The species was erroneously assigned to the genus Shurabia when first described (Aristov, 2004). A four-branch CuA typical for the genus Geinitzia is characteristic of the species under consideration, whereas the CuA of Shurabia has three branches (see identification table).
Material. Holotype.
-
Genus Stegopterum Sharov, 1961
-
Stegopterum kabalum Aristov, sp. nov.
-
Plate 6, fig. 3
Etymology. From the Shorian kabal (ash).
Holotype. PIN, no. 4887/520, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 1c). Anterior margin of the forewing convex. The costal field at the RS base 2 times wider than the subcostal. SC with dense simple anterior branches, ends at the boundary of the wing’s distal third. R with three anterior branches. RS base located in the basal third of the wing, interradial field expanded at the middle. RS with five or more branches, plicate in the backward direction, with an additional fork on the posterior branch. M with a distinct M5, divided into MA and MP approximately in the middle of the interval between the RS base and the first fork of CuA. MA and MP start branching at the middle of the wing and form three and two branches, respectively. CuA1 separated into two simple branches anterior to its middle, and the anterior branch ends anterior to the distal third of the wing, CuA2 bent weakly at the apex. Cross-veins mostly simple, but form several cells in the interradial field.
Measurements, mm: forewing length approximately 16.
Comparison. S. kabalum is most similar to S. hirtum Sharov, 1961 from the Kazanian Kaltan locality (Kemerovo Region, Russia). The new species is distinguished by early branching of MA and a short CuA. The MA is simple in S. hirtum, and the anterior CuA branch ends in the distal third of the wing (Aristov, 2004).
Material. Holotype.
-
Stegopterum pashka Aristov, sp. nov.
-
Plate 6, fig. 4
Etymology. From the Shorian pashka (other, different).
Holotype. PIN, no. 4887/533, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 1d). Anterior margin of the forewing convex. The costal field at the RS base 1.7 times wider than the subcostal. A thin convex vein parallel to the anterior margin of the wing runs at the base of the costal field. SC with dense simple anterior branches, ends anteriorly to the distal quarter of the wing. R with four or more anterior branches. RS base located at the boundary of the basal third of the wing, interradial field expanded in the middle. RS plicate in the backward direction, with four or more branches. M5 reduced to a thin cross-vein. First forks of M and CuA close to each other, M divides into MA and MP at a considerable distance anterior to the RS base. MA and MP start branching beyond the wing’s middle, MA probably has four branches, MP with two branches. CuA1 divides into two simple branches in the middle, and the anterior branch ends anterior to the distal third of the wing, CuA2 probably curved at the apex parallel to the posterior margin of the wing. Cross-veins mostly simple, H-shaped in the interradial field and Y-shaped in the intermedial field.
Measurements, mm: forewing length approximately 13.
Comparison. The new species is most similar to S. kabalum, from which it is distinguished by a longer SC, M5 reduction, and displacement of the first M fork towards the first CuA fork. The SC in S. kabalum ends anterior to the distal third of the wing, M5 is distinct, and the first forks of M and CuA are not very close to each other (see above).
Note. A vein at the base of the costal field in S. pashka is probably a “pseudo-costa” characteristic of orthoptera-like insects. This feature is extremely rare in reculids, although not unique. A similar structure of the base of the costal field was described for Parachauliodites orthopteroides Aristov, 2013 from the Severodvinian Isady locality (Chaulioditidae family: Aristov, 2013).
Material. Holotype.
-
Genus Permoshurabia Aristov, 2009
-
Permoshurabia tshasha Aristov, sp. nov.
-
Plate 6, figs. 5, 6
Etymology. From the Shorian chash (young).
Holotype. PIN, no. 4887/529, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Figs. 1e–1g). Anterior margin of the forewing convex. The costal field at the RS base approximately 2 times wider than the subcostal. SC with dense simple (less often dichotomic) anterior branches, ends at the middle of the wing or anterior to its distal third. R with three to six anterior branches. RS base located at the boundary of the basal third of the wing, interradial field not expanded. RS with five to seven main branches, divides into two veins in the middle of the wing or at the boundary of the distal third; the anterior vein is plicate in the anterior direction and the posterior one simple or dichotomic. M5 inconspicuous, M divides into MA and MP approximately in the middle of the interval between the RS base and the first CuA fork. MA with three to four branches, MP with two to three. CuA1 divides into two simple branches at its middle or more distally, the CuA2 apex with weak S-shaped curvature. Cross-veins simple. Coloring (preserved in the paratype no. 4887/519 only) as a dark band along the anterior edge of the wing and thin bands along the cross-veins.
Measurements, mm: forewing length approximately 16.
Comparison. The anterior RS branch plicate in the anterior direction distinguishes this species from other species of the genus. The RS in P. mesenensis Aristov, 2009 from Soyana (Kazanian Stage, Arkhangelsk Region, Russia) and P. komi Aristov, 2015 from Vorkuta (Kungurian Stage, Komi Republic, Russia) is dichotomic and has four branches (Aristov, 2009; Aristov, 2015b).
Material. Holotype and paratypes PIN, nos. 4887/519 and 555 from the same locality.
-
Genus Permuliercula Aristov, gen. nov.
Etymology. From the Permian and the Latin muliercula (female; feminine).
Type species. P. chimaera sp. nov.
Diagnosis. Small insects. The costal field at the RS base of the forewing somewhat wider than the subcostal. SC ends at the middle of the wing. RS dichotomic, starts at the end of the basal third of the wing, with the first fork proximal to the middle of the wing, posterior RS branch fused to MA. The interradial field expanded, crossed by a long robust cross-vein with an S-shaped bend. M5 flows into CuA posterior to the branching point of the latter, M branches at an equal distance from the first fork of CuA and the RS base. CuA1 with two branches, CuA2 apex with an S-shaped bend.
Species composition. Type species.
Comparison. The new genus Permuliercula gen. nov. is most similar to the genus Shurabia Martynov, 1937, and distinguished from it and all other Geinitziidae genera by the M5 flowing into CuA1. The M5 in all other Geinitziidae flows into CuA proximally to its branching point, similarly to what is observed in all Reculida (Storozhenko, 1998; Aristov et al., 2009; Aristov, 2015a, b).
Remarks. The fusion of M5 to CuA after its branching is one of the key synapomorphies of the superorder Orthopteroidea (Gorochov, 2004). This anastomosis is only observed in two Gryllones insect taxa that do not belong to the abovementioned superorder: the newly discovered genus and some Cnemidolestida families of the infraorder Parmapterina (Aristov, 2016).
-
Permuliercula chimaera Aristov, sp. nov.
-
Plate 6, fig. 7
Etymology. From the Latinized Greek Chimaera (chimera). The term “chimera” is used in paleontology to denote an organism that had never existed and had been erroneously reconstructed from the remains of organisms of different species.
Holotype. PIN, no. 4887/222, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 1h). Anterior margin of the forewing convex. SC with simple anterior branches with a weak S-shaped curvature. RS with four to five branches. MA with four branches, MP with two branches. The posterior CuA1 with an S-shaped bend. Intercubital field width decreases smoothly towards the wing’s anterior margin, CuP straight.
Measurements, mm: forewing length 10.
Material. Holotype.
-
Family Liomopteridae Sellards, 1909
-
Genus Khosara Martynov, 1937
-
Khosara ultima Aristov, sp. nov.
-
Plate 6, fig. 8
Etymology. From the Latin ultima (last).
Holotype. PIN, no. 4887/568, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 2a). Anterior margin of the forewing weakly convex. The costal field at the RS base 2 times wider than the subcostal. SC with simple anterior branches, ends in the distal quarter of the wing. RS starts immediately distal to the basal third of the wing. RS, MA, and MP simple anterior to the distal third of the wing. CuA1 straight, starts branching at the apex and forms a posterior crest of three or more branches. CuA2 simple, with an S-shaped bend. MA stem with kinks at cross-vein fusion sites. Coloring as a spot in the pterostigma area.
Measurements, mm: forewing length approximately 12.
Comparison. The new species is most similar to K. permiakovae Martynov, 1937 from the Urzhumian Kargala locality (Orenburg Region, Russia). A simple MA in the middle of the wing and a plicate CuA1 distinguish the new species. The MA of K. permiakovae is branched at the middle of the wing, and CuA1 has two branches (Storozhenko, 1998).
Material. Holotype.
-
Family Skaliciidae Kukalová, 1964
-
Genus Permotoma Aristov, gen. nov.
Etymology. From the Permian and the Tom’ River; feminine.
Type species. T. aba sp. nov.
Diagnosis. Small insects. The costal field at the RS base in the forewing twice wider than the subcostal. SC ends at the beginning of the wing’s distal third. RS separated in the basal quarter of the wing and starts branching beyond its middle. M divided into MA and MP beyond the RS base. MA and MP start branching at the middle of the wing. CuA1 simple, with an S‑shaped curvature, CuA2 branched at its base.
Species composition. Type species.
Comparison. The new genus is most similar to Afrogrylloblattus Aristov et Mostovsky, 2013 from the Upper Permian Ligetton locality in South Africa due to the presence of M divided into MA and MP beyond the RS base. A narrow costal field, a simple CuA1, and CuA2 that starts branching early are the distinctive features of the genus Permotoma. The costal field at the RS base is approximately as wide as the subcostal field in Afrogrylloblattus, CuA1 branched, and CuA2 starts branching at its middle (Aristov and Mostovskii, 2013).
-
Permotoma aba Aristov, sp. nov.
-
Plate 6, fig. 9
Etymology. From the Aba Group.
Holotype. PIN, no. 4887/193, positive and negative forewing imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Kedrovo beds.
Description (Fig. 2b). Anterior margin of the forewing weakly convex. SC with simple anterior branches joined by cross-veins. Interradial field wide but narrower than the subcostal. RS and MA with two or more branches, MP with two branches. CuA2 with two or more branches. Intercubital field base not expanded. Cross-veins simple, form a double series of cells in the interradial field and in the middle of the medial field. Color along the cross-veins and as small spots at the wing’s base and first M fork and a large spot in the distal half of the wing.
Measurements, mm: forewing length approximately 12.
Material. In addition to the holotype, specimen PIN, no. 4887/236 from the same locality.
-
GRYLLONES INCERTAE ORDINIS
-
Gryllones Incertae Familiae
-
Genus Tarakanikha Aristov, gen. nov.
Etymology. From the Tarakanikha beds; feminine.
Type species. T. satura sp. nov.
Diagnosis. Head large, with large eyes. Pronotum elongated, narrowing towards the anterior edge. Paranotal ring square, does not extend over the head. The lateral blades of paranotalia become narrower towards the posterior edge of the pronotum. Mesonotum square-shaped, shorter than the pronotum, metanotum shorter than the mesonotum. Anterior legs long, with broad inermous tibia. Middle and posterior tibia long, rear-facing. Anterior and posterior wing anlagen with rounded apices, parallel to the body axis. Abdominal tergites narrow, cerci short, with short segments, sharply narrowing towards the apex.
Species composition. Type species.
Comparison. The new genus is most similar to Tshekardites Aristov, 2004 from the Lower Permian Chekarda locality (Kungurian, Perm krai, Russia). Large eyes, a pronotum that narrows in the anterior direction, and thick tibia distinguish the new genus. Tshekardites have small eyes, a square pronotum, and slender tibia (Aristov, 2004; Aristov et al., 2006). The genus Tarakanikha has a narrow pronotum and short cerci with relatively long segments, in contrast to another similar genus Kaltanympha from the Kaltan locality (Kazanian Stage, Kemerovo Region, Russia). Kaltanympha has a square pronotum and long cerci with very short segments (Storozhenko, 1998).
Remarks. The general structure plan (the presence of paranotalia that do not extend over the head, parallel anterior and posterior wing anlagen, not enlarged hind legs with backward-facing tibia and segmented cerci) of the nymph described is characteristic of two Gryllones orders, Eoblattida and Reculida (the composition of these orders is presented in Aristov, 2015a). A large head and eyes, the lateral paranotalia lobes that expand towards the head, and short cerci are the characteristic features of Megakhosaridae family eoblattids. Imago of a Tshekhosara megakhosarid from the Lower Permian of Russia (Novokshonov, 1998) is most similar to the nymph described. However, we do not consider this similarity reliable enough to assign the new nymph to Eoblattida, let alone a specific eoblattid family.
Non-structured organic gut contents, presumably of plant origin, have been preserved at the background of the abdomen. The large eyes, the backward-facing cerci characteristic of stratobionts (on the contrary, forward-facing cerci are characteristic of aquatic nymphs: Aristov et al., 2006; Vršanský et al., 2019), the long legs, and the gut contents allow for the conclusion that the species under consideration was saprophagous and dwelled in more or less open areas.
-
Tarakanikha satura Aristov, sp. nov.
-
Plate 6, fig. 10
Etymology. From the Latin satura (sate).
Holotype. PIN, no. 4887/351, positive and negative nymph imprints; Babii Kamen’ locality; Upper Permian, Mal’tsevo formation, Tarakanikha beds.
Description (Fig. 2c). Pronotum 1.3 times as long as widest. Anterior and posterior edges of the paranotal ring narrow. Posterior wing anlagen considerably wider than the anterior ones. Cercae short, with five or six segments.
Measurements, mm: body length 10.
Material. Holotype.
REFERENCES
Aristov, D.S., Revision of the family Tomiidae (Insecta: Grylloblattida), Paleontol. J., 2003, vol. 37, no. 1, pp. 31–38.
Aristov, D.S., The fauna of grylloblattid insects (Grylloblattida) from the end of the late Permian to the first half of the Triassic, Paleontol. J., 2004, vol. 38, no. 5, pp. 514–522.
Aristov, D.S., The fauna of grylloblattid insects (Grylloblattida) of the Lower Permian locality of Tshekarda, Paleontol. J., 2004, vol. 38, no. suppl. 2, pp. 80–145.
Aristov, D.S., Review of the stratigraphic distribution of Permian Grylloblattida (Insecta), with descriptions of new taxa, Paleontol. J., 2009, vol. 43, no. 6, pp. 643–651.
Aristov, D.S., New and little known Grylloblattida (Insecta) from intertrappean deposits of the Tunguska Basin of Siberia, Paleontol. J., 2011, vol. 45, no. 5, pp. 537–545.
Aristov, D.S., New Grylloblattida (Insecta) from the Upper Permian and Lower Triassic of European Russia and the Upper Permian of Kazakhstan, Paleontol. J., 2013, vol. 47, no. 1, pp. 48–56.
Aristov, D.S., 1.3. Vyatkian–Olenekian localities, Paleontol. J., 2013a, vol. 47, no. 7, pp. 669–675.
Aristov, D.S., 2.6. New grylloblattids (Insecta: Grylloblattida) from the Upper Permian of the Vologda region, Paleontol. J., 2013b, vol. 47, no. 7, pp. 751–766.
Aristov, D.S., Classification of order Eoblattida (Insecta; Blattidea) with description of new taxa, Far East. Entomol., 2015a, no. 301, pp. 1–56.
Aristov, D.S., New gryllones insects (Insecta: Gryllones) from the Permian of Russia, Paleontol. J., 2015b, vol. 49, no. 12, pp. 1310–1333.
Aristov, D.S., Classification of the Parmapterina subord. n. (Insecta: Cnemidolestida), Far East. Entomol., 2016, no. 311, pp. 13–22.
Aristov, D.S., New Gryllones insects (Insecta: Gryllones) from the Babii Kamen’ locality (the Upper Permian of Russia). Part 1. Order Eoblattida Paleontol. J., 2020, vol. 54, no. 1, pp. 47–52.
Aristov, D.S., Novokshonov, V.G., and Pan’kov, N.N., Taxonomy Systematics of fossil grylloblattid nymphs of grylloblattid insects (Insecta;: Grylloblattida), Paleontol.JZh., 2006, vol. 40, no. 1, pp. 795–895.
Aristov, D.S. and Mostovski, M.B., Grylloblattids (Insecta, Grylloblattida) from the Upper Permian of South Africa, Paleontol. J., 2013, vol. 47, no. 5, p. 495–502.
Aristov, D.S., Grauvogel-Stamm, L., and Marchal-Papier, F., New grylloblattid insects (Insecta: Grylloblattida) from the Grès à Voltzia of the Vosges (Middle Triassic of France), Paleontol. J., 2011, vol. 45, no. 2, pp. 159–166.
Aristov, D.S., Bashkuev, A.S., and Vassilenko, D.V., Review of the entomofauna, Paleontol. J., 2013, vol. 47, no. 7, pp. 658–667.
Aristov, D.S., Wappler, T., and Rasnitsyn, A.P., New and little-known grylloblattids of the family Geinitziidae (Insecta: Grylloblattida) from the Triassic and Jurassic of Europe. Asia, and South Africa, Paleontol. J., 2009, vol. 43, no. 4, pp. 418–424.
Bashkuev, A., Sell, J., Aristov, D., Ponomarenko, A., Sinitshenkova, N., and Mahler, H., Insects from the Buntsandstein of Lower Franconia and Thuringia, Paläontol. Z., 2012, vol. 85, no. 3, pp. 175–185.
Gorochov, A.V., Primitive Titanoptera and the early evolution of Polyneoptera, Chteniya pamyati N.A. Kholodkovskogo (N.A. Kholodkovskii Memorial Readings), St. Petersburg: Zool. Inst. Ross. Akad. Nauk, 2004, vol. 57, no. 1, pp. 1–54.
Gorochov, A.V., Review of Triassic Orthoptera with descriptions of new and little known taxa: Part 2, Paleontol. J., 2005, vol. 39, no. 3, pp. 272–279.
Novokshonov, V.G., Tshekhosara improvida n. gen., n. sp. (Insecta; Grylloblattida: Megakhosaridae) aus dem unteren Perm des Urals, Paläontol. Z., 1998, vol. 72, pp. 65–66.
Ponomarenko, A.G., Insects during the time around the Permian—Triassic crisis Paleontol. J., 2016, vol. 50, no. 2, pp. 174–186.
Rasnitsyn, A.P., Aristov, D.S., and Rasnitsyn, D.A., Insects of the Permian and Early Triassic (Urzhumian–Olenekian ages) and the problem of the Permian–Triassic biodiversity crisis, Paleontol. J., 2013, vol. 47, no. 7, pp. 793–823.
Rasnitsyn, A.P., Aristov, D.S., and Rasnitsyn, D.A., Insects at the borderline between the Permian and the Early Triassic (Urzhum-Olenek) and the problem of Permian–Triassic biodiversity crisis, Zh. Obshch. Biol., vol. 74, no. 1, pp. 43–65.
Sadovnikov, G.N., The Trappean volcanism of Siberia and the “Permian–Triassic extinction”, Geol. Razved., 2016, no. 2, pp. 8–14.
Shcherbakov, D.E., On Permian and Triassic insect faunas in relation to biogeography and the Permian–Triassic crisis, Paleontol. J., 2008, vol. 42, no. 1, pp. 15–31.
Shcherbakov, D.E., Permian and Triassic ancestors of webspinners (Embiodea), Russ. Entomol. J., 2015, vol. 24, no. 3, pp. 187–200.
Shcherbakov, D.E., Bashkuev, A.S., Vasilenko, D.V., Karasev, E.V., Lukashevich, E.D., Tarasenkova, M.M., Strel’ni-kova, O.D., and Fel’ker, A.S., Petropavlovka, a new Lower Triassic insect locality, Paleostrat-2019. Godichn. sobranie (nauch. konf.) sektsii paleontol. MOIP i Mosk. otdeleniya Paleontol. ob-va pri RAN. Moskva, 28–30 yanvarya 2019 g. (Paleostrat-2019. Ann. Meeting (Sci. Conf.) Moscow Naturalist Soc. Paleontol. Section and Moscow Section of Paleontol. Soc., Russ. Acad. Sci., Moscow, January 28–30, 2019), Moscow: Paleontol. Inst. Ross. Akad. Nauk, 2019, pp. 68–69.
Storozhenko, S.Yu., Systematika, filogeniya i evolyutsiya grilloblattidovykh nasekomykh (Insecta: Grylloblattida) (Systematics. Phylogeny, and Evolution of Grylloblattid Insects: Insecta: Grylloblattida), Vladivostok: Dal’nauka, 1998.
Storozhenko, S.Yu. and Aristov, D.S., Review of the Paleozoic and Mesozoic families Megakhosaridae and Blattogryllidae (Insecta: Grylloblattida), Far East. Entomol., 2014, no. 271, pp. 1–28.
Vršanský, P., Sendi, H., Aristov, D., Bechly, G., Müller, P., Ellenberger, S., Azar, D., Ueda Kyoichiro, Barna, P., and Garcia, T., Ancient roaches further exemplify ‘no land return’ in aquatic insects, Gondwana Res., 2019, vol. 68, pp. 22–33. https://doi.org/10.1016/j.gr.2018.10.020
Yan, E.V., Beutel, R.G., Lawrence, J.F., Yavorskaya, M.I., Hörnschemeyer, T., Pohl, H., Vassilenko, D.V., Bashkuev, A.S., and Ponomarenko, A.G., Archaeomalthus—(Coleoptera, Archostemata) a 'ghost adult' of Micromalthidae from Upper Permian deposits of Siberia?, Histor. Biol., 2019. https://doi.org/10.1080/08912963.2018.1561672
ACKNOWLEDGMENTS
The author is grateful to A.P. Rasnitsyn and A.G. Ponomarenko (PIN) and to S.Yu. Storozhenko (FSC Biodiversity FEB RAS) for comments on the manuscript.
Funding
This work was supported by Program No. 15, “Problems of the Origin of Life and Biosphere Formation,” of the Presidium of the Russian Academy of Sciences and by the Russian Foundation for Basic Research (project no. 18-04-00322).
Author information
Authors and Affiliations
Corresponding author
Additional information
Translated by S. Semenova
Rights and permissions
About this article

Cite this article
Aristov, D.S. New Gryllones Insects (Insecta: Gryllones) from the Babii Kamen’ Locality (Upper Permian of Russia). 2. Order Reculida and Gryllones Ordinis Incertis. Paleontol. J. 54, 132–142 (2020). https://doi.org/10.1134/S0031030120020021
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S0031030120020021